
Abstract
Indoleamine 2,3-dioxygenase (IDO) catalyzes the initial and rate-limiting step of tryptophan catabolism in a specific pathway, resulting in a series of extracellular messengers collectively known as kynurenines. IDO has been recognized as an authentic regulator of immunity not only in mammalian pregnancy, but also in infection, autoimmunity, inflammation, allergy, transplantation, and neoplasia. Its suppressive effects are mostly mediated by dendritic cells (DCs) and involve tryptophan deprivation and/or production of kynurenines, which act on IDO-negative DCs as well as CD4+ and CD8+ T cells. We have found that mouse IDO contains two tyrosine residues within two distinct putative immunoreceptor tyrosine-based inhibitory motifs, VPY115CEL and LLY253EGV. We have also found that Suppressor of Cytokine Signaling 3 (SOCS3)—known to interact with phosphotyrosine-containing peptides and be selectively induced by interleukin 6 (IL-6)—binds mouse IDO, recruits the ECS (Elongin-Cullin-SOCS) E3 ligase, and targets the IDO/SOCS3 complex for proteasomal degradation. This event underlies the ability of IL-6 to convert otherwise tolerogenic, IDO-competent DCs into immunogenic cells. Thus onset of immunity in response to antigen within an early inflammatory context demands that IDO be degraded in tolerogenic DCs. These studies support the finding that IDO is regulated by proteasomal degradation in response to immunogenic and inflammatory stimuli.
Introduction
Suppressor of cytokine signaling (SOCS) proteins have emerged as critical modulators of cytokine-mediated processes. 1 A series of studies with various experimental approaches have demonstrated that SOCS3 modulates interferon (IFN)-γ-inducible genes and is involved in the prevention of IFN-γ-like responses in hepatocytes and macrophages stimulated with interleukin (IL)-6.2–4 It has been suggested that, in the absence of SOCS3, IL-6 may become immunosuppressive, activating genes typically induced by IFNs. 5 Not only does the feedback inhibitor SOCS3 attenuate IL-6 signaling, 6 but also upregulation of SOCS3 by IL-6 is responsible for inhibiting the IFN-γ–driven transcriptional expression of IDO. 7 In fact, although SOCS3 may be an important regulator of IDO in response to nitric oxide, 8 an inducer of SOCS3, 9 the underlying mechanisms could be broader than simply opposing IFN-γ signaling and the IFN-γ–like actions of IL-6. 10 SOCS proteins are, in general, critical modulators of immune responses, 11 and they possess an Src homology 2 (SH2) domain, which binds phosphotyrosine-containing peptides, and a SOCS box. The latter domain participates in the formation of an E3 ubiquitin ligase complex and targets several signaling proteins for proteasomal degradation.12–15 In murine dendritic cells (DCs), treatment with IL-6 up-regulates SOCS3 and prevents IDO activation by IFN-γ.7,16 We found an inverse correlation between SOCS3 and IDO expression.10,17,18 Not only does SOCS3 influence IL-6 transcriptional programs in DCs, but it can also contribute to posttranscriptional events that directly shape the presentation pattern of DC subsets programmed to direct tolerance. In this study we demonstrate that IL-6 exerts immunogenic effects on otherwise tolerogenic CD8+ DCs that are contingent on functional SOCS3 in vivo and impaired tryptophan catabolism in vitro.
Materials and Methods
Mice and reagents
Eight- to ten-week-old female DBA/2J (H-2d) mice were obtained from Charles River Breeding Laboratories. All in vivo studies were conducted in compliance with National (Italian Parliament DL 116/92) and Perugia University Animal Care and Use Committee guidelines. Murine rIL-6 was from Peprotech. Rabbit monoclonal anti-mouse IDO antibody (cv152) was obtained as described. 19 Anti-SOCS3 and anti-Flag M2 antibodies were purchased from Abcam and Sigma-Aldrich, respectively. Anti-ubiquitin was from Cell Signaling. The P815AB tumor peptide (amino acid sequence LPYLGWLVF) was synthesized and purified as described.20,21 Biotinylated phosphorylated and unphosphorylated peptides for pull-down experiments spanning the ITIM-contained tyrosines of mouse IDO (ITIM1: SGSGNIAVPY115CELSE; pITIM1: SGSGNIAVP-pY115CELSE; ITIM2: SGSGPEGLLY253EGVWD; pITIM2: SGSGPEGLLpY253 EGVWD) were obtained from Sigma-Genosys. The IDO inhibitor 1-MT (D, L-isomers; Sigma-Aldrich) and the proteasome inhibitor MG132 (Calbiochem) were also used.
DC purification and treatments
Splenic DCs were purified using CD11c MicroBeads (Miltenyi Biotec) in the presence of EDTA to disrupt DC-T cell complexes. 22 Cells were >99% CD11c+, >99% MHC I-A+, >98% B7-2+, <0.1% CD3+, and appeared to consist of 90%–95% CD8−, 5%–10% CD8+, and 1%–5% B220+ cells. DC populations were further fractionated according to CD8 expression in order to obtain purified CD8+ DCs by means of CD8α MicroBeads (Miltenyi Biotec). After cell fractionation, the CD8+ fraction was made up of >95% CD8+ DCs. Less than 1% CD8+ and <5% CD8− DCs expressed the B220 marker, respectively 23 DCs were exposed to 20 ng/ml rIL-6 for 24 h at 37 °C. Also used, in specific experiments, were 1-MT (4 μM) and MG132 (10 μM).
RT-PCR analysis
Expression of SOCS3 (sense, 5′-CAGCCTGCGCCT CAAGACCTT-3′; antisense, 5′-GCACCAGCTTGA GTACACAGTCG-3′) transcripts were evaluated by RT-PCR analysis using specific primers. RT-PCR products were also normalized to murine Gapdh.
siRNA synthesis and transfection
These procedures have been described previously.10,24 Briefly, the siRNA sequences specific for murine SOCS3 (sense, 5′-GGAGCAAAAGGGUCAGAGGtt-3′; antisense, 5′-CCUCUGACCCUUUUGCUCCtt-3′) were selected, synthesized and annealed by the manufacturer (Ambion). For transfection, siRNAs (5 μg) in 30 μl of transfection buffer (20 mM HEPES, 150 mM NaCl, pH 7.4) were pipetted into a sterile Eppendorf tube. In a separate polystyrene tube, 6.7 μg of 1,2 dioleoyl-3-trimethylammonium-propane (DOTAP) was mixed with 30 μl of transfection buffer and then both solutions were mixed gently by pipetting several times. After incubation at room temperature for 20 min, the mixture was added to 1 ml of complete medium containing 106 DCs and incubated for 24 h at 37 °C in the presence of rIL-6. Cells were then recovered, washed and immediately used for in vivo experiments. siRNA treatment resulted in the complete disappearance of SOCS3 transcripts at 24–48 h, as described.10,24 Control treatments consisted of cells treated with negative control siRNA (Ambion). After siRNA transfection, DC viability always exceeded 70%.
Construction and expression of mouse IDO-flag
Constructs expressing mouse IDO were generated amplifying the cDNA from purified DCs (Indo gene) with primers (Table 1) containing SpeI (sense) and NotI (antisense) restriction enzyme site sequences. The antisense primer also contained an N-terminal Flag-encoding sequence and a linker sequence coding for Gly3 to ensure flexibility of the resulting Flag-tagged protein. PCR products were digested with the appropriate restriction enzymes and cloned into a pEF-BOS plasmid. Since immunostimulatory sequences present in plasmid DNA (i.e. hypomethylated CpG motifs) may produce nonspecific effects, we transfected DCs by means of mRNAs. Briefly, plasmids obtained as above were linearized, purified using a Geneprep kit (Ambion), and used as templates for the in vitro transcription reaction using the mMES-SAGE mMACHINE T7 Ultra Kit (Ambion). Concentration and quality of in vitro-transcribed mRNAs were assessed by spectrophotometry and agarose gel electrophoresis. Control mRNA was obtained from the pTRI-Xef plasmid (supplied by manufacturer) containing the Xenopus elongation factor 1α gene, which codes for a 50.2 kDa protein. One million DCs were transfected with 2 μg mRNA, using DOTAP as described above for siRNA transfection. 7
Primer sequences.

Immunization and skin test assay
For immunization in vivo, fractionated DCs, either as such or treated and/or transfected as described above, were loaded with the P815AB peptide in vitro (5 μM, 2 h at 37 °C), before intravenous injection into DBA/2J recipient hosts. Three × 105 peptide-loaded CD8+ DCs were injected. A skin test assay was used for measuring class I-restricted, delayed-type hypersensitivity responses to synthetic peptides as previously described.7,25 The response to intrafootpad challenge with the eliciting peptide was measured at 2 weeks, and results were expressed as the increase in footpad weight of peptide-injected footpads over that of respective, vehicle-injected counterparts. 7 Data are the mean ± SD for at least six mice per group.
Kynurenine assay
IDO functional activity was measured in vitro in terms of the ability of DCs to metabolize tryptophan to kynurenine, the concentrations of which were measured by HPLC as described previously.19,26
Peptide pull-down experiments, Immunoprecipitation, and immunoblot analyses
Cells [2–5 × 105/sample for immunoblots of whole cell lysates; 3 × 106 DCs/sample for immunoprecipitation] were lysed on ice in RIPA buffer (50 mM Tris-HCl, pH 7.4, 150 mM NaCl, 1% NP-40, 0.25% Na-deoxycholate, 1 mM EDTA, 1.4 mM Na3VO4, and protease inhibitors). Lysates were either immunoprecipitated with the anti-Flag antibody or directly run on SDS-PAGE. For pull-down experiments, cell lysates were incubated sequentially with 10 μg biotinylated peptide (2 h) and streptavidin agarose beads (2 h). Immunoblots involved the use of specific antibodies in combination with the appropriate horseradish peroxidase conjugates, followed by ECL.
Statistical analysis
Student's t-test was used to analyze the results of in vitro studies in which data are mean values (± SD). In the in vivo skin test assay, statistical analysis was performed using two-tailed paired t-test by comparing the mean weight of experimental footpads with that of control, saline injected counterparts. 21 Data are mean values (±SD) of three experiments with at least six mice per group per experiment, as computed by power analysis to yield a power of at least 80% with an α level of 0.05. 27
Results and Discussion
SOCS3 is required for the effect of IL-6 on CD8+ DCs
Mouse splenic DCs can present peptide antigens in an immunogenic or tolerogenic way, with the distinction depending on either the occurrence of specialized DC subsets or the maturation or activation state of the DC. 28 Environmental factors are crucial in conditioning the outcome of DC presentation of the synthetic tumor/self nonapeptide P815AB,21,29 a poorly immunogenic antigen of mouse mastocytoma P1.HTR. 30 DC populations in the spleens of DBA/2 mice consist of CD8” (~90%) and CD8+ (~10%) fractions that mediate the respective immunogenic and tolerogenic presentation of the synthetic nonapeptide P815AB. 31 Upon transfer into recipient hosts, peptide-loaded CD8− DCs initiate immunity, and CD8+ DCs initiate anergy when Ag-specific skin test reactivity is measured at 2 wk after cell transfer.20,29 Consistent with previous results of adjuvant activity by IL-6, 7 otherwise tolerogenic CD8+ DCs were made capable of immunogenic presentation of P815AB when exposed to IL-6 before peptide loading and transfer into recipient hosts (Fig. 1A). IL-6 is a multifunctional cytokine that regulates inflammatory responses, and overproduction of IL-6 is associated with autoimmunity and chronic inflammatory diseases. 5 It has been shown that IL-6 strongly activates STAT3 and induces the expression of IFN-responsive genes in SOCS3-deficient macrophages, indicating that IL-6 might mimic the actions of IFNs.2,3 We examined whether SOCS3 expression is required for IL-6 effects on CD8+ DCs. Splenic DCs were fractionated, and specific SOCS3 gene silencing was achieved in the CD8+ fraction with siRNA technology. P815AB-pulsed CD8+ DCs were injected either untreated or treated with rIL-6, with or without concomitant siRNA-SOCS3 treatment (Fig. 1A). The results showed that SOCS3 was an absolute requirement for the occurrence of regulatory effects by rIL-6 on CD8+ DCs. Thus, in the absence of SOCS3, CD8+ DCs are refractory to immunomodulation by externally added IL-6, leaving their default tolerogenic program apparently unaffected. Also, the effect of IL-6 on SOCS3-deficient CD8+ cells was due to active suppression involving IDO because it could be reversed by the addition of the enzyme inhibitor 1-MT during cell exposure to the cytokine (Fig. 1A). On analyzing IDO expression by Western blot using rabbit monoclonal IDO-specific Ab, we found increased detection of the enzyme protein after IL-6 treatment in cells lacking SOCS3 (Fig. 1B). This was reflected by enhanced conversion of tryptophan to kynurenine (Fig. 1C). Therefore, IL-6 strongly activates tryptophan catabolism in DCs with silenced expression of SOCS3. It is also of interest that IL-6 may increase SOCS3 transcriptional expression (Fig. 1D).
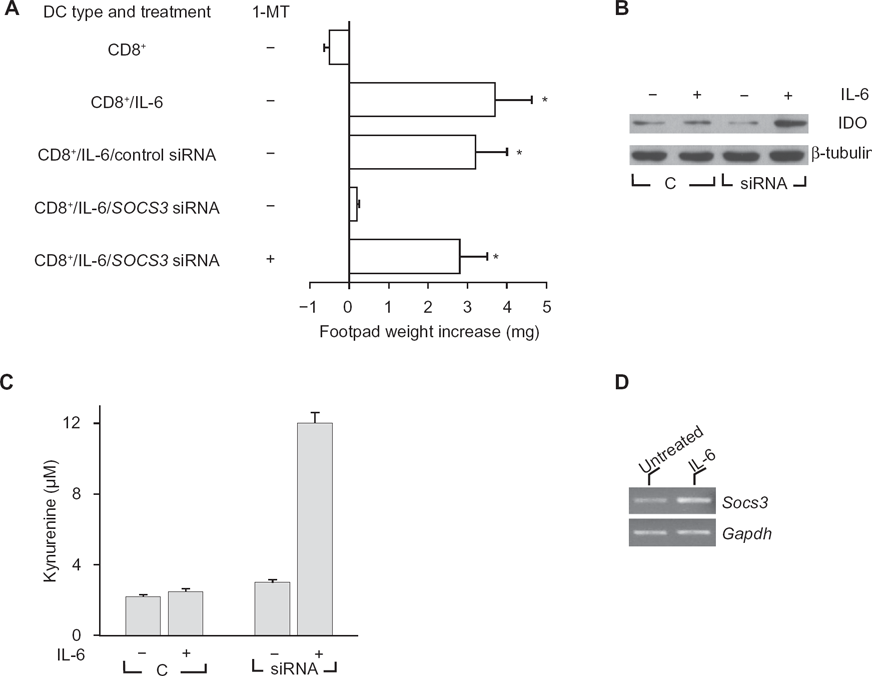
IL-6 becomes immunosuppressive in DCs lacking SOCS3, and modulation of SOCS3 by gene silencing enhances IDO expression, and tryptophan catabolism in response to IL-6.
Proteasome-mediated degradation of IDO
The mechanisms of action of SOCS proteins include SOCS box targeting of bound proteins to ubiquitinproteasome-mediated degradation. The proteasome is a major protein-degrading enzyme, which catalyzes degradation of oxidized and aged proteins, signal transduction factors and cleaves peptides for antigen presentation. As mentioned above, 1-MT is a specific and widely used inhibitor of IDO activity, 32 and MG132 is a specific proteasome inhibitor. We examined the inverse relationship between SOCS3 and IDO functions by using the two inhibitors in combination. In a skin test assay with P815AB, CD8+ DCs rendered immunogenic by IL-6 (Fig. 2A) would revert their phenotype when co-treated with MG132, yet the addition of 1-MT restored immunogenicity. Studies of IDO function in vitro with CD8+ DCs treated with IL-6 confirmed that MG132 activated the metabolic conversion of tryptophan to kynurenine—the initial IDO-dependent catabolite—and it did so in a 1-MT-sensitive manner (Fig. 2B). Therefore, an inverse relationship appeared to occur in DCs between functional IDO and SOCS3-proteasome-mediated effects.
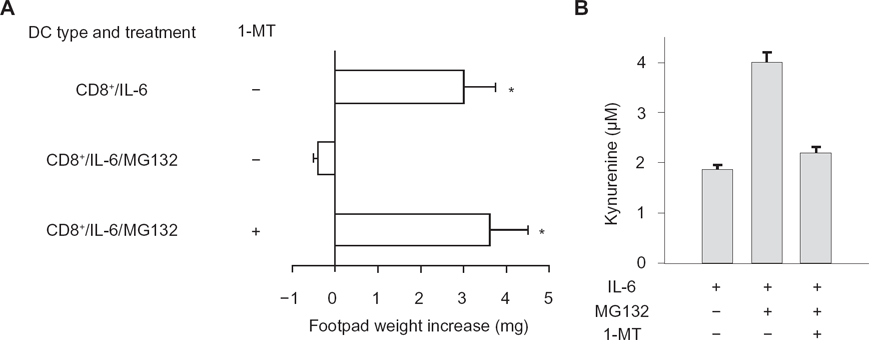
The proteasome inhibitor MG132 confers IDO-dependent, immunosuppressive properties on IL-6 in CD8+ DCs. CD8+ DCs were conditioned by overnight incubation with IL-6. The proteasome inhibitor, MG132, was added at 10 μM for 1 h before addition of the stimuli. The IDO inhibitor, 1-MT, was added to selective cultures at 4 μM.
SOCS3 binds IDO via specific phosphotyrosine binding and mediates ubiquitination and proteasomal degradation of IDO
SOCS3-associated SH2 domains bind protein sequences shared by inhibitory receptors,12,33 i.e. ITIMs. A prototypic ITIM is the I/V/L/SxYxxL/V sequence,
34
where x denotes any amino acid. As previously recognized by our studies, IDO contains two tyrosines within two distinct canonical ITIMs (ITIM1,
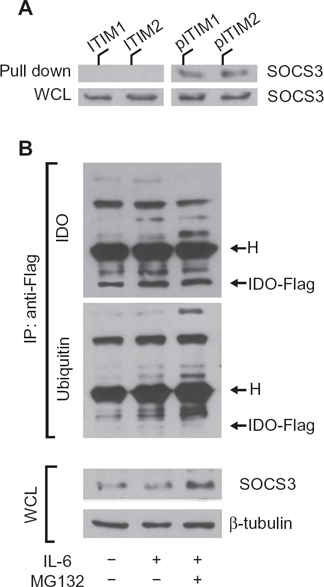
IDO contains ITIM sequences that are necessary for SOCS3-mediated ubiquitination and proteasomal degradation.
Disclosures
This manuscript has been read and approved by all authors. This paper is unique and is not under consideration by any other publication and has not been published elsewhere. The authors report no conflicts of interest.