
Abstract
Myeloma is the most frequent malignancy to involve the bone. The bone microenvironment plays an important role in supporting tumor growth, bone destruction and resistance to chemotherapy. Until the advent of novel therapies such as bortezomib, the prognosis for patients with myeloma did not change significantly over 40 years. The median survival of patients until 1996 was approximately 30 months, and has now improved to almost 5 years. Bortezomib is the first-in-class proteasome antagonist approved for treatment of myeloma. It is active in newly diagnosed, relapsed and refractory patients and is now being used as a platform for combinations with other new agents for myeloma. Its major side effects include neuropathy and thrombocytopenia. In addition to its anti-myeloma effect, bortezomib also targets the bone microenvironment and can inhibit osteoclast formation, and stimulate osteoblast activity in patients with myeloma. Potentially, combination of bortezomib with other agents that stimulate bone formation or block bone resorption will further enhance the anti-myeloma effects of bortezomib and overcome the contribution of the tumor microenvironment to myeloma growth.
Multiple myeloma (MM) is a primary plasma cell malignancy, which is the most frequent malignancy to involve the bone. Over 80% of patients with MM have bone involvement during the course of their disease. 1 The prognosis for patients with MM had not changed significantly over the last 40 years until the last 5 years. 2 The median survival of patients up to 1996 was approximately 30 months, and this has now improved to almost 5 years with the advent of newer therapies. The new therapies that have made a major impact on the survival and quality of life in MM patients have been the introduction of immunomodulatory drugs (IMiDs) and bortezomib, which is the first-in-class proteasome antagonist to come to the clinic.
Both the IMiDs, thalidomide and lenalidomide, and bortezomib have been used alone and in combination with dexamethasone as well as with each other to treat MM patients. These agents have increased response rates and prolonged both progression free survival and overall survival of patients with relapsed and/or refractory MM from 12 months to 24 months and newly diagnosed MM from 30 months to 45 months. Further, these drugs also have effects on MM bone disease, either by suppressing osteoclast (OCL) activity or in the case of bortezomib enhancing osteoblast (OBL) activity.
In this review, we will examine the mechanism of action of bortezomib and its capacity to target MM cells as well as the marrow microenvironment in patients with MM.
Role of the Marrow Microenvironment in Myeloma
The marrow microenvironment plays a critical role in both tumor growth and the bone destructive process in MM (Fig. 1). MM cells home to the bone marrow by means of SDF-1 expressed by stromal cells in the marrow microenvironment and CXCR4 and CXCR7 on MM cells. 3 MM cells then bind to marrow stromal cells through adhesive interactions that activate multiple signaling pathways within the MM cells and in marrow stromal cells 4 that result in enhanced tumor growth and bone destruction. The marrow microenvironment supports the growth of MM cells and enhances osteoclastic bone destruction through expression of growth factors such as VEGF, IGF-1, IL-6, TNF-α and RANK ligands. 5 In addition, MM cells block OBL differentiation through production of the Wnt signaling antagonists, DKK1 and sFRP2, and IL-7 and IL-3. 6 All of these factors play an important role in the increased growth of tumor and bone destruction in myeloma. Thus, agents that target both MM cells and the marrow microenvironment should have a greater impact on the disease than agents which only target MM cells themselves.
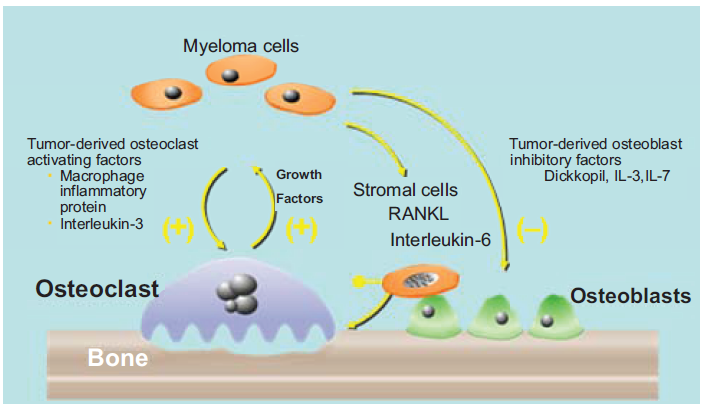
Tumor-microenvironmental interactions in myeloma. Myeloma cells produce factors that directly or indirectly activate osteoclasts such as MIP-1α and IL-3. In addition, they induce TNF-α, RANK ligand and IL-6 production by marrow stromal cells to enhance osteoclast formation and tumor growth. The bone destructive process releases growth factors such as IGF-1 and others that increase the growth of myeloma cells, further exacerbating the osteolytic process. Myeloma cells also produce DKK1, IL-3, soluble frizzle-related protein-2, and IL-7, which suppress osteoblast differentiation and new bone formation.
When MM cells home to the marrow they bind marrow stromal cells through integrins, cadherin, selectins, and syndecans expressed on the surface of the MM cells. 7 Recently, MM cells have been shown to also express the surface glycoprotein, CS-1, a member of the immunoglobulin gene superfamily, 8 as well as HLA-1 and β2-microglobulin that also play a role in tumor progression. 9 In particular, VLA-4 and VLA-5 bind to VCAM-1 and other receptors on marrow stromal cells to enhance signaling within the MM cells and marrow stromal cells that result in increased growth of tumor and osteoclastogenesis.9,10
In addition, when MM cells bind to marrow stromal cells, they become more chemoresistant. 12 These results suggest that both MM cells and the marrow microenvironment must be targeted to eradicate MM.
The Ubiquitin Proteasome System in Myeloma
The ubiquitin proteasome pathway is involved in intracellular protein degradation and is crucial to maintain cellular homeostasis.13,14 This system plays a critical role in controlling the location, activity and level of various cellular proteins. The proteasome is a multi-subunit complex, which processes ubiquitinated proteins for degradation (Fig. 2). These ubiquitinated proteins are processed through the 26S proteasome complex, which contains a 20S proteasome core and 19S flanking subunits. The 19S subunits control entry of ubiquitinated proteins into the core where they are degraded by chymotrypsin-like, trypsin-like, and caspase-like activities 13 The proteasome then degrades these ubiquitinated proteins into small peptides, which are ultimately broken down to amino-acids for new protein synthesis. To form these ubiquitinated proteins, E1 ubiquitinating enzyme activates ubiquitin and attaches it to the ubiquitin conjugating enzyme E2 in an ATP-dependent manner. E3 ubiquitin ligase then links the ubiquitin molecule to the targeted protein and repeats this process to form polyubiquitinated proteins. These polyubiquitinated proteins are then targeted to the proteasome for degradation. In addition to the 26S proteasome, there are also an immunoproteasome and the 20S core which can also play a role in protein degradation especially in conditions associated with an intensive immune response 15 Ubiquitination can affect many cellular processes in addition to proteasome degradation. Polyubiquitination tags proteins for proteasome degradation through a K48 linked polyubiquitin chain, while a K63 linked polyubiquitin chain is also formed in cells and K63-linked proteins are associated with signal transduction. 16
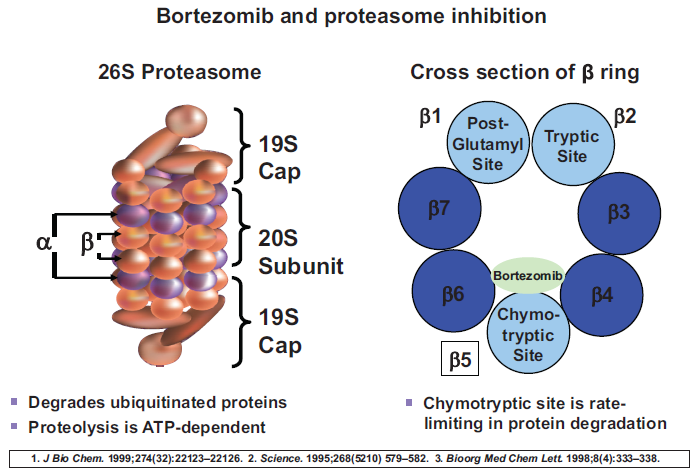
The proteasome is a multi-subunit complex, which processes ubiquitinated proteins for degradation. These ubiquitinated proteins are processed through the 26S proteasome complex, which contains a 20S proteasome core and 19S flanking subunits. The 19S subunits control entry of ubiquitinated proteins into the core where they are degraded by chymotrypsin-like, trypsin-like, and caspase-like activities. The proteasome then degrades these ubiquitinated proteins into small peptides, which are ultimately broken down to amino-acids for new protein synthesis.
Preclinical studies in a variety of tumor types suggested that proteosome inhibitors might be useful as anti-cancer agents.17,18 In addition, dose-response studies demonstrated that transformed cells were more sensitive to proteosome inhibition than normal cells. One of the initial mechanisms postulated for the effects of proteasome inhibitors on cell survival was that they blocked the NF-κB pathway. This mechanism was proposed because Iκβ binds to NF-κB and prevents the nuclear translocation of NF-κB and thereby blocks NF-κB driven transcription. When IκB is phosphorylated, it degraded by the proteasome and translocation of NF-κB to the nucleus occurs. Inhibiting proteasome activity blocks IκB degradation and stabilizes the Iκβ:NF-κB complex, preventing the transcriptional activity of NF-κB. 19 Since NF-κB plays an important role in multiple cellular processes, including cell proliferation, cell survival, cell differentiation and angiogenesis, blocking NF-κB activity should have profound effects on cell survival and tumor growth. Proteasome inhibitors also have other cellular effects including blocking degradation of p21, cyclin-dependent kinase inhibitor. Persistence of p21 inhibits cell cycle progression. 20 Protein inhibition also reduces levels of anti-apoptotic proteins such as Bcl-2 and activates caspase-9 and caspase-8-mediated apoptosis. 21
Bortezomib, the First-in-Class Proteasome Antagonist in the Clinic
As noted above, preclinical studies demonstrated the anti-tumor activity of bortezomib both on MM cell lines as well as in vivo models of MM. Use of bortezomib in human MM xenograph models showed that doses given twice weekly intravenously significantly reduced tumor volume. 22 These studies further reported that bortezomib was a potent and reversible proteasome inhibitor, which impacted multiple signaling pathways, and decreased MM cell proliferation and survival both in vitro and in vivo. These exciting preclinical findings led to the development of the CREST and SUMMIT trials, which were open-label, phase II studies of bortezomib. The CREST trial examined the efficacy of 1.0 vs. 1.3 mg/m2 of bortezomib in patients with relapsing MM. 23 The recently updated survival results of this trial showed that one and five year survival rates were 82% and 32% for the 1.0 mg/m2 dose and 81% and 45% for the 1.3 mg/m2 dose respectively. 24 These results clearly demonstrated the clinical efficacy of bortezomib in MM. In the SUMMIT trial, more than 200 patients with MM were entered. These patients were both relapsed and refractory to their most recent therapy and had received at least two prior therapies for MM. The patients received bortezomib at 1.3 mg/m2 intravenously, on days 1, 4, 8 and 11 with a 10 day rest period on a 21 day cycle. 25 The patients received 8 cycles of therapy. In this heavily pretreated group of patients receiving bortezomib alone, 28% of these patients had a complete or partial response, of which 2.7% of the patients had a complete response. These response rates were independent of both the number and types of previous therapies. Importantly, responses seen in patients whose MM cells had chromosome 13 abnormalities, which previously was a poor prognostic feature for MM treated with standard chemotherapy, and the responses were independent of performance status, type of MM and β2-microglobulin levels. 25 The median duration of response for these patients was 12.7 months, the median time to response was a very rapid 38 days, and the median overall survival of this group of patients was 17 months. 26 Approximately 1/3 of the patients developed peripheral neuropathy, 2/3 had nausea and about 1/2 the patients diarrhea and fatigue. Another common side-effect of bortezomib treatment was thrombocytopenia, which was reversible. The overwhelming majority of the side-effects were grade 1 and 2, with very few if any grade 4 toxicities. This trial demonstrated clearly that in a heavily pretreated group of patients with MM, who were both relapsed and refractory to their last therapy, that bortezomib was active and had a significant therapeutic impact on MM.
These exciting results led to an international phase III trial, the APEX study, in which patients with relapsed MM were randomized to receive either dexamethasone 40 mg/day, days 1-4, 9-12 and 17-20 every five weeks for four cycles or single agent bortezomib. 27 Patients who had progressive disease on the dexamethasone arm were able to crossover to bortezomib in a companion study. After initial induction, patients received maintenance therapy with dexamethasone or bortezomib. The updated results of this study showed that the median time to progression of the patients on bortezomib was 6.2 months versus 3.5 months (p < 0.001) and median survival was 29.8 months versus 23.7 months in the bortezomib versus dexamethasone treated group. 28 More importantly, the overall response rate was 43% in the bortezomib arm of the study compared to 18% with dexamethasone. The duration of response to bortezomib was 8 months versus 5.6 months with dexamethasone. The major adverse events associated with bortezomib therapy again were peripheral neuropathy and thrombocytopenia. As shown in the SUMMIT trial, the APEX trial confirmed that chromosome 13 deletion had no impact on either response rates or survival. In contrast, chromosome 13 deletion was associated with significant decrease of survival in the dexamethasone arm.
Based on the efficacy of bortezomib in relapsed patients, bortezomib has been used in newly diagnosed MM patients. A phase II study of bortezomib +/- dexamethasone as a first line therapy for MM was conducted by Jagannath et al. 29 They reported that bortezomib given at 1.3 mg/m2 on days 1, 4, 8 and 11 with dexamethasone given the day of and the day after bortezomib for 6 three week cycles resulted in an estimated one year survival of 87% with a median time to response of 2 months. Harousseau and coworkers have reported the use of bortezomib and dexamethasone as induction treatment before autologous stem cell transplant in newly diagnosed patients with MM. 30 In their initial trial patients were treated with bortezomib and dexamethasone prior to autologous stem cell transplant. The efficacy was very high with a 68% overall response rate, 31% of patients having either a complete remission or very good partial remission. The therapy was extremely well-tolerated with no grade 4 neuropathy or infection and no grade 3 or 4 thrombocytopenia. More recently, Orlowski and coworkers compared the combination of bortezomib with doxorubicin versus bortezomib in patients with relapsed and/or refractory MM. 31 They showed that there was a superior response duration in patients receiving bortezomib with doxorubicin compared to bortezomib (10.2 months versus 7.0 months), and that the time to progression for these patients was significantly increased in the doxorubicin-bortezomib arm versus the bortezomib arm (9.3 months versus 6.5 months, p = 0.000004).
Bortezomib and dexamethasone has also been compared to vincristine, adriamycin and dexamethasone (VAD) for induction therapy prior to autologous stem cell transplant by Harousseau and coworkers. 32 They found that the response rate after induction was significantly greater in patients receiving velcade-dexamethasone versus VAD (21% versus 8%) with an overall response rate after the first transplant of 41% versus 29% for CR and near CR. The overall response rate was 94% after the first stem cell transplant in patients receiving bortezomib and dexamethasone compared with 89% in those receiving VAD.
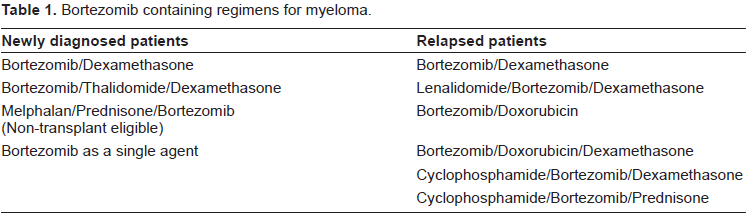
The successful use of bortezomib for newly diagnosed patients in both the transplant and non-transplant setting has led to development of multiple regimens that use combinations of bortezomib with thalidomide or lenalidomide and dexamethasone or with melphalan and prednisone (Table 1). In the VISTA trial, which was recently reported by Dr. San Miguel,33–35 the combination of bortezomib, melphalan and prednisone was compared to melphalan and prednisone (MP). This trial entered patients greater than 65 years of age or less than 65 years who were not eligible for stem cell transplant. The overall response rates in more than 300 patients receiving VMP was 82% compared with 50% for MP, with a complete remission rate of 35% and 5% in patients receiving VMP versus MP. The major difference in toxicity with VMP was that GI toxicity and peripheral neuropathy were increased. Thus, bortezomib is active in patients who are transplant eligible and non-transplant eligible and appears to have equal efficacy in younger and elderly patients. Further, the previous prognostic markers of 13q deletion by cytogenetics and t(4; 14) and t(14; 16) translocations do not appear to impact on the responses to bortezomib. Taken together, these data demonstrate the activity and importance of bortezomib for patients with relapsed refractory MM as well as newly diagnosed MM. Multiple trials are now ongoing, which use combinations of IMiDs with bortezomib and dexamethasone, and have overall response rates in newly diagnosed patients that approach 100%.
Bortezomib containing regimens for myeloma.
Use of Bortezomib in Patients with Renal Dysfunction
Renal failure is a common presenting feature of MM with about 20% of patients having decreased renal function. 36 There is a similar percentage of patients who develop renal failure in the course of their disease. Roussou and colleagues have examined the response to bortezomib-based regimens in patients with renal failure and MM. 37 They found in 8 of 20 patients that the renal failure was reversed, and that the median time for reversal was 17 days. Moreover 10 of the 20 patients had a 50% decrease in their serum creatinine. Further, 1 out of the 5 patients who was on hemodialysis became independent of dialysis on bortezomib. Overall, 85% of patients with an elevated serum creatinine had a decrease in their creatinine on bortezomib-based therapy. Further, the toxicity profile was similar in patients with renal failure as in patients without renal failure. Importantly, the overall response rate to treatment with bortezomib in patients with renal failure was similar to those without renal failure. Similar results have been reported by Ludwig et al. 38 Thus, bortezomib is an active agent for treating patients with renal failure and MM, and the dose of bortezomib does not have to be adjusted for decreased creatinine clearance.
Peripheral Neuropathy and Bortezomib
A major toxicity of bortezomib-based therapy is peripheral neuropathy. The pathogenesis of underlying peripheral neuropathy in patients receiving bortezomib is still not clear. What has been reported is that mitochondrial and endoplasmic reticulum damage occurs in nerve cells with bortezomib. Silverman and coworkers, using in vivo mouse models and the PC12 rat fetal cell line, have shown that a primary target for bortezomib-associated peripheral neuropathy is the dorsal root ganglion neuronal cell body. 39 Bortezomib caused chromatolysis of the dorsal root ganglion neurons and an accumulation of eosinophilic material in the cytoplasm. Argyriu and coworkers have recently reported a comprehensive review of the literature for bortezomib-induced peripheral neuropathy in patients with MM. 40 They found that metabolic changes have been reported with bortezomib resulting in mitochrondrial-mediated dysregulation of calcium homeostasis and neurotrophins. They further noted that bortezomib-induced neuropathy may be a proteasome inhibitor class effect that results in painful axonal sensory distal neuropathy. Neurophysiologic studies in patients receiving bortezomib for the treatment of MM have shown bortezomib-induced pain is associated with deficits in all three nerve fiber types. 41 The risk factors for bortezomib-induced neuropathy include cumulative dose and evidence of pre-existing neuropathy. 42 Current treatment for bortezomib-induced peripheral neuropathy includes dose reduction or holding the dose and restarting it at lower doses. 43 Dose and treatment reduction schedules have been suggested including reduced dose from 1.3 to 1.0 mg/m2 for patients that have grade 1 neuropathy with pain or grade 2 neuropathy, for grade 2 with pain or grade 3 or greater bortezomib therapy is held until the neuropathy resolves and is reinstituted at a dose of 0.7 mg/m2. However, patients who develop grade 4 neuropathy should have the drug discontinued. Importantly, prolonged administration of bortezomib was not significantly associated with increased incidences and severity of peripheral neuropathy in a study by Richardson et al. 43 However, there are currently no effective prophylactic treatments that prevent peripheral neuropathy in patients receiving bortezomib, and the treatment is predominantly symptomatic. 44
Thrombocytopenia Associated with Bortezomib Therapy
Thrombocytopenia is a common toxicity associated with bortezomib therapy.45,46 In most cases, it is readily reversible and is seen more frequently in patients who have lower platelet counts at the start of therapy. A mechanism suggested for bortezomib-induced thrombocytopenia is decreased budding of platelets from megakaryocytes, which is a NF-κB-dependent process.47,48 The suppression of platelet production is readily reversible within a few days and may actually return to normal levels between treatments. The current recommendation for treatment of patients on bortezomib with thrombocytopenia is to try to maintain the dose if possible and decrease dose as directed in the package insert.
In the APEX trial, thrombocytopenia occurred in 26% of patients on bortezomib but only 4% of patients had clinically significant bleeding. 48 Thrombocytopenia on bortezomib therapy was cyclic and associated with therapy and improved almost to baseline during the week off therapy.
Bortezomib as a Bone Active Agent in Myeloma
The proteasome pathway is an important regulator of OBL differentiation and proteasome inhibition can induce new bone formation in rodents by increasing BMP2 expression, a potent inducer of OBL differentiation and bone formation. 49 Similarly, Oyajobi et al have reported that bortezomib increase bone formation in mouse calvarial cultures. 50
Clinical trials have also suggested that bortezomib is a bone anabolic agent. Zangari et al conducted a retrospective analysis of three trials of bortezomib in patients with relapsed MM. 51 In all three trials, patients who had a partial response to bortezomib therapy experienced a transient increase in serum levels of alkaline phosphatase compared to non-responders.
Heider et al analyzed bone markers in relapsed MM patients enrolled in clinical trials using bortezomib and compared them to other agents. 52 Patients treated with bortezomib had increased levels of bone-specific alkaline phosphatase and osteocalcin, both markers of increased OBL activity. The increase in alkaline phosphatase and osteocalcin was observed in patients whose MM responded to bortezomib treatment as well as non-responders, but was not seen in the other treatment groups regardless of response. 52
Terpos et al have also reported that bortezomib monotherapy decreased the elevated bone resorption markers in patients with MM and decreased DKK1, an OBL inhibitor, and RANKL, an OCL stimulator, while increasing bone formation markers in patients with relapsed myeloma. 53 However, bortezomib did not affect OBL function in patients receiving other anti-myeloma therapy such as melphalan. 54 These results suggested that the effects of bortezomib were from direct effects on OBL and not a consequence of reducing MM tumor burden.
Giuliani et al recently reported the in vitro and in vivo effects of bortezomib in patients with MM. 55 In human OBL precursor cultures, bortezomib increased alkaline phosphatase and osteocalcin, OBL differentiation, as well as the OBL-specific transcription factors, RUNX2, and enhanced bone nodule formation. Bone marrow samples of patients responding to bortezomib had a significantly increased number of osteoblastic cells and osteocalcin staining compared to non-responders. These studies suggested that bortezomib stimulates OBL differentiation in patients whose MM responded to bortezomib, but that the persistence of MM cells inhibits the effects of bortezomib on OBLs. In support of this notion, Oyajobi et al have also reported that bortezomib increases bone formation in murine models of MM, and that the effects of bortezomib were inhibited by DKK1. Tian and coworkers reported that primary MM cells produce DKK1, which inhibits OBL differentiation. 56 These results suggest that suppression of MM cell DKK1 secretion by bortezomib is required for the bone anabolic effects of bortezomib to occur. Recently, Qiang et al reported that bortezomib can induce OBL differentiation by OBL progenitor from MM patients independently of Wnt signaling. They found that bortezomib stabilized B-catenin and its nuclear translocation in a Wnt-independent manner. 57
There may be other beneficial effects of increasing OBL activity in MM in addition to increasing bone formation. Yaccoby et al reported that OBLs inhibit the growth of MM cells. 58 Several groups have shown that increasing OBL activity through increased canonical Wnt signaling in the bone microenvironment can reduce tumor burden in animal models of myeloma.59,60 In contrast, Edwards et al reported that increased Wnt signaling can increase the growth of myeloma cells outside the bone microenvironment. 60 However, this has not been found in studies by Qiang et al 60 Therefore, the safety of enhanced Wnt signaling in MM remains to be determined.
Bortezomib can inhibit OCL differentiation through its capacity to inhibit NF-κB signaling.61,18 Terpos and colleagues and Giuliani et al 55 reported that bone resorption markers were decreased in patients receiving bortezomib. These results suggest that bortezomib suppresses OCL activity in vitro and in vivo. 54 In support of these findings, Pennisi et al have shown in a preclinical model of MM that bortezomib stimulates bone formation by both inhibiting osteoclastogenesis and stimulating new bone formation. 18
Summary
Bortezomib is the first-in-class proteasome antagonist approved for treatment of MM. It is active in newly diagnosed, relapsed and refractory patients and is now being used as a platform for combinations with new agents directed at a multiplicity of biologic targets in MM. In addition to its anti-myeloma effect, bortezomib can inhibit OCL formation through blocking NF-κB activity as well as stimulate OBL activity, at least transiently, in patients with MM. Potentially, combination of bortezomib with other agents, which can stimulate bone formation or block bone resorption, will further enhance the anti-myeloma effects of bortezomib and overcome the contribution of the tumor microenvironment in supporting MM growth.
Disclosure
The author reports no conflicts of interest.