
Abstract
Sensing the presence of a pathogen is an evolutionarily ancient trait, especially for cells of the innate immune system. The innate immune response against pathogens, such as viruses, begins with recognition of pathogen-associated molecular patterns (PAMPs) by specific pattern-recognition receptors (PRRs). Cytosolic double-stranded RNA (dsRNA) is emerging as a critical PAMP in the detection of viral infections. This recognition results in the production of antiviral and proinflammatory cytokines and, often, the death of the virus-infected cell. This review focuses on the current developments in the role of inflammasomes in response to the presence of cytosolic dsRNA in host cells. More importantly, it highlights important unanswered questions that if addressed will help us better understand the ways in which host cells respond to viral infection, in particular RNA viruses.
Keywords
Introduction
The innate immune system is the first line of defense against invading pathogens. It serves to monitor the extracellular, vacuolar, and cytosolic cellular compartments for signs of infection. To do this, the innate immune system has evolved to recognize pathogen-associated molecular patterns (PAMPs) and damage-associated molecular patterns (DAMPs) through germline-encoded pattern-recognition receptors (PRRs). 1 Several cytoplasmic and membrane-bound PRRs participate in innate immune sensing, including Toll-like receptors (TLRs), NOD-like receptors (NLRs), the cytoplasmic retinoic acid-inducible gene (RIG)-I-like receptors (RLRs), and, more recently, an expanding family of DEXDc helicases (DLRs). Microbial nucleic acids are one of the PAMPs recognized by PRRs.
Recognition of nucleic acids by specific PRRs is critical for the innate immune system to detect an infection (eg, a viral infection) and mount an appropriate response. The cytosol, in particular, represents an essential subcellular niche in the life cycle of RNA and some DNA viruses. It also functions as an escape route for a number of intracellular bacteria to evade the vacuolar killing pathways. Recognition of foreign nucleic acids may be on the basis of base modifications, sequence, unusual structures, or an abnormal location in the cell. Double-stranded RNA (dsRNA) is a replication intermediate for RNA viruses, 2 but long stretches of dsRNA are not normally found in the mammalian cell. Interestingly, dsRNA may also be formed in an infection by DNA viruses where the genome is used so economically that both strands are transcribed. TLR3 3 and TLR9 4 in the endosomal compartment are able to recognize RNA and DNA, respectively, and initiate a proinflammatory response (eg, interleukin (IL)-6 and tumor necrosis factor (TNF)-α). However, a different set of receptors is engaged 5 when nucleic acids are present in the cytosolic space, as in a viral infection.
Cytosolic double-stranded DNA (dsDNA) is recognized by the interferon (IFN)-inducible absent in melanoma 2 (AIM2) protein to activate a molecular complex known as the inflammasome,6–9 which ultimately leads to IL-1β and IL-18 secretion. Cytosolic dsDNA also induces type I IFN secretion via an unknown sensor. On the other hand, RLRs and melanoma differentiation-associated gene 5 (MDA5) recognize cytosolic dsRNA to induce a classical antiviral type I IFN response10,11 as well as apoptosis.12,13 There is evidence from the literature to suggest that intracellular dsRNA can trigger the release of proinflammatory cytokines, IL-1β and IL-18, via the inflammasome.
This review addresses the current knowledge on cellular responses elicited by cytosolic dsRNA in macrophages (Fig. 1). More importantly, the role of dsRNA in triggering inflammasome responses is discussed.
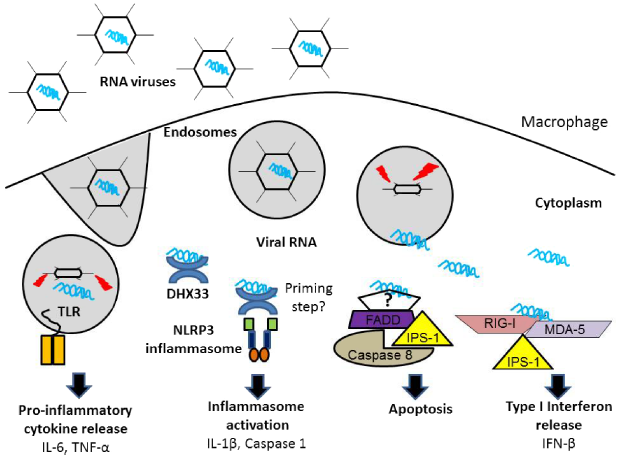
The known cellular responses to cytosolic dsRNA in primary macrophages. During an RNA virus infection, viruses are taken up by endosomes for degradation. At this point, RNA can escape into the cytosol. Viral RNA that is retained in the endosomes is recognized by TLR3 (for dsRNA) and/or TLR7/8 (ssRNA). In the cytosol, RIG-I and MDA5 recognize cytosolic 5’ triphosphorylated ssRNA to activate the classical type I IFN pathway via adaptor protein IPS-1. In addition, IPS-1 interacts with Fas-associated protein with death domain (FADD) to initiate apoptosis through caspase-8/caspase-10 activation. The RNA sensor that is triggering this apoptotic response still remains to be identified. Cytosolic dsRNA engagement with DHX33 results in NLRP3 inflammasome activation resulting in caspase 1 activation and IL-1β release. However, it is not yet clear whether an initial signal (ie, TLR agonist priming step) is required for dsRNA-mediated inflammasome activation to occur.
Cell Activation by Cytosolic Dsrna–-il-1β Release via the nlrp3 Inflammasome
Inflammasomes are cytosolic multiprotein complexes that activate caspase 1, an aspartate-specific cysteine protease family member. Activated caspase 1 then processes the inactive precursor molecules proIL-1β and proIL-18 into their biologically active secreted forms. 14 These multiprotein complexes consist of a regulatory protein (AIM2 or NLR) and, in most cases, the adaptor protein; apoptosis associated speck-like protein containing a carboxyl terminal CARD (ASC). 15 Once assembled, the inflammasome then initiates caspase 1 activation. To date, there are four well-studied inflammasomes (NLRP1, NLRP3, NLRC4, and AIM2), and each of these inflammasomes responds to a specific danger signal. 16 With the exception of AIM2, these inflammasomes contain a danger-sensing component belonging to the NLR family.
These inflammasome sensors are triggered by a range of activators. The NLRC4 inflammasome is activated by the presence of cytosolic flagellin from gram-negative bacteria or components of the type III or IV secretion systems (reviewed extensively by Franchi et al),
17
whereas NLRP1 is triggered by the anthrax lethal toxin18,19 and recently by the parasite
Microbial nucleic acids are an important group of immune stimulatory PAMPs that activate the inflammasome. AIM2 has been identified as a cytosolic dsDNA sensor that activates the ASC-containing inflammasome and triggers caspase 1-dependent IL-1β production.6–9 AIM2 has been shown to respond to dsDNA from various sources, including viruses, bacteria, and synthetic dsDNA, resulting in IL-1β secretion. Furthermore, the AIM2 inflammasome is also triggered by DNA viruses such as vaccinia virus and intracellular bacteria such as
There is increasing evidence that cytosolic dsRNA can also induce an inflammasome response. Work done with several RNA viruses has shown that they can activate the inflammasome (reviewed by Gram et al). 28 Studies implicate dsRNA recognition as the dominant signal in inflammasome activation. However, this is difficult to prove since viral proteins (eg, viral porin proteins) could also participate in triggering the inflammasome to result in physiological changes such as perturbations in intracellular Ca2+ levels.29,30 Some studies have detected activation of the NLRP3 inflammasome by synthetic dsRNA, polyinosinic:polycytidylic acid (poly I:C), and virus- and bacteria-derived dsRNA,31–36 but the current data are conflicting and others have not confirmed this.29,37 Synthetic poly I:C has been used extensively for decades to mimic the cellular effects of dsRNA, both in vivo and in vitro.
The first report implicating NLRP3 in recognition of dsRNA in primary bone marrow-derived macrophages (BMMs) included ATP in all cell treatments, and therefore, the observed triggering was probably predominately via ATP and not dsRNA. 32 One study showed that both dsRNA and single-stranded RNA (ssRNA) could elicit IL-1β secretion via NLRP3 in THP-1 cells, a monocytic cell line. 31 However, in these experiments, poly I:C was added to the cells extracellularly, whereas the ssRNA analog, ssRNA40, was complexed with a transfection reagent, Lipofectamine. Thus, the response seen with poly I:C in this study could have been a TLR3-mediated phenomenon as it is taken up endosomally. In addition, ssRNA40, a guanine–uridine (GU)-rich ssRNA, is recognized by another human TLR family member, TLR8.
It is unclear which cell type activates the inflammasome and whether initial priming is required or not prior to dsRNA transfection. In one study, poly I:C transfection in BMMs and primary bone marrow-derived dendritic cells (BMDCs), but not in THP-1 monocytic cells, induced caspase 1-mediated IL-1β secretion with LPS priming. 33 In contrast, another study showed IL-1β release in BMMs and BMDCs transfected with bacterial dsRNA without the need to prime the cells with LPS. 36 In line with the findings from Rajan et al, 33 Muruve et al 37 also showed that THP-1 cells were unable to secrete IL-1β in response to transfected poly I:C, mammalian, or viral RNA. A recent study identified that an RNA helicase family member, DHX33, can bind directly to dsRNA, and that DHX33 interacts with NLRP3 to form the inflammasome complex in both THP-1 cells and primary human monocyte-derived macrophages (HMDMs). 38 Though this was seen in HMDMs, the observation of a dsRNA-mediated inflammasome activation in THP-1 cells is questionable as it contradicts previous observations where THP-1 cells failed to secrete IL-1β in response to transfected dsRNA.33,37
The disparity in the inflammasome responses seen in primary cells (ie, BMMs and BMDCs) compared to a promonocytic cell line, such as THP-1 cells, is evident. This is not surprising as there are obvious physiological differences between these two cell types. The extent to which THP-1 cells mirror that of primary macrophages/monocytes has been questioned,39,40 despite the fact that THP-1 cells are still widely used as a model cell type to study macrophage responses. In light of these contradictory findings, the role of dsRNA in eliciting inflammasome responses warrants further investigation.
Cell Death by Cytosolic Dsrna–-Apoptosis or Pyroptosis?
Cell death is another cellular response elicited by cytosolic dsRNA. The initiation of cell death pathways is a fundamental defense against invading pathogens such as viruses,41,42 and as a consequence, viruses have evolved proteins to block these pathways. Viral RNA recognition not only results in inflammatory cytokine release (eg, IL-1β) and apoptosis but also may lead the cells to an inflammasome-mediated type of death known as pyroptosis. Pyroptosis is a caspase 1-dependent type of cell death characterized by loss of plasma membrane integrity and release of the cytoplasmic content and membrane vesicles into the extracellular milieu.43–45 Pyroptosis is also an inflammatory type of cell death where it induces an immunologically active environment, which is advantageous for a host cell, particularly a viral infection. This is in contrast to apoptosis, which is an immunologically silent cell death mediated by caspases. 46
Early work by Kibler et al 47 showed that dsRNA transfection of HeLa cells can induce apoptosis and is dependent on the activated dsRNA-dependent protein kinase (PKR). This effect was also observed when 85% of BMMs transfected with poly I:C became apoptotic eight hours post-transfection. 48 Interestingly, work by Lu et al 49 reported that PKR is required for the activation of NLRP3, NLRC4, and AIM2 inflammasomes in macrophages. The study showed that PKR was indispensable for poly I:C-mediated inflammasome activation. However, these findings were later refuted by another study that reported that PKR plays no role in activating any of the above-mentioned inflammasome sensors. 50
Iordanov et al 51 demonstrated that dsRNA triggers caspase-8 activation via the formation of a death-inducing signaling complex (DISC) in HeLa cells. Interestingly, caspase-8 activation is the apical caspase in the dsDNA-mediated apoptosis pathway involving the AIM2 inflammasome. 52 Recent work has directly implicated the RIG-I and MDA5 signaling adaptor protein, IFNβ promoter stimulator-1 (IPS-1) (also known as VISA, MAVS, and Cardif), to directly interact with caspase-8 to mediate apoptosis. 53 There-fore, it is possible that a yet unidentified RNA sensor is triggering this response via caspase-8.
In a microbial infection setting, the decision that drives a cell to an inflammasome-mediated IL-1β/IL-18 release or an inflammatory cell death fate is unknown. However, it is possible that the dose of a particular inflammasome stimulant (eg, viral RNA) could tip the balance between cell activation and cell death (ie, a high RNA dose may be cytotoxic and a low RNA dose could result in cell activation). Inflammasomes can activate both apoptotic and pyroptotic death, 52 although at high stimulus strength, pyroptosis predominates as it is rapid and lytic. Thus, the observation that dsRNA-treated cells die by apoptosis is not consistent with an inflammasome response. The key to this may be that NLRP3 inflammasome function requires priming with a TLR stimulus such as LPS. These important questions remain to be addressed.
Summary
In summary, cytosolic dsRNA is both a very potent cellular activator (ie, IFN and cytokines) and cell death stimulant. To date, the findings on the relationship between the presence of cytosolic dsRNA and activation of the inflammasome are ambiguous and inconsistent. It is likely that dsRNA can activate the inflammasome since both DNA and RNA are major viral PAMPs and that dsRNA is known to be cytotoxic. However, no work has clearly demonstrated which cell death pathway cytosolic dsRNA initiates. The decision to take the cellular activation or the cell death pathway may depend on the concentration of cytosolic dsRNA, cell type, and priming with a TLR stimulus (eg, LPS). This initial TLR priming may switch dsRNA-mediated cell death from apoptosis to inflammasome-dependent pyroptosis. To summarize, the important questions that remain to be answered are as follows. (1) Does inflammasome activation (ie, IL-1β release and caspase 1 activation) in macrophages occur with or without initial PAMP priming (eg, LPS)? (2) What is the optimal dose of dsRNA needed for macrophages to die by apoptosis? (3) Would cell death switch to pyroptosis if macrophages were pre-primed with a TLR agonist prior to dsRNA transfection? Future research in this field can contribute to the better understanding of how cells respond to an RNA virus infection. By establishing the link between cytosolic dsRNA and the inflammasome, we can then devise antiviral strategies targeting the essential components of the cellular pathways involved in eliciting dsRNA-mediated inflammatory responses.
Author Contributions
Wrote the first draft of the manuscript: AI. Contributed to the writing of the manuscript: AI. Made critical revisions and approved the final version: AI. The author reviewed and approved the final manuscript.
Footnotes
Acknowledgment
I would like to acknowledge Associate Professor Anne Cunningham from the PAP Rashidah Sa'adatul Bolkiah (PAPRSB) Institute of Health Sciences, Universiti Brunei Darussalam (UBD), for proofreading this article at the pre-submission stage.