
Abstract
Drug discovery and its methodologies have been very effective in terms of treating cancers and immunological disorders but have not been able to stop genetic diseases as most of the drugs target at the protein level. They merely mitigate the symptoms of the disease. Spinocerebellar ataxia is a neurological genetic disorder that is caused by the formation of an abnormal protein. There have been several reports on ataxic drug development but actual clinical treatment is yet to be achieved. Oligonucleotide therapy called sequence specific siRNA mediated gene silencing has evolved with promising results. This approach emphasizes on suppressing the expression of the diseased gene at mRNA level. However, there is a limitation in delivery of siRNA to the target site. Several methods have been developed over the last decade to enhance the target specific delivery of DNA, siRNA, protein and small drug molecules for therapeutic purpose with less or no side effects. This review discusses the latest upcoming technologies in the field that focus on a number of nonviral nanocarriers for targeted delivery. In this review, we explore the promise and potential of novel therapeutics with interest on ataxia therapy.
Keywords
Introduction
The DNA, a vital biomolecule of a living organism, contains a set of information coded in the form of nucleotides and is passed from generation to generation, but still no two individuals are alike because this set of information is made from the combination of two different individuals. This type of permutation and combination results in a gene pool and the dominant traits are positively selected by natural selection because of their enhanced capability of surviving, or their competency and favorable environmental conditions. Such dominant characters need not always be beneficial for an individual as they could be problematic too. Mutational events like deletion, insertion, polymorphisms and inversions produce such defective features. Such mutations if dominant in character are mostly inheritable and form the major causes of genetically inherited diseases. The cell's machinery also has some internal checks and safety mechanisms to counteract such unintentional lethalities, amongst them is the mechanism of RNA interference (RNAi), which is fast gaining prominence for its potential in therapeutic applications.
The application of RNAi as a novel therapeutic modality depends on the development of an efficient and clinically feasible means for siRNA delivery and administration. Various delivery methods have been employed to deliver such therapeutic molecules resulting in on-target efficient gene silencing. The research focuses to understand the intricacies of our biological system. The immune mechanism that guards our system is intelligent to shoot out all the invaders but the invaders are more intelligent. Their unique characteristics allow them to easily evade, cross barriers and reach their targets. Such invading systems have led researchers to mimic their natural characteristic and design a therapeutic system that may cross all the barriers and reach the affected target without meddling with the normal biological functions. Modern science recognizes these intelligent molecules as biomimetics.
In this review, we focus on genetically inherited neurodegenerative disease, Spinocerebellar ataxia and its pathogenesis that leads to degeneration of cells. The upcoming therapeutic developments using RNA interference technology, challenges faced by them and the promise shown by recent research in developing nanomedicines to combat such diseases.
Neurodegeneration–-Spinocerebellar Ataxia
Neurodegenerative diseases encompass a separate domain among neurological disorders and are specifically characterized by progressive loss of neurons as well as their structural and functional degeneration. Neurons having their primary site in brain and spinal cord perform different functions like processing information, making decisions and controlling voluntary and involuntary functions of the body. Neurodegeneration leads to loss of mobility or memory depending on which part of the brain it affects. Therefore, any defect in any part of the brain can affect a person emotionally, psychologically and physically. The defect can have many causes starting from environmental to genetical, which may lead to degeneration of the cells in brain leading to diseases like Parkinson's, Alzheimer's, Huntington's, Amyotrophic lateral sclerosis and Spino Cerebellar degeneration. These diseases tend to progress slowly over the time and generally target older population. Spinocerebellar degeneration (SCD) is a neurodegenerative disease involving loss of cells in cerebellum and spinal cord. Cells in the cerebellum are associated with regulation and coordination of movement, posture and balance. Thus, the disease leads to a state called ataxia, typified by damage to a part of the brain responsible for movement resulting in lumbering, unsteady and maladroit motion of the body due to the failure of fine coordination of muscle movements.
Different isoforms of Autosomal Dominant Cerebellar Ataxia.
Autosomal Dominant Cerebellar Ataxia
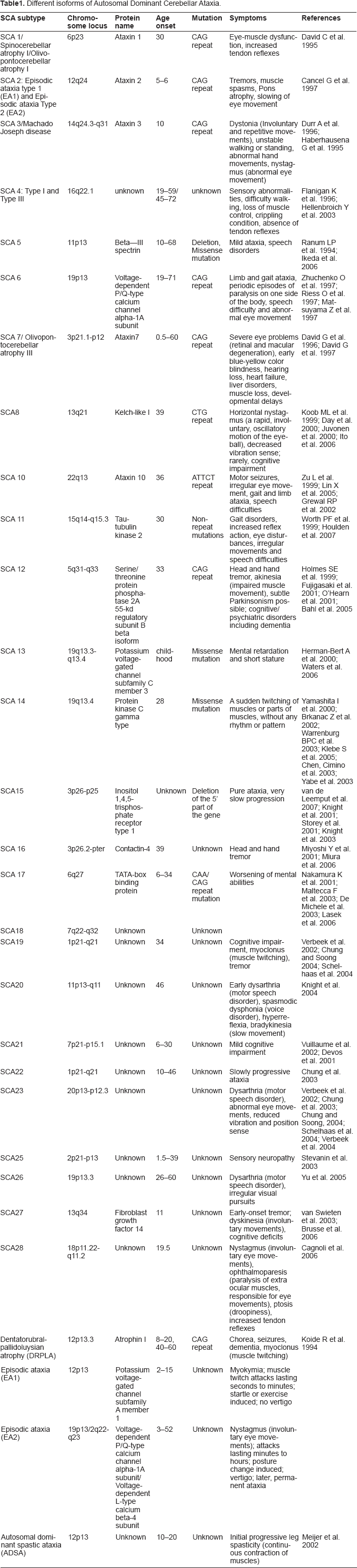
As summarized in Table 1, most of the late onset neurodegenerative diseases are caused by protein aggregation and are known as proteinopathies (Rubinsztein, 2006). These proteins are either secreted in cytoplasm or nucleus, where they alter the neuronal activities related to signaling pathways, disintegrating the neuronal network (Palop, 2006). As a consequence it causes considerable toxicity to the cells. In certain cases even genetic mutations may lead to rapid formation of fibrillar protein aggregates than wild type (Chow et al. 2004). In the case of SCD, specifically SCA1, 2, 3, 6, 7, 12, 17 and DRPLA, the disease is caused due to the formation of intranuclear inclusion bodies, which are the aggregates of an abnormal protein formed from an unstable expanded CAG trinucleotide repeat forming a poly glutamine tract (Zoghbi and Orr, 2000). The severity and the onset of disease depend on the length of CAG repeat; that is, longer the length earlier will be the onset (Duenas et al. 2006). Autosomal Dominant Cerebellar Ataxia normally sets in after the person attain the age of 25 years. It poses 50% chance of inheritance and requires the presence of mutation in only one of the two copies of a gene (maternal or paternal) to trigger the onset of the disease. Autosomal Recessive Cerebellar Ataxia and Episodic ataxia occurs before 25 years of age (Worth, 2004) and requires the inheritance of the mutation in both maternal and paternal genes. An allele having CAG triplets within the range of 14–35 with one or more CAA interruption (Sobczak et al. 2005) in between is characterized as a normal allele that results in the formation of a protein called ataxin. Whereas, an allele having more than 35 to 45 continuous CAG triplets is characterized as an abnormal allele (Ross et al. 2003), which results in the formation of an abnormal/mutant protein. Figure 1 summarizes the implication of CAG repeat sizes on the disease.
(a) The upper and lower CAG repeat sizes indicate disease and normal allele sizes respectively. (b) The abnormal allele has continuous CAG repeats whereas normal allele has CAA interruptions in between CAG repeats.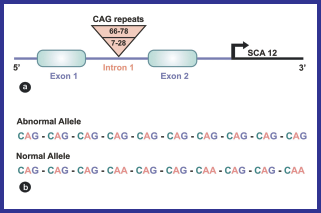
The abnormal protein formed not only accumulates within the purkinje cells (neurons) of the brain (Lin et al. 2000) but have also been observed to affect the cytoplasm, dendrites and axonal processes. It is still a controversy as to whether the formation of inclusion bodies confers a toxic effect on neurons or is actually the response of the cellular protective mechanism (Kopito, 2000). But they definitely act as biomarkers providing clues for pathogenicity (Coon et al. 2004). Also, the formation of inclusion bodies inhibits the ubiquitin-proteasome pathway (Holmberg et al. 2004; Park et al. 2005), which also relates to neurotoxicity.
Pathogenic Pathway—Formation of Aggregates and Inclusion Bodies
The pathogenic pathway involved in polyglutamine aggregation is similar to its related diseases like Alzheimer involving Aβ protein and Parkinson's involving α- synuclein that causes neurodegenerative diseases (Ross et al. 2003). Biophysical analysis of expanded polyglutamine aggregates using various methods like x-ray fiber diffraction, circular dichroism, fourier transform infra red spectroscopy revealed an antiparallel β-sheet structure of protein aggregates (Perutz, 1999; Chen et al. 2002; Poirier et al. 2002) described as “polar zipper” by Max Perutz (Perutz, 1994).
Inclusion bodies are the main sources causing the onset of disease. Their formation involves two suggestive models: Thakur and Wetzel proposed that the unfolded polyglutamine monomers undergo a structural transition after the nucleation activities resulting in an antiparallel four-stranded β-sheet helix, where each sheet comprises of seven glutamine residues. This acts as a nucleus to which similar structurally transitioned monomers forming β-sheets come together and set alternatively resulting in an elongated strand. Another pathway suggested was based on the biochemical and morphological studies of polyglutamine aggregates which involves the assembly of oligomeric intermediates resulting in the formation of protofibrils followed by fibril formation and ultimately inclusion bodies are formed (Ross et al. 2003) (Fig. 2).
Model of Polyglutamine Initiation and Elongation depicting the pathway of formation of inclusion bodies (adopted and modified from Ross et al. 2003).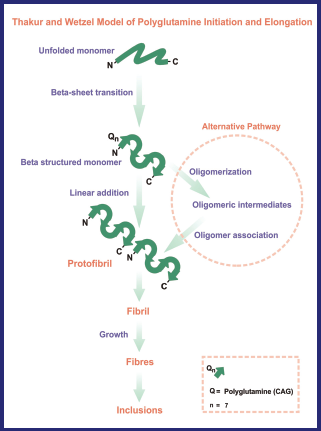
The pathogenic mechanism involving proteolytic cleavage and accumulation of protein is toxic to the nucleus (Cummings et al. 2000). The proteolytic cleavage releases toxic fragments that contain an expanded polyglutamine tail and further enhances the entry of cytoplasmic proteins into the nucleus, the mechanism of which is still not clearly understood. Other processes like apoptotic activation, accumulation, misfolding, aggregation, and sequestration of other proteins such as transcription factors and chaperones, leading to dysfunction of proteins and their intranuclear or intracellular accumulation adds up to the disease process.
Clinical Therapeutic Options
Different drugs/chemicals available for the treatment of ataxic symptoms.

Research and Therapeutics—RNAi, a New Technology for Treating Ataxia
One of the promising technology is the use of RNA interference (RNAi) to treat Ataxia. Research is being carried out in this area to develop therapeutic techniques targeting the associated SNP variant (Miller et al. 2003) by developing specific small interfering RNA (siRNA), which will silence the effect of mutant gene (Forte et al. 2005). SiRNAs, also known as “guide RNAs”, are 19 to 21bp long duplexes with 2-nucleotide 3’ overhangs (Bernstein et al. 2001; Hutvagner and Zamore, 2002) corresponding to both sense and antisense strands of a targeted gene (Hamilton and Baulcombe, 1999; Voinnet, 2002). They are formed when a DICER molecule (RNAse III type enzyme) dices the dsRNA (introduced directly or via a transgene or virus). RNAi is an effective and an efficient natural process, which prevents viral infection. It was first documented in the plant Petunia and then later in other plant species (Napoli et al. 1990). Experiments in Caenorhabditis elegans (Fire and Mello, 1998) and Drosophila (Elbashir et al. 2001) revealed that the same phenomenon also occurs in animal systems. The dsRNA when processed by DICER into siRNAs is recruited into nuclease complex to form the RNA induced silencing complex, or RISC. RISC facilitates the unwinding of siRNA, the one with the antisense strand is retained within the complex and the other (sense strand) is destroyed. RISC further guides the antisense strand to silence the targeted mRNA This therapeutic gene silencing acts before the mRNA is being translated into protein (Buckingham et al. 2004), thus it is also known as post-transcriptional gene silencing. A brief mechanism of siRNA mediated gene silencing is explained in Figure 3. Initially, immunogenicity was considered one of the major concerns with respect to siRNAs but it was later elucidated that siRNA molecules smaller than 30 bp do not cause any immune response. The major advantage of using siRNAs is their straightforward synthesis that does not involve any complex purification schemes and a cellular expression system (Sah et al. 2006). Though, siRNA may be the most effective method to treat the disease, there will be a problem with down regulating the gene due to its inherent instability, off-target effects and lack of effective delivery systems. ShRNA are also gaining importance over siRNAs as they are more stable and are expressed continuously via a plasmid or a viral vector. They are more promising showing stable targeted gene silencing (Paddison et al. 2004). They act as precursors to siRNAs (Brummelkamp et al. 2002) and comprise of a non-complimentary loop that separates sense and the antisense strands. The formation of siRNA from shRNA follows the same mechanism which includes Dicer that processes the shRNA into 19–21 long nucleotide base pairs. In order to achieve efficient gene silencing, shRNAs are placed under the control of Polymerase III promoter, which is usually a U6 small nuclear RNA promoter or RNase P H1 subunit promoter (Rodrigueze-Lebron and GonZalez-Alegre, 2006).
Mechanism of RNAi: Step 1 – Digestion of dsRNA by Dicer, Step 2 – Unwinding of formed siRNA to which a protein complex (RISC) attaches, Step 3 – Binding of RISC, directs the antisense strand of siRNA towards target mRNA, Step 4 – Degradation of the target mRNA.
Use of RNAi in in vitro studies dealing with polyglutamine diseases shows a promising therapy. Efficient, sequence specific siRNAs have been synthesized to target the mutant gene containing CAG repeat expansion or its adjacent gene sequences (Caplen et al. 2002; Miller et al. 2003). Targeting directly to CAG repeats may set an off-site suppression of normal genes or other genes containing CAG repeats that may have some important biological function. Such siRNAs have been designed and can carry out allele-specific silencing of the diseased gene without affecting its normal counterpart that is, it can target the mutant gene that differs from normal gene by only one nucleotide (Schwarz et al. 2006). Such a stringent scheme has already been developed for several neurodegenerative diseases including MJD (Machado Joseph Disease or SCA3) wherein the targeted region is the SNP (Single nucleotide Polymorphism) linked to the mutation site. This achievement in RNAi research has given a new outlook to the treatment of neurodegenerative diseases involving expansions, missense mutations like in case of SOD in amylotrophic lateral sclerosis (Ding et al. 2003), Tau in frontotemporal dementia (Miller et al. 2003), amyloid precursor protein (APP) in alzheimer's disease (Miller et al. 2004) and deletion in the case of DYT1 involving 3 base pair deletion (GAG) in coding region of TOR1A gene. The allele-specific gene silencing suppresses the expression of mutant Torsin A protein and allows continuous expression of the wild type protein (Gonzalez-Alegre, 2003).
RNAi with its promising results on primary neuronal cells (Krichevsky, 2002), mouse neuroblastoma cells (Gan et al. 2002), in vitro for treatment of neurological diseases has led a foundation for gene therapy experiments, paving its way towards in vivo experiments. Successful animal model of transgenic mice expressing human SCA1 gene has been established, the study confirms that extended CAG repeats expressed in purkinje cells produce degeneration of neurons causing ataxia (Burright et al. 1995). Animal model studies on transgenic mice were conducted by Xia et al. They delivered recombinant adeno-associated virus expressing siRNA against mutant ataxin-1 allele that code for Mutant ataxin-1 inclusion proteins in SCA1 mice model. The physiological and morphological examination of the mice revealed restoration and improvement of the cerebellum and motor coordination in mice (Xia et al. 2004).
siRNA Specificity
Different rule sets employed by different companies to develop an efficient siRNA algorithm.








Challenges in siRNA Delivery: Viral vs Non-Viral
The organizational complexity of nervous system offers many challenges to deliver therapeutic molecules. The blood-brain-barrier (BBB) and Cerebrospinal fluid (CSF) comprise a systemic vascular systemic that insulates the parenchymal and ventricular compartment of the CNS respectively. In general BBB doesn't allow the passage of any compound that is hydrophilic, ionized, and is more than 500 Da in molecular weight (Sanovich et al. 1995). Certain strategies have been developed to transiently increase the permeability of the BBB by using a bradykynin analogue, RMP-7. It stimulates the B2 receptors present on endothelial cells, increases the intracellular calcium influx and facilitates transcellular penetration by means of receptor-mediated transcytosis of therapeutic molecules (Riley et al. 1998). It is further believed that discovery and identification of novel transporters expressed on the surface of brain capillary endothelium would facilitate rapid uptake of therapeutic molecules thus, solving the problem of BBB (Forte et al. 2005).
Crossing and traversing through these insulations/barriers and reaching the targeted site that too cell specific does not only depend on the delivery mechanisms of siRNA or its carrier but also on the route of administration (Wood et al. 2003). The route of administration determines the amount of dose required to obtain an optimal effect, proper tissue distribution and minimal side-effects. For broad tissue distribution intravenous administration is often preferred while others like intraperitoneal (ip) and subcutaneous (sc) can also be utilized. Local delivery can be achieved by direct injection into joint or tumor or by topical application to the surface i.e. transepithelial application via oral, rectal or vaginal mucosa. Direct administration to central nervous system (CNS) can also be achieved by intrathecal or intraventricular injection (Behlke, 2006). Local delivery achieves less risk against systemic side effects with less amount and high concentration of the drug at the anticipated target site. Though, chemically modified duplexes are stable, they can be rapidly cleared by the plasma, therefore delivery vehicles are used to improve the durability and pharmacokinetics.
Viral Mediated siRNA Delivery
Viral delivery of therapeutic molecules has been demonstrated in few publications for neurodegenerative diseases. Recombinant adeno-associated (rAAV) virus and lentiviruses has gained a lot of importance to deliver shRNA to CNS. AAV possess a small capsid of 20 nm in diameter encasing a small genome of 4.6 Kilobases (Rodriguez-Lebron et al. 2006). Ten known AAV serotypes have already been cloned and are now being used as recombinant vectors, out of which serotype 1 and 5 have been reported to successfully transduce any part of CNS (Burger et al. 2004). Moreover, production protocols have already been optimized for large scale production of clinical grade rAAV (Grimm and Kleinschmidt, 1999). They have been used to deliver shRNAs against ataxin -1 in SCA1 transgenic mice and mediate an efficient transduction (Xia et al. 2004). The same has also been achieved against Huntington disease (Harper et al. 2005). Like Spinocerebellar ataxia, Huntington disease is also caused by CAG expansion leading to neurodegeneration but in the striatum and cortex Reduction in the production of protein after treatment can be quantified by western blot analysis and the corresponding reduction in mRNA levels can be assessed by quantitative PCR.
Likewise, lentiviral (LV) vectors have also been used to deliver shRNAs resulting in effective silencing of the diseased gene like in case of superoxide dismutase (SOD1) transgenic mouse model of ALS (Ralph et al. 2005; Raoul et al. 2005) and in alzheimer's disease by using BACE1 shRNA in LV vectors. BACE1 is responsible for initial cleavage of APP that is believed to play a major role in the pathogenesis of Alzheimer disease. Therefore, inhibition of enzyme BACE1 using shRNAs has given a new perspective to treat Alzheimer disease (Singer et al. 2005). Also, different studies have adopted use of modified virus envelopes as carriers like Fusogenic influenza virus (de Jonge et al. 2006), hemagglutinating virus of Japan (Ito et al. 2005) and SV40 pseudovirions (Kimchi-Sarfaty et al. 2005) to deliver siRNAs in vivo.
Recombinant viruses are able to transfect neurons because their envelope is replaced by another virus to achieve the tropism of the desired cell target. This is done using a technique called pseudotyping, which involves deletion of disease-promoting genes and incorporation of glycoproteins from another virus. This helps them to effectively transduce neurons (Mazarakis et al. 2001). Though the modification of retroviruses has delivered successful transfection studies as carriers, the major risk lies in their character to integrate their genetic material with the host genome, causing oncogenesis. Studies do support the activation of an immune response against viral vectors, nullifying their effect as carriers (Peden et al. 2004).
Viruses as carriers when observed to increase cellular toxicity (Davis et al. 2004), immunological complications (Lu et al. 2005) and in many cases poor intracellular uptake and limited blood stability instigated researchers to develop non-viral delivery mechanisms. siRNAs has been delivered to the central nervous system both naked or with the help of some transfection regeant in vivo targeting different molecular targets in different parts of nervous system showing effective gene silencing. Direct doses of siRNAs administered intrathecally (Dorn et al. 2004) or intracerebro-ventricularly (Thakker et al. 2004, 2005, 2006) pose a widespread inhibition of molecular targets that are broadly expressed in different parts of the brain that may lead to off- targeting. Therefore, use of transfection reagents was encouraged to use the dose in less quantity and less frequently. These transfection reagents provide simple means of gene silencing using siRNAs. Some of the widely used transfection reagents are LipofectAMINE-2000, CellFectine and Oligofectamine from Invitrogen, Effectene from Qiagen and siPORT-Amine and siPORT-Lipid from Ambion. However, cytoxicity caused by these tranfection reagents has to be taken into consideration before going into clinical trials.
Non-Viral Mediated siRNA Delivery—Nanoparticles
Considering above factors pertaining to delivery and in order to reduce cytotoxicity, poor intracellular uptake, limited blood stability and off-target effects, cationic plexes (a positively charged vehicle that can carry a negatively charged therapeutic molecule into the target cell) can be used for siRNA transfection, which will also address concerns of safety and immunogenicity of the siRNAs. These cationic polymers exist in the range of 0.1 nanometers to 100 nanometers (Schiffelers et al. 2004). They are not only small, but have entirely different physical and chemical properties than their more macro versions. They form a neutral complex when encapsulating a negatively charged molecule that can readily be taken up by the cells (neuronal) through endocytosis without showing any cytotoxic effects caused due to cationic charge. Cationic nanoplexes used for siRNA delivery are much smaller than human cells (10,000–20,000 nm in diameter) and organelles but are similar to biological macromolecules like enzymes and receptors. These nanoparticles are widely being accepted as an alternative approach to gene delivery and can be developed from cyclodextrin polycations, poly-L-Lysine, polyamidoamines, chitosan, quantum dots and liposomes to deliver siRNA for on-target gene silencing (McNeil, 2005). These particles have to overcome many anatomical, biophysical and physiological barriers that the body stages against a standard intervention such as administration of drugs or contrast agents (Molas et al. 2003) (Fig. 4). Besides bypassing these barriers, the physiology of nanoparticles is also to be considered, like the size of the carrier, which should be in accordance with type of tissue or cell to be targeted as well as its stability and safety. The final success of a drug/gene delivery system lies in the accurate delivery of nanoparticles to the desired targeted location.
Nanoparticle-mediated drug/gene delivery: overcoming the barriers.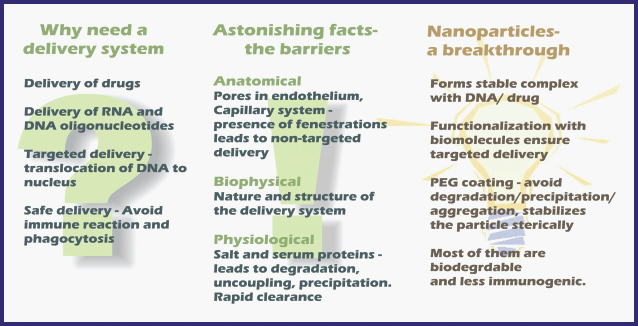
Limitations of Nanoparticles for siRNA Delivery
Biodistribution or bioavailability and toxicity of the particle along with its payload go hand in hand as the factors which govern them are interdependent on each other. The major factors are size, shape and surface charge (zeta potential) of nanoparticles (Labhasetwar, 2005). An encapsulated drug can only be released from the nanoparticle when it degrades and releases the drug at the target site. This refers to the biodistribution aspect and the removal of these degraded particles from the body with minimal toxicity effect. The purpose of using nanoparticles gets defeated if they stay inside the body, causing toxicity to the organs in the vicinity or are carried to the places where they are not required and thus cause undesired side-effects. Therefore, it is preferable to use natural materials than synthetic ones to develop nanoparticles. Natural materials pose less negative impact pertaining to toxicity and are believed to be degraded once after releasing their payload.
Size highly determines the bioavailability of the nanoparticle. The larger sized nanoparticles are more attracted by the phagocytes and are cleared from the system, though the clearance time also depends on the surface charge. It has also been observed that smaller nanoparticles can permeate faster—a nanoparticle of size 14 nm takes 2 minutes to pass through whereas a particle of size 415 nm takes 30 minutes (Szentkuti, 1997). Size of a nanoparticle varies at the time of synthesis. It also depends on different surfactants and stabilizers used over its surface to make it more biocompatible and bioavailable. While encapsulating the therapeutic molecule, the size of the particle slightly increases, likewise shape of the particle also changes. Though, shape never remains constant but is an important factor to maintain the integrity of the particle. Factors like dilution, temperature, pH, purification, surfactants/emulsifiers used and interaction with blood proteins have a major role in changing the average size and shape of the particle (Heydenreich et al. 2003; Dong and Feng, 2004; Neradovic et al. 2004). It is always preferred for a particle to be flexible enough or deformable so that it can enter and cross through the fenestrations like blood cells.
Cytotoxicity is induced mainly by the charged nanoparticles and it tends to increase with increase in the molecular weight of polymers having similar chemical structure (Fischer et al. 2003). Hence, the surface chemistry affects the cytotoxicity, inflammogenicity and fibrogenicity. Self-assembled nanoparticles (chemical monomers that naturally polymerize, creating a polymer mesh) have an advantage as altering the original monomers can modify their surface properties. Complexes whose charges are close to neutrality show high transfection efficiency by efficient receptor-mediated endocytosis but the drawback is that, neutral complexes tend to aggregate among themselves due to predominant van der Waal forces and further aggregation and adsorption of serum proteins resulting into rapid clearance of the particle. To avoid their rapid clearance by phagocyte capture they are PEGylated thus enhancing permeation and retention effect (EPR) with low toxicity and they also suppress any eliciting immunological response. PEGylation not only enhances the EPR effect but also helps to escape from reticulo-endothelial system (RES), which consists of various degrading enzymes like nuclease, proteases and lipases. Nevertheless PEGylation stabilizes the particle sterically but also stabilizes it in salt and serum solutions (Ahn et al. 2002; Oupicky et al. 2002; Park et al. 2002). When PEG is conjugated to the particle it modifies the conformation and the electrostatic binding properties thereby, reducing the surface potential of the nanoplexes with siRNAs from 35 ± 4 mV to 6 ± 1 mV (Schiffelers et al. 2004). This modification enhances the biocompatibility and stability of the particle by decreasing the electrostatic repulsion between surface charges (McNeil, 2005). The overall transfection efficiency of PEGylated nanoplexes is based on the cell types and density, chemical nature of nanoplexes, length of exposure of cells to polyplex-siRNA complexes and the size and concentration of siRNA.
The siRNAs incorporated into PEGylated nanoparticles are ligated to monoclonal antibodies or cell surface receptor ligands, which will target the molecules present on the surface of the cells. Mostly used targeting molecules are folate, luteinizing hormone-releasing hormone (LH-RH), thiamine, receptor specific peptides, aptamers and the monoclonal antibodies used against cell surface receptors such as integrins, transferrin (TFR), EGF, insulin receptors etc. (McNeil, 2005). Presence of monoclonal antibodies targeting to such surface receptors facilitates selective cell binding (Schiffelers et al. 2004) and ensures an easy entry of nanoparticles into the targeted cell by a mechanism of receptor-mediated endocytosis. These nanoparticles being biodegradable in nature release the incorporated siRNA to the target site and ensure site-specific gene silencing. The mechanism of transfection of siRNA through cationic nanocarriers in the cell via receptor-mediated endocytosis is described in Figure 5.
Mode of action of therapeutic transfection of siRNA through cationic nanocarriers in the cell via receptor-mediated endocytosis.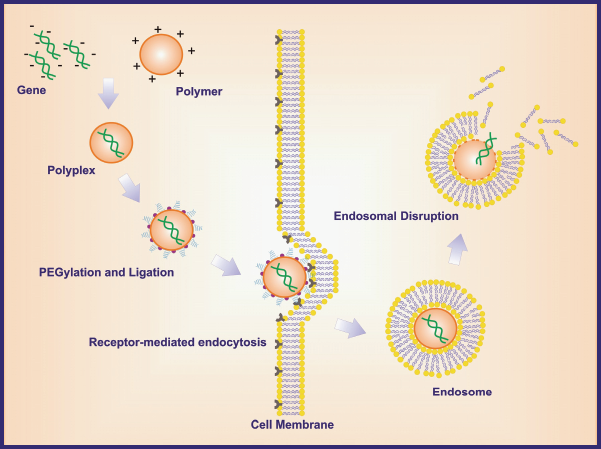
Studies reveal that the reason for reduced intracellular uptake of naked siRNAs in aqueous solution is due to their strong anionic character while in the case of PEGylated nanoplexes the steric layer reduces the cellular uptake but those which are PEGylated and have cell surface receptor ligands are easily taken up by the cell system and are successful in inhibiting the gene expression by ~90%.
Targeting CNS for therapeutic treatment has mostly been ineffective due to the tight endothelial lining made of capillaries forming BBB. As discussed before that only non-ionized, hydrophilic and low molecular weight can diffuse across the BBB. Development of synthetically designed biodegradable nanoparticles has shown a possibility to overcome this barrier. Poly (butyl cyanoacrylate) (PBCA) nanoparticles has most widely been used for brain delivery (Kreuter et al. 2001). To make PBCA nanoparticles more biocompatible and to enhance the penetration they were overcoated with polysorbate80 both in vitro and in vivo. Using this mechanism, researchers have successfully delivered several drugs like hexapeptide dalargin (Alyautdin et al. 1995), doxorubicin (anticancer drug) to the brain (Gulyaev et al. 1999). The success of using Polysorbate80 overcoated nanoparticles was because they act as an anchor for apolipoprotein E, which is involved in uptake of LDL by the brain. Upon systemic delivery of nanoparticles the uptake by reticulo-endothelial system (RES) is another barrier that restricts nanoparticles to reach BBB, overcoating with surfactants like polysorbate80 and polaxmine908 has been observed to increase the residence time of the particle and reduce uptake by RES.
Conclusion
Autosomal Dominant Cerebellar Ataxia is a complicated genetic disorder. There are specific occupational and physical therapies available for this disease. Several drugs/therapies are in the clinical trial phases. The use of siRNA has shown to be the promising approach to silence the diseased gene. However, targeted delivery of siRNA's/therapeutic molecules at the specific site matters. Precision, specificity, and efficiency is an art to achieve therapeutic benefits. However, research development and progress in medical sciences is trying to generate a new life by combining different sections of research together. Research on polymeric based biodegradable nanoparticles to be used as nanomedicne is still in its infancy and has induced researchers to explore and exploit their novel ideas.
Several issues arise when considering the delivery aspect of siRNA. A logical action is required to make siRNA drugs as attractive therapeutic option in various target sites such as liver cells, macrophages and neuronal cells that are slow dividing as an ideal place. But the problem arises in rapidly dividing cells such as cancerous cells, as the amount of siRNA gets exhausted with the very fast cell division. Also, siRNAs have very short half-life in blood and mammalian cells poorly take them. The concentration of siRNA could also pose a problem as high concentration of siRNAs can activate interferon response. The solution to this could come from the use of cationic nanoplex vectors for siRNA delivery and proper control of siRNA concentration administered to the cells.
Yet there are several questions still unanswered—what if the nanoplex delivered siRNAs compete with limited amounts of enzyme, Dicer and RISC to interfere with endogenous RNAi pathways required to maintain the cell in its differentiated state? What are its possible side effects? Will the mammalian cell take the siRNAs through endocytosis so easily? How long does the effect last?
The answers may start coming from various clinical trials, probably in cancer and infectious diseases, provided that developers can resolve technical challenges, unwanted cellular reactions, compound stability, and delivery. However, the future siRNA delivery products include nanoparticle formulations containing various targeting peptides and endo-osomolytic polymers that would play a vital role in treatment of various diseases and disorders including neurodegenerative diseases. Nanotechnologies have enabled the development of entirely new drugs and altered the properties of already marketed drugs to create potentially safer, more directed and effective pharmaceuticals. Nanotechnology enables intervention on the level at which biological systems actually operate. Pharmaceutical companies are actively pursuing this approach of using nanotechnologies that can perhaps revitalize drug pipelines to treat neurodegenerative diseases like ataxia. However, therapeutic approaches need to be standardized and optimized.
Footnotes
Acknowledgements
We acknowledge the Canadian Institute of Health Research (CIHR) grant to S. Prakash. We also acknowledge support of McGill Faculty of Medicine Postgraduate Scholarship to M. Malhotra, Ms. Jasmine Bhathena and Mr. Arun Kulamarva for proof-reading this article.