
Abstract
Molecular biologists have long recognized carcinogenesis as an evolutionary process that involves natural selection. Cancer is driven by the somatic evolution of cell lineages. In this study, the evolution of somatic cancer cell lineages during carcinogenesis was modeled as an equilibrium point (ie, phenotype of attractor) shifting, the process of a nonlinear stochastic evolutionary biological network. This process is subject to intrinsic random fluctuations because of somatic genetic and epigenetic variations, as well as extrinsic disturbances because of carcinogens and stressors. In order to maintain the normal function (ie, phenotype) of an evolutionary biological network subjected to random intrinsic fluctuations and extrinsic disturbances, a network robustness scheme that incorporates natural selection needs to be developed. This can be accomplished by selecting certain genetic and epigenetic variations to modify the network structure to attenuate intrinsic fluctuations efficiently and to resist extrinsic disturbances in order to maintain the phenotype of the evolutionary biological network at an equilibrium point (attractor). However, during carcinogenesis, the remaining (or neutral) genetic and epigenetic variations accumulate, and the extrinsic disturbances become too large to maintain the normal phenotype at the desired equilibrium point for the nonlinear evolutionary biological network. Thus, the network is shifted to a cancer phenotype at a new equilibrium point that begins a new evolutionary process. In this study, the natural selection scheme of an evolutionary biological network of carcinogenesis was derived from a robust negative feedback scheme based on the nonlinear stochastic Nash game strategy. The evolvability and phenotypic robustness criteria of the evolutionary cancer network were also estimated by solving a Hamilton–Jacobi inequality – constrained optimization problem. The simulation revealed that the phenotypic shift of the lung cancer-associated cell network takes 54.5 years from a normal state to stage I cancer, 1.5 years from stage I to stage II cancer, and 2.5 years from stage II to stage III cancer, with a reasonable match for the statistical result of the average age of lung cancer. These results suggest that a robust negative feedback scheme, based on a stochastic evolutionary game strategy, plays a critical role in an evolutionary biological network of carcinogenesis under a natural selection scheme.
Keywords
Introduction
Biological systems are subject to intrinsic random genetic mutations, epigenetic alterations, and noises arising from internal and external stochastic fluctuations.1,2 However, it is very important that the potential perturbative effects of stochastic noise, 3 genetic mutations, and epigenetic alterations are buffered at the systemic phenotypic level. Indeed, biological systems are expected to display phenotypes that are robust to these genetic and epigenetic variations and noise.4–8 However, phenotypic changes in evolutionary fitness pressure require that biological systems are able to adapt by producing heritable phenotypic variants. This ability has been termed as evolvability. Recently, a paradoxical question about the intuitively expected trade-off between robustness and evolvability has attracted much attention9–12: how can biological systems generate phenotypes that are resistant to genetic mutations, epigenetic alterations, and noise, yet also retain their ability to evolve as a result of these effects? One answer is that the ability of a phenotype to resist mutations might enable genetic diversity, which could then be translated to phenotypic diversity as a result of subsequent genetic and epigenetic alterations, or environmental changes in selection pressure 13 through what has been termed the neutral network.8,14–16
The onset of cancer is characterized by an accumulation of genetic mutations and epigenetic alterations that are caused by different stresses, including tobacco, chemical agents, radiation, and viruses. These mutations and alterations typically modify the structures of DNA and chromatin, and consequently alter the gene products or the regulation of gene expression. 17 Molecular biologists have long recognized carcinogenesis as an evolutionary process that involves natural selection. Indeed, cancer is driven by the somatic evolution of cell lineages.18–31 In this work, the somatic evolution of cell lineages in an organ was modeled as a nonlinear stochastic biological network to study natural selection in evolutionary carcinogenesis. The evolutionary biological network of an organ is driven by intrinsic random fluctuations because of genetic and epigenetic variations and external disturbances attributed to carcinogens and other stressors. Therefore, we modeled the intrinsic random fluctuations because of genetic and epigenetic variations as a Poisson counting process, and the external disturbances were described by an uncertain signal. Therefore, an organ with different cell species undergoing carcinogenesis was modeled by a nonlinear stochastic system with an intrinsic Poisson counting process and external random disturbances.32,33
A natural selection scheme develops a robust property to efficiently attenuate intrinsic fluctuations and withstand external disturbances at the beginning of carcinogenesis. This maintains the normal function (ie, phenotype) of an evolutionary biological network under intrinsic random fluctuations because of genetic and epigenetic variations and external disturbances attributed to environmental stresses. A robust feedback scheme9–11 may exist in natural selection to maintain the healthy phenotype of a biological network at the desired equilibrium point x e (attractor; Fig. 1). Because the environmental disturbances and the intrinsic random Poisson fluctuations are unpredictable, the minimax evolution game strategy was employed. This strategy utilized a robust optimal feedback scheme that minimized the worst-case effects of external disturbances and intrinsic fluctuations on the robust stability of the phenotype at an equilibrium point favored by natural selection. Therefore, the strategy of natural selection on the evolutionary biological network during carcinogenesis was formulated as a nonlinear stochastic evolutionary game problem. Based on the stochastic Nash game strategy, effective feedback loops could be developed by natural selection for a cancer-associated biological network. This strategy can significantly improve the network's robustness to genetic and epigenetic variations and extrinsic disturbances, as well as maintain the organ's normal phenotype. However, neutral genetic mutations and epigenetic alterations gradually accumulated, and eventually overrode the network's robustness developed through natural selection. The robust stability of the biological network thereby deteriorated, which leads to a shift from the healthy phenotypic state at the equilibrium point x g to the cancerous phenotypic state at another equilibrium point x1e. Hence, cancer initiated another new evolutionary process (Fig. 1). As time increased, the abovementioned shift of phenotypic state always occurred when the accumulated genetic mutations and epigenetic alterations were large enough. Therefore, cancer can be considered an aging disease.

Four equilibrium points in the phenotypic landscape of the nonlinear stochastic biological network of an organ in (10) or (11) in carcinogenesis. The dots in the basin of the equilibrium points denote the accumulation of Poisson phenotypic fluctuations (population) of biological networks because of genetic and epigenetic changes in carcinogenesis. The vertical states illustrate the relative stability of robust states, healthy state, and tumor state of different stages at different equilibrium points, in the multiple dimensional state space. (A) At the beginning, the phenotypic trait of biological network is near the equilibrium point x e at healthy state. (B) By suffering the accumulation of neutral phenotypic variations and the influence of environmental stresses in carcinogenesis, the robust stability of healthy state cannot be maintained through the evolutionary strategy u(t) with the effect of natural selection, and then, the phenotypic trait may be shifted from x e to x1e of tumor state at the early stage to start another evolutionary process (carcinogenesis). (C) Similarly, if the accumulated neutral phenotypic variations are large enough and environmental disturbances v(t) are strong enough, the evolutionary strategy u(t) cannot confer enough robust network stability to maintain the network local stability at x1e. Then, the phenotypic trait of biological network is shifted from cancer stage I at the equilibrium point x1e to cancer stage II at the equilibrium point x2e to begin another stage of evolutionary process (carcinogenesis). (D) Finally, if the accumulation of neutral phenotypic variations is large enough to violate the phenotypic robustness criterion at x2e, cancer is transited from cancer stage II to cancer stage III. Since the equilibrium points x e in (B) and (C) are still locally stable, they are curable (treatable), ie, the curable stages in (B) and (C) may be reversible to (A) by adequate surgical or medical treatment (one kind of enhanced negative feedback). If the healthy state x e is incurable (saddle point) as shown in (D), then it is impossible that this process is revertible to (A).
In this study, based on the stochastic Nash game strategy for the robust stabilization of evolutionary nonlinear biological networks in carcinogenesis,34–39 both natural selection and external disturbances affected network evolution. Natural selection minimized network evolution (or maximized network fitness), while external disturbances, including genetic mutations and epigenetic alterations, maximized network evolution (or minimized network fitness). We solved a Hamilton–Jacobi inequality (HJI)-constrained optimization problem using the stochastic Nash game strategy to obtain a robust evolutionary feedback scheme for natural selection during carcinogenesis. Furthermore, the phenotype robustness criterion was also derived from the stochastic Nash game strategy to obtain insights into the evolvability of the cancer biological network. In order to simplify the procedure, the global linearization method was employed to interpolate several local linear stochastic systems and to approximate the nonlinear stochastic system of an organ undergoing carcinogenesis. Therefore, the HJI-constrained optimization problem could be replaced by a linear matrix inequality (LMI)-constrained optimization problem, which was solved easily using the LMI Control Toolbox in MATLAB.40–42
Finally, an in silico example of evolutionary oncogenesis in lung cancer has been provided to illustrate natural selection based on the stochastic Nash game strategy. The nonlinear stochastic biological network of lung cancer-related tissue contains parenchymal cells, stromal cells, immature vascular endothelial cells (VECs), and VECs. The network was constructed using Poisson genetic and epigenetic variations and environmental disturbances. By using a robust feedback scheme with the natural selection property based on the proposed nonlinear stochastic Nash game strategy, we found that the tissue maintained a normal phenotype for ~54.5 years in the aging process under genetic and epigenetic variations and environmental disturbances. When the robustness of the tissue network was violated by the accumulated neutral genetic and epigenetic variations and environmental disturbances, the normal phenotype of the tissue shifted from its original equilibrium point to another equilibrium point with a new, cancerous phenotype. This new phenotype24,43 took ~1.5 years to progress from stage I to stage II cancer, and about 2.5 years to progress from stage II to stage III cancer.
Stochastic Biological Network of Organ in Carcinogenesis
First, we considered a simplified analysis by using the linear biological network of a tissue or organ in carcinogenesis as the following random dynamic process.
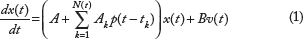



In general, the biological processes in the cancer are time-varying, ie, A(t) and B(t) in (1). In this study, because of discontinuous time-varying parameter variations, A(t) is replaced by
In order to operate successfully in the organ within the context of the organism, all the cells in the organ must be integrated into an architecture as system matrix A in the biological network of the organ in (1).
From (1), we get a linear stochastic evolutionary biological network in carcinogenesis as

Since the weighted Poisson counting process
Under the phenotypic variations and environmental disturbances in (5), in order to maintain its normal function and homeostasis, ie, to operate successfully around the equilibrium point x e , the natural selection strategy of biological network in (5) needs to select some adequate phenotypic variations to improve the biological network robustness to resist the effects of genetic and epigenetic variations and environmental changes in carcinogenesis.
Suppose that the natural selection strategy of evolutionary biological network in the organ is to select the following phenotypic variations to improve the network robustness in carcinogenesis



Therefore, the stochastic evolutionary biological network of an organ under somatic genetic and epigenetic variations, environmental disturbances, and natural selection in (5) can be represented as follows:

In the real organ, the stochastic evolutionary biological networks for n cell species, x1,…,x
n
, in carcinogenesis are always nonlinear, and the linear stochastic evolution network in (5) is thus modified as

Suppose that the phenotypic variations in (10) could be separated into the part of evolutionary strategy


The nonlinear stochastic biological system in (11) represents an evolutionary organ under evolutionary strategy
The nonlinear stochastic biological network in (10) and (11) has many equilibrium points (phenotypic traits) (Fig. 1). Suppose that the phenotypic trait of the biological network near the stable equilibrium point x
e
is the healthy state, which is the phenotypic trait with normal function of interest by natural selection. For the convenience of analysis, the origin of nonlinear stochastic biological network in (11) is shifted to the equilibrium point (phenotypic trait) x
e
. In this case, if the shifted nonlinear stochastic network is robustly stable at the origin, the equilibrium point (phenotype trait) x
e
of interest is also robustly stable. Let us denote the phenotypic deviation around the phenotype at x
e
as

In this situation, the origin
Remark 1
The linear stochastic biological network of an organ in (9) was considered as the linearized system of (13) at the equilibrium point
The nonlinear stochastic system in (13) was considered as a Darwinian dynamic equation that satisfies Darwin's conditions of variability, heritability, and the strategy to survive. 37 However, the heritability in this study implies the heritability of somatic cell evolution in carcinogenesis.
Natural Selection in Carcinogenesis
In carcinogenesis, the evolutionary biological network of an organ was modeled as the nonlinear stochastic system in (13). Under the effects of random genetic and epigenetic variations and environmental disturbance v(t), in order to maintain the normal function of evolutionary biological network to be in the healthy state at the equilibrium point x
e
in Figure 1, an evolutionary strategy u(t) =

The physical meaning of evolution performance in (14) is that the worst-case effect of the initial condition, all possible environmental disturbance v(t), neutral genetic and epigenetic variations on the robust stability performance of
The phenotypic fitness (or phenotypic robustness) of the biologic network around the healthy phenotypic trait at the equilibrium point x
e
is inversed to the evolution level of biological network,
39
ie,

In other words, a biological network with a low evolution level e in carcinogenesis will lead to a high fitness. Therefore, the maximization of the biological network fitness to the phenotypic trait at equilibrium point x
e
under the effect of genetic and epigenetic variations and environmental disturbances is equivalent to the minimization of the evolution level as

Therefore, by combining (14) and (16), the evolutionary strategy u(t) for the biological network suffers from the effect of natural selection to tolerate neutral phenotypic variations, to resist environmental disturbances, and to finally maintain the normal phenotype of biological network in carcinogenesis. It was to maximize the fitness of the biological network to the healthy state x
e
as the following minimax evolutionary game problem

The evolution game in (17) has two players u(t) and v(t). The environmental disturbance v(t) and neutral phenotype variations
In general, it is very difficult to solve the stochastic Nash game problem in (17) directly for the evolutionary strategy of nonlinear stochastic biological network of organs in carcinogenesis. In this study, a suboptimal method was introduced to solve the stochastic Nash game problem by minimizing the upper bound e of e0 indirectly as follows:

The upper bound e will be given beforehand to solve the suboptimal Nash game problem in (18) at first and e will be decreased as small as possible to approach to the solution e0 of the stochastic Nash game problem in (17).
The suboptimal stochastic Nash game problem in (18) is equivalent to the following constrained stochastic Nash game problem34,35,38
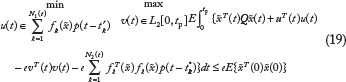
Let us denote
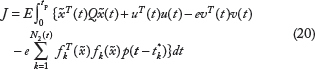
Therefore, we developed two steps to solve the constrained Nash stochastic game in (18) or (19). The first step was to solve the folla

The second step is to solve the following constraint problem

Minimax nash game strategy of linear evolutionary biological network in carcinogenesis
For the convenience of illustration, we first solve the evolutionary game strategy of linear biological network in carcinogenesis. By solving the constrained stochastic Nash game problem in (21) and (22) [or (19)] for the natural selection strategy of the linear evolutionary stochastic biological network in (9) with the effect of the natural selection in carcinogenesis, we get the following result.
Proposition 1
For the linear stochastic evolutionary biological network (9) in carcinogenesis, the constrained stochastic Nash game problem in (18) or (19) can be solved by the following evolutionary game strategy u*(t) and worst-case environmental disturbance v*(t)




Proof: see Appendix A
The Proposition 1 is the solution of the suboptimal Nash stochastic game in (18) or (19). We need to minimize the upper bound e to approach the network evolvability e0 in (17) of the linear biological network (9) in carcinogenesis. The stochastic Nash game in (17) for network evolvability e0 of the biological network in (9) needs to solve the following constrained optimization problem.

Remark 2
The solution of the above network evolvability e0 of linear biological network (9) in carcinogenesis can be obtained by decreasing e until no positive solution P in (25) within 0 < P ≤ eI in (27).
After solving e0 from (27), the Riccati-like inequality in (25) becomes

The term
After solving e0 and P > 0 from the constrained optimization problem in (27), the solution of stochastic Nash game strategy in (17) for biological network of an organ by natural selection in carcinogenesis is obtained by modifying (23) and (24) as



The eigenvalues of
Further, the phenotypic robustness criterion in (31) of evolutionary biological network in carcinogenesis was reformulated as

The physical meaning of network phenotypic robustness criterion in (32) is that if the enhanced network robustness of the biological network through negative feedback loops of the minimax evolutionary strategy in (17) can confer the environmental robustness to resist the environmental disturbances and phenotypic robustness and tolerate the neutral Poisson phenotypic variations, the phenotypic trait of biological network of an organ is maintained in carcinogenesis. Otherwise, the phenotypic trait of biological network of an organ may not be maintained under environmental disturbances and random Poisson genetic and epigenetic variations in carcinogenesis.
36
According to the statistical results,
45
the lifetime risk of many different cancers is strongly correlated (0.81) with the total number of divisions of the normal self-renewing cells maintaining that tissue's homeostasis. Also, the violation of phenotypic robustness criterion in (32) can be explained by the number of cell divisions, and a third of cancer risk among tissues is attributable to environmental factors. Namely, the cause of the violation of phenotypic robustness criterion in (32) during carcinogenesis is attributed 1/3 risk
Through the minimax game strategy of biological network of an organ in carcinogenesis, the accumulated phenotypic variations

Because the eigenvalues of (A – P) of biological network are in the far left-hand side of the complex s-domain, the biological network of an organ is more robust by the minimax game strategy of natural selection to resist environmental disturbances and to tolerate neutral phenotypic variations.
The minimax game strategy by natural selection can provide negative feedback loop –Px(t) for a greater network stability robustness improvement to buffer more neutral phenotypic variations
Minimax game strategy of nonlinear biological network in carcinogenesis
After solving the minimax game strategy of linear biological network of an organ by natural selection in carcinogenesis, we will solve the minimax game problem in (17) for the evolutionary strategy of nonlinear biological network in (13) by natural selection in carcinogenesis. First, we solved the suboptimal Nash game problem in (18), or equivalently in (21) and (22), for nonlinear stochastic biological network in (17) operating near the healthy state x e in Figure 1A.
Proposition 2
For the nonlinear stochastic biological network (13) in carcinogenesis, the suboptimal stochastic Nash game problem in (18) or (21) and (22) can be solved by the following evolutionary game strategy u*(t) and worst-case environmental disturbance v*(t)
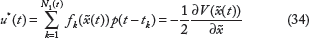



Proof: see Appendix B
After solving the above suboptimal Nash game problem in (18) or (21) and (22), for the stochastic biological network in (13), the minimax evolutionary strategy by natural selection in (17) was solved by minimizing the upper bound e as follows:

IfV(

In this situation, the evolutional game strategy u*(t) in (34) and the worst-case environmental disturbance v*(t) in (35) were modified, respectively, as follows:


The robust stability criterion in (39) was rewritten as the following phenotypic robustness criterion in carcinogenesis.

The first two terms are because of neutral phenotypic variations from the genetic and epigenetic variations in carcinogenesis, the last two terms on the left-hand side of (42) are by the worst-case environmental disturbance, and the terms on the right-hand side of (42) are because of the negative feedback through evolutionary minimax game strategy

Form (43), it is seen that
If the network robustness on the right-hand side of (42) is improved by the minimax game evolutionary strategy, from the landscape of phenotype in Figure 1A, the healthy phenotypic trait is with deeper basin and steep cliff at the equilibrium point x
e
, and the nonlinear biological network can harbor much more neutral phenotypic variations
Remark 3
For the early stage of tumor at the equilibrium point xe1 of stage I in Figure 1B, by a similar analysis as above, we could shift the origin of nonlinear stochastic biological network of an organ to xe1 as follows:

As the minimax game evolutionary strategy in (40) is chosen for nonlinear stochastic network in (43) by natural selection in the healthy stage of carcinogenesis, the biological network of an organ with early cancer stage is of the following form through the evolutionary game strategy at xe1:

As the neutral phenotypic variations
Remark 4
The feedback loop
In Figure 1B and C, the cancer states at x1e and x2e are still curable, ie, the carcinogenesis is still reversible to Figure 1A by an adequate surgical or medical treatment (one kind of enhanced negative feedback). If the healthy state is incurable (saddle point) as shown in Figure 1D, then it is impossible that this process is reversible to the healthy state in Figure 1A.
The stochastic evolutionary game of nonlinear cancer-associated network in carcinogenesis
In general, it is very difficult to solve the HJI-constrained optimization problem in (38) for the evolution game strategy u*(t) of the evolutionary biological network in (11) in carcinogenesis. At present, there is no efficient method to solve the nonlinear partial differential HJI in (39) either analytically or numerically. Therefore, in order to simplify the stochastic evolutionary game strategy, the global linearization technique 40 is employed to interpolate the nonlinear stochastic biological network in (13) with a set of local stochastic linearized biological networks like (9) at different operation points. Using the global linearization technique, 40 we suppose that all the global linearization of the nonlinear stochastic biological network in (13) are bounded by the following L vertices as 40


Based on the global linearization theory,
40
if (46) holds, every trajectory of nonlinear stochastic evolutionary biological network in (13) can be represented by a convex combination of L local linear biological networks in (47), ie,

Proposition 3
For the interpolated evolutionary biological network (48) in carcinogenesis, the stochastic evolutionary Nash game problem in (19) is solved by the following evolutionary strategy u*(t) and the worst-case environmental disturbance v*(t)
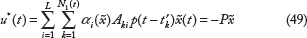



Remark 5
In comparison with Proposition 1 of a stochastic linear biological network in carcinogenesis in (9), the Riccati-like inequality in (25) is based on the local linearized network at
The Riccati-like inequalities in (51) can be considered as the local linearization of HJI in (36) at the L vertices of polytope in (46).
For solving P in (51), the L Riccati-like inequalities in (52) can be transformed to the following equivalent LMIs by the Schur complement transformation method
40

These LMIs in (53) can be easily solved by using the LMI toolbox in MATLAB. However, e in (51) and (52) is only the upper bound of the network evolvability e0. The network evolvability e0 of the nonlinear biological network under evolutionary game strategy can be obtained by solving the following LMI-constrained optimization problem

Remark 6
The e0 in (54) could be solved by decreasing e until (53) has no positive solution for 0 < P ≤ eI with the help of LMI toolbox in MATLAB.
After solving e0 in (54), the Riccati-like inequalities for the network robustness criteria of local linearized biological networks in (51) become

After solving P and e0 from (54), the evolutionary game strategy
The Riccati-like inequalities for local network robustness criteria in (51) can be rearranged as

Obviously, every interaction matrix A
i
of local linearized biological networks is shifted by
The minimax game evolutionary strategy u*(t) is to select the total amount of phenotypic variations
The phenotypic robustness criterion in (56) can be rearranged as
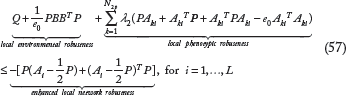
The biological meaning in (57) is that if the enhanced local network robustness of each local linearized biological network by using evolutionary game strategy u*(t) under natural selection can confer both local phenotypic robustness to buffer neutral local genetic and epigenetic variations and local environmental robustness and to resist the local environmental disturbances in carcinogenesis, the phenotype of nonlinear stochastic network with Nash game evolutionary strategy can be maintained in the basin of the equilibrium point in carcinogenesis.
From the phenotype robustness criterion in (57) in the cancer evolutionary process, it is more appealing to make the left-hand side of (57) as small as possible and to make the right-hand side of (57) as large as possible. In this situation, the coupling matrix B between biological network, environmental disturbances, and phenotypic effects A
ki
(because of genetic mutations and epigenetic alterations) should be as small as possible. In order to maintain network robustness, there always exists some membrane or transparent membrane to make B smaller to isolate the biological network from environmental disturbance, and some redundant and modular structures make the phenotypic effect A
ki
of genetic mutations and epigenetic alterations as small as possible. Further, the negative feedback loop
Because the Nash game evolutionary strategy can improve significantly the local network robustness of each local linearized biological network, the nonlinear stochastic biological network will harbor more neutral genetic and epigenetic variations. However, as the harbored neutral genetic and epigenetic variations are accumulated to the extent that the third term in (57) becomes very large and the phenotypic robustness criterion of biological network in (57) cannot be guaranteed in carcinogenesis, the phenotype of biological network might be shifted to cancer state at another equilibrium point in Figure 1 with the help of the environmental disturbances and start another period of cancer network evolution at the other equilibrium point favored by natural selection.
In Silico Example
Considering the dynamic evolutionary game strategy in the oncogenesis of lung cancer, some cell lineages related to lung cancer are described as follows. The tissue contains parenchymal cells and stromal cells. The stroma typically contains VECs, pericytes, and smooth muscle cells in its blood vessels.27,28,30,31 Along with fibroblasts and other cell types, parenchymal cells tend to exhibit considerable phenotypic variations because of genetic and epigenetic variations during oncogenesis. 24
Let x1(t) and x2(t) be the mass of parenchymal cells with phenotypes 1 and 2, respectively. Further, let y(t) be the mass of immature VECs within the tumor and z(t) be the mass of VECs, which can be represented by the length of existing microvessel in 1 g of undiseased tissue (ie, one unit is equal to microvessel length in 1 g of undiseased tissue). The dynamic system of cells related to lung cancer is thus described as follows
24
:

The variable m(t) represents tumor vascularization (perfusion) in microvessel units per gram of parenchyma [m(t), to a physiologist, is proportional to tumor microvessel length density]. The functions


In carcinogenesis, the tissue dynamic system in (58) suffers from phenotypic variations because of genetic and epigenetic variations and environmental disturbances as the following nonlinear stochastic Poisson system

The equilibrium point x of the nominal lung tissue system is at x
e
= [xe1,xe2,xe3,xe4]
T
= [6.19 × 10–11 2.20 × 10–3 0 9.88 × 10–6]
T
of the nominal system in (58) simulated from the initial condition
Based on the above evolutionary game strategy, the phenotypic variations

Based on the evolutionary game strategy in Proposition 3, natural selection is to select the phenotypic variations to construct the following negative feedback loops to improve the system robustness



The trajectories of the nonlinear stochastic biological network of lung cancer cells in carcinogenesis with states
The neutral phenotypic variations

The Poisson counting process of the neutral genetic and epigenetic variations N2(t) that are accumulated in carcinogenesis of lung cancer cell lineages in the example. The vertical axis denotes the counting number of phenotypic changes of Poisson counting process N2(t) in carcinogenesis.

The evolutionary simulation in phase plane for the biological network of in silico example in carcinogenesis. This evolutionary simulation in phase plane could support the phenotypic transition of lung cancer from the healthy state at the equilibrium point x e = [6.19 × 10–11 2.20 × 10–3 0 9.88 × 10–6] T , to the cancer stage I state at the equilibrium point x1e = [1.46 × 10–4 1.80 × 10–3 1.02 × 10–4 2.75 × 10–4] T , then to the cancer stage II state at the equilibrium point x2e = [7.09 × 10–4 1.60 × 10–3 1.30 × 10–4 3.60 × 10–4] T , and finally to the cancer stage III state at the equilibrium point x3e = [1.01 × 10–3 1.35 × 10–3 1.80 × 10–4 5.12 × 10–4] T . Moreover, the units of state x1, x2, and y are grams, and the unit of state z represents the length of existing microvessel in 1 g of undiseased tissue (ie, one unit is equal to microvessel length in 1 g of undiseased tissue).
Discussion
For ecologists and evolutionary biologists, natural selection and evolution are usually viewed as the domains of peppered moths and finches, driven to adapt by predators and competition. A long time ago, few evolutionary biologists could conceive that their field of biology would have a pivotal role in the understanding and combat of complex diseases such as cancer.
48
Recently, molecular biologists have acknowledged carcinogenesis as an evolutionary process involving natural selection that buffers neutral phenotypic variations and withstands environmental disturbances in the aging process.
49
In order to maintain the phenotypic stability of interest by natural selection, a stochastic Nash game strategy was developed using natural selection to improve the stability of a cancer-associated biological network. This strategy minimized the worst-case effects of uncertain neutral genetic and epigenetic variations and environmental disturbances on the variations from its normal carcinogenic phenotype. With the stochastic Nash game strategy, the robust phenotypic stability criterion for evolutionary biological networks in carcinogenesis is found in (42) or (57). This criterion states that the phenotypic robustness plus the environmental robustness should be less than or equal to the enhanced network robustness. Essentially, if the enhanced network robustness by the stochastic Nash game strategy can confer both phenotypic robustness to buffer neutral Poisson and epigenetic and genetic variations, and environmental robustness to withstand environmental disturbances in carcinogenesis, by considering natural selection, the network phenotype of the cancer-associated biological network can be robustly stable in the basin around the equilibrium point x
e
. In the evolutionary period, the linear network interaction matrix A is modified by
From the phenotypic robustness criterion in (42) or (57), the results reveal two ways to improve the phenotypic robustness of the evolutionary biological network in carcinogenesis. One way is to improve the stability and robustness of the network by making the right-hand side of (42) or (57) as large a value as possible so that the phenotypic robustness criterion always holds. The other way to improve the stability and robustness of the network is to reduce the effect of neutral genetic variations A
ki
and the coupling B of environmental disturbances v(t) on the biological network. This has the effect of making the left-hand side of (42) or (57) as small as possible. The evolutionary game strategy involves selecting certain somatic phenotypic variations to develop negative feedback loops
Recently, it has been reported that the phenotype of a synthetic gene network always evolves because of genetic mutations from one generation to another generation, thus making it difficult to properly design a robust synthetic gene network.43,50–53 Therefore, the robust phenotypic criterion in (42) and (57), based on the proposed stochastic Nash game strategy, may be a suitable remedy for the limitations of conventional synthetic design methods.
Conclusion
In this study, evolutionary biological networks with unpredictable somatic genetic variations and uncertain environmental disturbances in carcinogenesis were modeled as nonlinear Poisson dynamic systems. An evolutionary Nash game strategy was developed to select certain phenotypic somatic variations to construct negative feedback loops. This strategy buffered neutral genetic and epigenetic variations and unpredictable environmental disturbances to efficiently improve network robustness and to avoid shifting to the cancerous state, thus maintaining the phenotype favored by natural selection during carcinogenesis. The phenotypic robustness criterion for each cancer-related state at an equilibrium point of the evolutionary biological network during carcinogenesis was also derived from this study. We determined that the evolutionary Nash game strategy was able to select certain phenotypic variations because of somatic Poisson genetic and epigenetic variations. Negative feedback loops were constructed to make the basin of the phenotype equilibrium point significantly deeper and wider in the phenotypic landscape (Fig. 1), so that the biological network could buffer more neutral genetic and epigenetic variations and withstand additional environmental disturbances during carcinogenesis. However, as enough random neutral genetic and epigenetic variations accumulated over a long period of time, they provided the raw heterogenic material for new adaptation and evolutionary innovation. This enabled the shift to a new phenotype at a neighboring equilibrium point to commence another period of network evolution during carcinogenesis. Taken together, our results demonstrate why cancer is a disease associated with aging.
The global linearization technique was also employed to simplify the solution procedure for the nonlinear biological network in carcinogenesis. In this manner, a difficult HJI-constrained optimization problem for the evolutionary Nash game strategy could be simplified to an equivalent LMI-constrained optimization problem. Therefore, the trade-off between evolvability, genetic and epigenetic robustness, and network phenotypic robustness can be easily discussed in terms of the local stability robustness and local environmental disturbance-filtering ability of a set of local linear evolutionary biological networks. In the example of a dynamic system of cells related to lung cancer, the results of the simulation support the proposed game strategy of an evolutionary biological network during carcinogenesis.
Author Contributions
Conceived and designed the experiments: BSC. Analyzed the data: BSC. Wrote the first draft of the manuscript: BSC. Contributed to the writing of the manuscript: BSC, KWT, CWL. Agree with manuscript results and conclusions: BSC, KWT, CWL. Jointly developed the structure and arguments for the paper: BSC, KWT, CWL. Made critical revisions and approved final version: BSC, KWT, CWL. All authors reviewed and approved of the final manuscript.
Footnotes
Appendix A: Proof of Proposition 1
Appendix B: Proof of Proposition 2
Appendix C: Proof of Proposition 3
Since we replace the nonlinear stochastic network in (13) by the interpolated local linear biological network in (48), the HJI in Proposition 2 can be replaced by
The above inequality hold if the Riccati-like inequalities in (51) hold. Further,
Q.E.D.