
Abstract
Background
The present study investigated tumor proliferation in a tumor model using murine ovarian cancer cells with increased carbonyl reductase (CR) expression.
Methods
CR cDNA was transfected into murine T-Ag-MOSE ovarian cancer cells by lipofection. CR-transfected cells (CR induction group) or empty vector-treated cells (control group) were injected into the backs of 8-week-old nude mice at a concentration of 0.5 × 10 6 per 0.2 mL. Subsequent tumor proliferation in both groups was observed for 5 weeks.
Results
The control group showed an increase in tumor volume during the 5 weeks of observation. However, tumor volume in the CR induction group increased up to the second week but then decreased continuously until the fifth week of observation. The tumor growth curves for the two groups showed a significant difference (Mann-Whitney U test, P < 0.001). Histological and biochemical experiments were performed using tumor tissues isolated in the third week. Necrosis and inflammatory cell infiltration were noted for tumors in the CR induction group. Also, the number of apoptotic cells was significantly increased in the CR induction group compared with the control group (P < 0.001). Milk fat globule EGF factor 8, an “eat-me” signal for phagocytes such as macrophages, was expressed extensively in the tumor cytoplasm and interstitial cells of the CR induction group, and engulfment of apoptotic cells by macrophages was observed. Vascular endothelial growth factor expression in tumors was notably decreased in the CR induction group compared with the control group.
Conclusion
Increased necrosis due to engulfing of apoptotic cells by phagocytes attracted by increased milk fat globule EGF factor 8 was considered to be the mechanism of spontaneous tumor regression in the CR induction group.
Introduction
Ovarian cancer is the sixth most prevalent cancer among women globally, and its annual incidence is approximately 200,000. 1 Ovarian cancer accounts for one quarter of reproductive system neoplasms in women. It is often not detected until the advanced stages, namely stages III or I V, and is the most frequent cause of death among the gynecologic cancers. The current management of advanced epithelial ovarian cancer generally includes cytoreductive surgery followed by combination chemotherapy. A combination of paclitaxel with a platinum analog can achieve a remission rate of over 70% for ovarian cancer, 2 but recurrence remains high. The 5-year survival rate remains at about 50%, 3 and an estimated 130,000 deaths occur from ovarian cancer per year worldwide. 1 Because anticancer agents alone are not sufficiently effective, development of other agents, including molecularly targeted drugs, is necessary.
Clofibric acid, a peroxisome proliferator-activated receptor α ligand commonly used for the treatment of hyperlipidemia, may be one of the candidates for this. Our previous in vivo and in vitro studies have demonstrated that clofibric acid has an antitumor effect comparable with that of cisplatin against human ovarian cancer. 4 Because administration of clofibric acid in cancer-bearing animals induced a significant decrease in prostaglandin (PG) E2 concentration in serum and ascitic fluid and a remarkable increase in carbonyl reductase (CR) expression, we inferred that CR expression is the core factor in clofibric acid treatment, which induced a decrease in PGE2, then caused tumor shrinkage due to inhibition of angiogenesis and induction of apoptosis. 4 CR is present in a variety of organs, and its primary function is to control fatty acid metabolism. 5 CR is also referred to as PGE29 reductase, and its other important function is converting PGE2 to PGF2α. 6 In order to prove this hypothesis, we transfected ovarian cancer cells with an increased CR expression vector and investigated the effects of CR on the proliferation process in vivo.
Materials and Methods
Cell line and cell culture
T-Ag-MOSE, established by introduction of SV40 large T antigen DNA into C3H/He mouse ovarian surface epithelium and could be transfected cDNA with great efficiency, 7 was obtained from the Health Science Research Resources Bank (Osaka, Japan). Inoculation of T-Ag-MOSE cells onto the back region of nude mice can produce xenografted solid tumors. 4 The cell line was grown in Dulbecco's Modified Eagle Medium supplemented with 10% fetal calf serum at 37°C in a water-saturated atmosphere with 5% CO2/95% air.
Transfection
T-Ag-MOSE cells were plated into 24-well plates and cultured to 60%–80% confluency in Dulbecco's Modified Eagle Medium containing 10% fetal calf serum. Full-length cDNA coding for mouse CR1 was ligated into the pBabe Puro expression vector to generate a CR expression vector. 8 Then, 10 μg of the coding plasmid was transfected into T-Ag-MOSE cells using the GenePorter transfection reagent (Gene Therapy Systems, San Diego, CA) in accordance with the manufacturer's recommendations. The vector without CR1 cDNA was used as the control. Transfected cells were cultured in Dulbecco's Modified Eagle Medium with 10% fetal calf serum containing 1.5 mg/mL puromycin for 48 hours. We confirmed expression of the CR construct using Western blotting. 4
Animal Experimentation
The animal experiments were conducted in accordance with the guidelines for animal experimentation of Hirosaki University. Eight-week-old female Balb/c nu/nu mice were used in this study. All mice were group-housed in plastic cages with stainless-steel grid tops in an air-conditioned room in the Institute for Animal Experiments of Hirosaki University, on a 12-hour light-dark cycle, and fed with water and food ad libitum.
Cancer-bearing mouse model
CR-transfected or control T-Ag-MOSE cells (0.5 × 10 6 cells) in 0.2 mL of Dulbecco's Modified Eagle Medium were inoculated subcutaneously on the back region of the nude mice. All the mice were numbered, housed separately, and examined twice weekly for tumor development. The tumor dimensions were measured twice weekly using a caliper, and tumor volume was calculated using the equation V (mm3) = A × B 2 /2, where A is the largest diameter and B is the smallest diameter. 9 Tumors were isolated from selected mice of both groups in the third week of observation, and examined pathologically and biochemically.
Apoptosis
Apoptosis was measured on tissue sections by the terminal deoxyribonucleotidyl transferase (TdT)- mediated dUTP-biotin nick end labeling (TUNEL) assay as described by Gavrieli et al 10 with some modifications. Briefly, 6 μm sections were stripped from proteins by incubation with 10 mg/mL proteinase K for 15 minutes and immersed in H2O2 0.3% in methanol for 15 minutes to block the endogenous peroxidase. The sections were then incubated in TdT buffer mixture (200 mM potassium cacodylate, 25 mM Tris-HCl, pH 6.5, 0.25 mg/mL bovine serum albumin, 1 mM CoCl2, 0.01 mM biotin-dUTP, 520 U/mL TdT) at 37°C for one hour. After rinsing in phosphate-buffered saline, the sections were exposed to avidin-biotin-peroxidase complex (Vecta Laboratories, Burlingame, CA) at 37°C for 30 minutes. Cells undergoing apoptosis were visualized with diaminobenzidine (Sigma-Aldrich, St Louis, MO). TUNEL-positive cells per 0.75 mm2 were counted under a high-power field. Three fields were counted per tumor, and the average was taken as the frequency of apoptotic cells of each tumor.
Immunohistochemistry
Six micron sections of formalin-fixed and paraffin-embedded tissue specimens were stained by an established method, as described previously. 11 Sections were incubated with antibodies specific for CR 12 and MFG-E8 (Santa Cruz Biotechnology, Santa Cruz, CA) for 2 hours. Slides were incubated with appropriate biotinylated species-specific secondary antibodies for 30 minutes and then exposed to avidin-biotin-peroxidase complex. Sections were treated with 0.02% diaminobenzidine as a chromogen, and counterstained with hematoxylin.
Western blot analysis
Cell lysates (50 μg protein) were prepared from tumor tissues, electrophoresed through a 12.5% sodium dodecyl sulfate polyacrylamide gel, and blotted as described previously. 13 The protein concentration was determined using Bradford's method. The blots were probed with the following diluted antibodies for 2 hours: CR at 1:1000, cleaved caspase-3 (Santa Cruz Biotechnology) at 1:1000, milk fat globule EGF factor 8 (MFG-E8) at 1:1000, mouse vascular endothelial growth factor (VEGF) 164 (R&D Systems, Minneapolis, MN) at 1:500, and β-actin (Sigma-Aldrich) at 1:2000. The membranes were then incubated for one hour with the appropriate biotinylated secondary antibodies, transferred to avidin-biotin-peroxidase complex reagent, and incubated in this solution for 30 minutes. Diaminobenzidine was used as a substrate. Quantification of the results was performed by scanning the membrane with Photoshop software (version 5.5, Adobe Systems) followed by densitometry with the public domain software, NIH Image, version 1.62.
Double fluorescence staining for detection of apoptotic cells and macrophages
Six micron sections were stripped from proteins by incubation with 10 mg/mL proteinase K for 15 minutes. The sections were then incubated in TdT mixture buffer at 37°C for one hour and in TB buffer at room temperature for 15 minutes, and exposed to the blocking solution at room temperature for 10 minutes to block the nonspecific reaction. After being rinsed in phosphate-buffered saline, the sections were incubated with the antibody specific for mouse macrophage (Novocastra Laboratories, Newcastle upon Tyne, UK) at room temperature for 30 minutes and then with FITC-labeled streptavidin at 37°C for 30 minutes. After being rinsed in phosphate-buffered saline, the sections were incubated with antimouse IgG Alexa Fluor 555 (Invitrogen) at room temperature for one hour. The sections were then counterstained with DNA dye 4′,6-diamidino-2-phenylindole (DAPI) and the results were viewed using fluorescence microscopy. Here, proteinase K, TdT mixture buffer, TB buffer, the blocking solution, and FITC-labeled streptavidin in the Mebstain apoptosis kit II (MBL International, Woburn, MA) were used.
Statistical Analysis
Differences in tumor volume between the control and CR induction groups were analyzed using the one-tailed Student's t-test. The significance of the difference in the frequency of apoptotic cells and expression of MFG-E8 between the two groups was also carried out using the Student's t-test. A result was deemed to be statistically significant at P < 0.05.
Results
Spontaneous regression of tumors with increased CR expression
Tumor volume was measured in five mice from each group. Tumor volume continued to increase in the mice in which T-Ag-MOSE cells without CR had been injected (control group, Figure 1). However, it started to decrease spontaneously after the second week of observation in mice into which CR-transfected T-Ag-MOSE cells had been injected (CR induction group, Figure 1). A significant difference was observed in tumor volume between the control group and the CR induction group from the second week of observation (Fig. 1). The experiment was repeated three times, producing similar results. Figure 2 shows hematoxylineosin staining of tumor tissues isolated in the third week of observation. Tumor cells were densely packed in tumor tissues of the control group, but were sparse in tissues of the CR induction group. Large portions of cells were also necrotic in the CR induction group. Coagulative necrosis with inflammatory cells was observed around the tumor tissue (Fig. 2B).

Changes in tumor volume in control (n = 5) and carbonyl reductase induction (n = 5) groups.
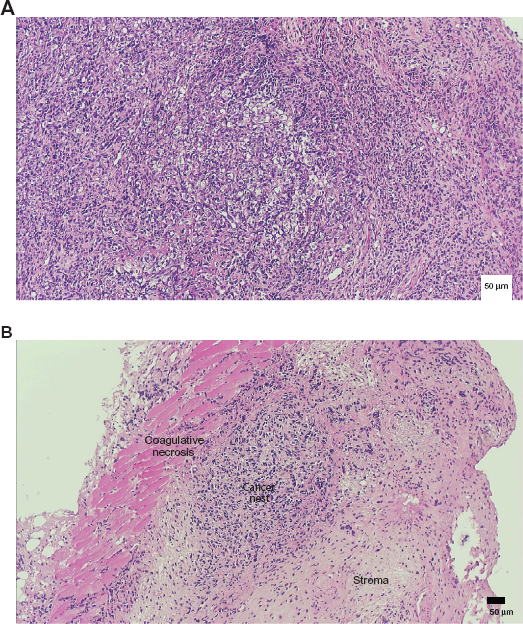
Hematoxylin-eosin staining of tumor tissue isolated in the third week of observation. (
VEGF expression in CR induction group tumors
Expression of CR is known to be negatively correlated with that of VEGF, an angiogenesis factor. 4 An analysis of VEGF expression in tumors of both the control group and the CR induction group revealed a notable decrease in VEGF in the CR induction group (Fig. 3). However, there was no significant difference in microvessel density identified with the anti-CD31 antibody using immunostaining in tumor tissues between the CR induction and control groups (data not shown). Thus, there has not been enough evidence to show a decrease in angiogenesis in tumors of the CR induction group.

Relationship between vascular endothelial growth factor and carbonyl reductase expression in carbonyl reductase induction group and control group.
Comparison of apoptosis in tumor tissue
The expression of apoptotic cells in tumors was evaluated using the TUNEL method in the third week of observation. To evaluate the extent of apoptosis in tumor tissue, apoptotic cells were stained by the TUNEL method, and TUNEL-positive cells per 0.75 mm2 were counted in a high power field. Frequency of the incidence of TUNEL-positive cells (number/mm2) was 10.1 ± 3.0 for the control group (n = 5) and 28.7 ± 4.1 for the CR induction group (n = 5, Figure 4A). The incidence of apoptotic cells was significantly higher in the CR induction group than in the control group (Fig. 4A, P < 0.001). To examine whether the expression of caspase-3, which becomes activated during apoptosis, in tumors in the third week of observation is altered, caspase-3 expression was evaluated by Western blotting. This analysis revealed that caspase-3 expression increased by 3.2-fold in CR induction tumors compared with the controls (Fig. 4B).
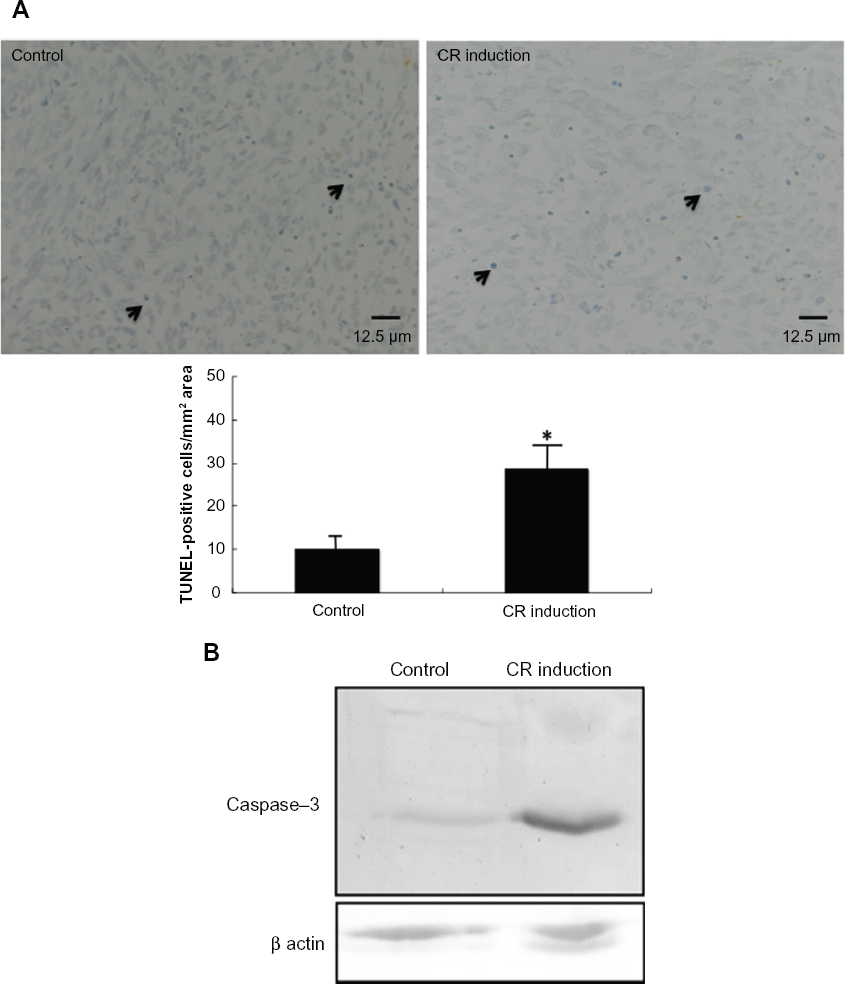
(
Comparison of eat-me signal expression in tumor tissue
Eat-me signals on apoptotic cells are known to attract inflammatory cells such as macrophages at the time of apoptosis. MFG-E8, an eat-me signal, binds to apoptotic cells and attracts macrophages. An assay of MFG-E8 expression in tumors isolated from the control group and from the CR induction group revealed significantly higher MFG-E8 expression in tumors in the CR induction group (Fig. 5A). In addition, immunohistochemical staining for investigation of the localization and expression of MFG-E8 showed that MFG-E8 was absent in tumors from the control group but was extensively expressed in the tumor cytoplasm and interstitial cells in tumors of the CR induction group (Fig. 5B). Double fluorescence staining of tumors isolated from the CR induction group showed engulfment of apoptotic cells by macrophages (Fig. 6). These findings suggested that apoptosis occurred frequently in tumors of the CR induction group and that necrosis was induced due to phagocytosis (phagocytes such as macrophages are attracted to MFG-E8 and capture cancer cells).
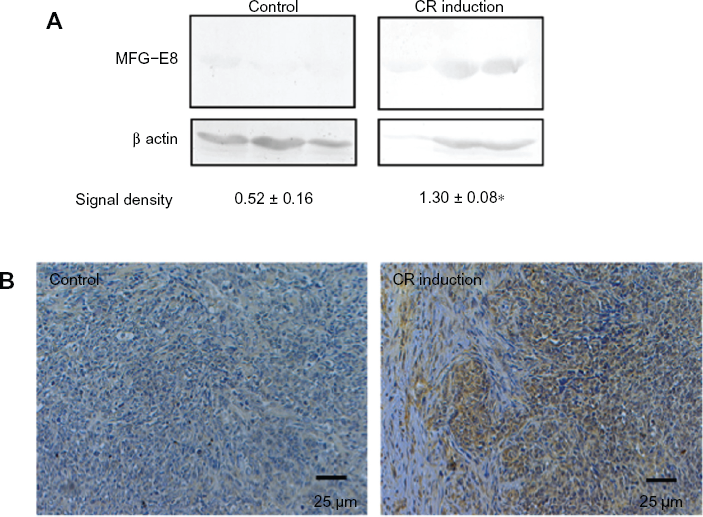
(

Double fluorescence staining of apoptotic cells and macrophages.
Discussion
The findings of the present study clearly demonstrate that tumors containing cells with increased CR expression undergo spontaneous regression. This spontaneous regression was considered to occur as a result of apoptosis as well as necrosis. As shown in Figure 5, MFG-E8 expression was significantly increased in tumors containing cells with increased CR expression, suggesting that the notable increase in necrosis in tumors containing cells with increased CR expression was induced by engulfment of apoptotic cells by phagocytes, such as macrophages.
It is generally accepted that MFG-E8 is secreted from cells that have synthesized it, and then binds to the surface of apoptotic cells. 14 However, the present study raises the issue of whether the MFG-E8 detected is synthesized and secreted by CR-transfected cells or by the host's leukocytes stimulated by CR-expressing cancer cells, given that localization of CR, MFG-E8, and leukocytes was not confirmed in consecutive sections. Localization of MFG-E8 seen in Figure 5 demonstrates that MFG-E8 is present in both cancer cells and interstitial cells, indicating either or both could be possible. Wakui et al reported increased apoptosis and occurrence of necrosis in ovarian cancer tissue treated with photodynamic therapy, and demonstrated immunohistochemically that MFG-E8 expression was localized in the nuclei and cell membranes of cancer cells. 9 These findings support the hypothesis that MFG-E8 synthesized in cancer cells is secreted into the surrounding tissues.
There remains the issue of the difference in host response between tumors containing cells with high CR expression and tumors containing control cells. The results of the present study seem to suggest that this is because apoptosis in tumors containing CR- expressing cells induce MFG-E8 secretion that, in turn, induces phagocytosis by attracting macrophages and other phagocytes. However, necrosis of CR- expressing cells may also be due to secretion of a factor similar to tumor necrosis factor alpha (TNFα) by macrophages that are attracted to an eat-me signal-like substance secreted by CR-expressing cells. TNFα is a homotrimer of 17 kDa protein subunits, and is a potent mediator of diseases and metabolic functions. 15 TNFα induces necrosis in cancer cells in vivo, and has cytolytic effects on cancer cells in vitro. Maslinic acid, which is both an antioxidant and anti- inflammatory substance, has been reported to inhibit pancreatic cancer cell growth significantly by activating TNFα. 16 Maslinic acid achieves this by significantly decreasing the expression of genes associated with the NF-κB signal pathway, such as cyclin-D1, cyclo-oxygenase-2, survivin, matrix metalloproteinase-9, and VEGF, that are involved with cell proliferation, antiapoptosis, infiltration, and angiogenesis. 10 Therefore, a thorough investigation of the TNFα signaling pathway may be necessary in cells into which CR, which is also both an antioxidant and anti- inflammatory substance, have been transfected.
Recently, the exactly opposite evidence concerning a crucial role of MFG-E8 in tumor progression have been reported.17,18 While MFG-E8 promotes VEGF-dependent angiogenesis, 17 MFG-E8 regulates the antitumor effect by efficient phagocytosis of apoptotic cells via induced granulocyte-macrophage colony stimulating factor activity. 18 This complicated evidence suggests pleiotropic roles for MFG-E8. Indeed, in breast cancers, MFG-E8 is expressed in triple-negative [estrogen receptor (–), progesterone receptor (–), erbB2 (–)] breast cancers as a target gene of the p63 pathway, whereas it exhibits a suppressive function in estrogen receptor (+) and erbB2 (+) breast cancers. 19 In this study, increased MFG-E8 in CR-transfected tumors was related to engulfment of apoptotic cells by phagocytes, such as macrophages, resulting in tumor regression. However, MFG-E8 expression did not correlate with either VEGF expression or angiogenesis. Such different functions of MFG-E8 warrant continued study of whether MFG-E8 is a tumor promoter or a suppressor.
In a previous study, we found that clofibric acid, a peroxisome proliferator-activated receptor α ligand, exerts an antitumor effect, partly by increasing CR expression in ovarian cancer tissue. 4 We also demonstrated that increased CR expression in tumors achieved tumor regression by inhibiting angiogenesis and inducing apoptosis. 4 The present study also showed that VEGF expression in tumors containing CR-transfected cells was decreased in comparison with that in control tumors. However, there was no significant difference in microvessel density in tumor tissue between the two groups. Thus, there is enough evidence to show a decrease in angiogenesis in tumors of the CR induction group. Further study would be necessary to elucidate the inverse relationship between expressions of VEGF and CR in spontaneous tumor regression.
Clofibric acid is the only drug known to increase CR expression in tumors. The findings of the present study have strengthened the evidence that CR expression in a tumor induces an antitumor effect. This suggests clofibric acid is a promising substance for molecularly targeted drugs that could increase CR expression in tumor tissue. Clinical trials should be urgently planned to establish the antitumor effect of clofibric acid, a primary component of antihyperlipidemic agents.
Disclosures
Author(s) have provided signed confirmations to the publisher of their compliance with all applicable legal and ethical obligations in respect to declaration of conflicts of interest, funding, authorship and contributorship, and compliance with ethical requirements in respect to treatment of human and animal test subjects. If this article contains identifiable human subject(s) author(s) were required to supply signed patient consent prior to publication. Author(s) have confirmed that the published article is unique and not under consideration nor published by any other publication and that they have consent to reproduce any copyrighted material. The peer reviewers declared no conflicts of interest.
Footnotes
Acknowledgments
This study was supported in part by a Grant-in Aid for Cancer Research (20591935) from the Ministry of Education, Science and Culture of Japan and by the Karoji Memorial Fund of the Hirosaki University Graduate School of Medicine.