
Abstract
The arterial baroreflex system can be divided into the neural arc, from pressure input to efferent sympathetic nerve activity (SNA), and the peripheral arc, from SNA to arterial pressure (AP). Plotting the neural and peripheral arcs on a pressure–SNA plane yields a baroreflex equilibrium diagram. We examined the effects of vagotomy on the open-loop static characteristics of the carotid sinus baroreflex in normal control rats (NC, n = 10) and rats with heart failure after myocardial infarction (MI, n = 10). In the NC group, vagotomy shifted the neural arc toward higher SNA and decreased the slope of the peripheral arc. Consequently, the operating-point SNA increased without a significant change in the operating-point AP on the baroreflex equilibrium diagram. These vagotomy-induced effects were not observed in the MI group, suggesting a loss of vagal modulation of the carotid sinus baroreflex function in heart failure.
Keywords
Introduction
The arterial baroreflex is an important negative feedback system that stabilizes arterial pressure (AP) during daily activity. The sympathetic limb of the arterial baroreflex system can be divided into two principal subsystems: neural arc (controller) and peripheral arc (effector).1–3 The neural arc determines the efferent sympathetic nerve activity (SNA) in response to a baroreceptor pressure input, and the peripheral arc determines AP at a given SNA. Under normal physiological conditions, AP serves as an input to the neural arc as well as an output from the peripheral arc. SNA serves as an output from the neural arc as well as an input to the peripheral arc. Hence, it is difficult to quantify the input–output relationships of the two arcs under baroreflex closed-loop conditions. 4 To circumvent the problem, we and others have employed a framework of open-loop systems analysis by isolating the baroreceptor regions.5–8
In open-loop systems analysis relating to the carotid sinus baroreflex, the vagal nerves together with the aortic depressor nerves are often sectioned in order to focus only on the reflex responses mediated by the carotid sinus baroreflex. 8 This does not mean that the vagal nerves can be ignored in understanding sympathetic AP regulation. Pharmacological activation 9 or electrical stimulation 10 of the vagal afferents shifts the neural arc toward lower SNA, suggesting that vagotomy may exert an opposite influence on the neural arc. According to a study by Wang et al. 7 , however, vagotomy does not significantly alter the renal SNA or AP response to carotid sinus pressure (CSP) in anesthetized dogs. Their results suggest that vagal afferents may not significantly modify carotid sinus baroreflex function under anesthetized conditions. Nevertheless, reinvestigating the effect of vagotomy on open-loop carotid sinus baroreflex function in rats would be worthwhile, as this species is increasingly used in cardiovascular research. 8
In cardiovascular research, heart failure is one of the important study topics. Autonomic imbalance with sympathetic excitation and vagal withdrawal has been shown to aggravate heart failure. 11 Although the mechanisms of autonomic imbalance are not completely understood, impairment of the arterial and cardiopulmonary baroreflexes is a major contributing factor.12,13 While we have also demonstrated that the open-loop static characteristics of the carotid sinus baroreflex are significantly depressed in a rat model of chronic heart failure following myocardial infarction (MI), 14 the experiment was conducted only under vagotomized conditions. Hence, the purpose of this study was to examine the carotid sinus baroreflex function before and after vagotomy in normal control (NC) rats, and compare the results with those obtained from rats with post-MI heart failure. This study differs from the previous study by Wang et al. 7 with respect to the animal species (rats versus dogs) and the model of heart failure (post-MI versus pacing-induced heart failure). In addition, hemodynamic changes were monitored over time after vagotomy. We hypothesized that the effect of vagotomy on carotid sinus baroreflex function, if any, is significantly suppressed in MI rats.
Materials and Methods
Animal care was provided in strict accordance with the Guiding Principles for the Care and Use of Animals in the Field of Physiological Sciences; it was approved by the Physiological Society of Japan. All protocols were reviewed and approved by the Animal Subject Committee of the National Cerebral and Cardiovascular Center.
Surgical Preparation
The left coronary artery was permanently ligated under halothane anesthesia to induce a large MI in 8-week-old male Sprague-Dawley rats according to a previously established procedure.15–17 Eight weeks after MI, surviving rats underwent an acute baroreflex experiment described below. Among 17 MI rats that had survived, two died from cardiopulmonary insufficiency during the acute experiment, three did not allow a reliable estimation of baroreflex function due to unstable hemodynamics or very weak baroreflex responses, and two showed no significant ventricular remodeling in postmortem examination (normalized biventricular weight <2.5 g/kg; see Discussion section for exclusion criterion). Eventually, data obtained from 10 MI rats were compared with those obtained from 10 NC rats.
In the acute baroreflex experiment, each rat was anesthetized with an intraperitoneal injection (2 mL/kg) of a mixture of urethane (250 mg/mL) and α-chloralose (40 mg/mL), and mechanically ventilated with oxygen-supplemented room air. An appropriate level of anesthesia was maintained by continuous intravenous infusion of the above anesthetic mixture. Central venous pressure was measured from a venous catheter inserted into the left femoral vein and advanced to the inferior vena cava. Fluid balance was maintained by intravenous infusion of lactated Ringer's solution (6 mL kg−1 h−1). An arterial catheter was inserted into the right femoral artery to measure AP and heart rate (HR). A heating pad was used to maintain the body temperature of the rat at approximately 38 °C.
A postganglionic branch of the splanchnic sympathetic nerve was exposed through a left flank incision. A pair of stainless steel wire electrodes (AS633; Cooner Wire) was fixed to the nerve using silicone glue (Kwik-Sil; World Precision Instruments). Preamplified nerve signals were band-pass filtered at 150-1,000 Hz, and then full-wave rectified and low-pass filtered at 30 Hz to quantify SNA. A ganglionic blocker hexamethonium bromide (60 mg/kg, bolus) was injected intravenously at the end of the experiment to confirm the disappearance of SNA.
Carotid sinus baroreceptor regions were isolated from systemic circulation.18,19 The isolated carotid sinuses were filled with warmed Ringer's solution through catheters inserted into the common carotid arteries. CSP was controlled using a servo-controlled piston pump. Heparin sodium (100 U/kg) was given intravenously to prevent blood coagulation.
The right and left aortic depressor nerves were identified as far as possible by recording AP pulse synchronous activities and were sectioned. A 5-0 silk suture snares were prepared around the right and left cervical vagal nerves for later vagotomy.
Protocol
After the surgical procedures were completed, hemodynamics were allowed to stabilize for more than 30 minutes. Thereafter, CSP was decreased to 60 mmHg for 4 minutes, and then increased staircasewise from 60 to 180 mmHg in increments of 20 mmHg per minute.14,16 Seven input sequences (S1 through S7) were implemented, and bilateral cervical vagotomy was performed one minute after the completion of S3 (Fig. 1).

Typical experimental time series of carotid sinus pressure (CSP), sympathetic nerve activity (SNA), arterial pressure (AP), and heart rate (HR) obtained from a normal control (NC) rat and a rat with chronic heart failure after myocardial infarction (MI). Gray and black lines in SNA represent 10-Hz resampled signals and 2-s moving averaged signals, respectively. Gray and black lines in AP and HR represent 200-Hz sampled signals and 2-s moving averaged signals, respectively. Staircase-wise CSP input was repeated throughout the protocol. After three input sequences under baseline condition (S1 through S3), bilateral cervical vagal nerves were sectioned (vagotomy). Thereafter, four input sequences were observed (S4 through S7). At the end of the experiment, hexamethonium bromide (C6) was injected intravenously to confirm the disappearance of SNA.
Data Analysis
Data were sampled at 200 Hz using a 16-bit analog-to-digital converter. For each input sequence, SNA, AP, and HR data were averaged during the last 10 seconds of each CSP level to assess the carotid sinus baroreflex function. The noise level of SNA measured after hexamethonium administration was considered to be zero. As the absolute amplitude of SNA varied among animals depending on recording conditions, SNA was normalized in each animal, so that the averaged value of SNA at the CSP level of 60 mmHg for the sequences before vagotomy (S1 through S3) was 100%.
The sigmoidal input–output relationships of AP, HR, and SNA versus CSP were quantified using the following four-parameter logistic function8,20:
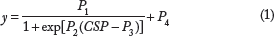
where y denotes the output; P1 the response range; P2 the slope coefficient; P3 the midpoint pressure of the sigmoid curve on the CSP axis; and P4 the minimum value of the sigmoid curve. The maximum gain or the maximum slope is calculated from Gmax = P1 X P2/4. Open-loop static characteristics of the peripheral arc approximated a straight line 8 and the slope and intercept were estimated.
Statistical Analysis
All data are presented as mean and standard error. Differences in baseline hemodynamics between the NC and MI groups were tested using the unpaired t-test. The overall effects of MI and vagotomy on the parameters of baroreflex static characteristics were examined, taking into consideration the time effect over the seven input sequences, using multiple linear regression: 21

where b0 is an estimate of a given parameter, p, before vagotomy in the NC group; bMI is a coefficient for the effect of MI; DMI is a dummy variable encoding the two groups (DMI = 0 for NC and 1 for MI); bVX is a coefficient for the effect of vagotomy; DVX is a dummy variable encoding vagotomy (DVX = 0 for pre-vagotomy and 1 for post-vagotomy);
Note that a significant positive/negative bMI indicates that the parameter is greater/smaller in the MI than in the NC group. A significant positive/negative bVX indicates that vagotomy increases/decreases the parameter as an overall effect. A significant positive/negative bTime indicates an increasing/decreasing trend of the parameter across the seven input sequences.
When any of bVX, bINT, and bTime was significantly different from zero, post-hoc analysis was performed within the NC or MI group using Dunnett's test. Changes in the parameter values were tested versus the third input sequence (S3). In all statistical analyses, differences were considered to be significant at P < 0.05.
Results
At necropsy of the MI rats, macroscopic inspection revealed that most of the left ventricular free wall was replaced by membrane-like scar tissue. The scar tissue occupied 40-50% of the left ventricular circumference in a section cut approximately midway between the base and apex. The biventricular weight was significantly increased in the MI group compared to that of the NC group (Table 1). At baseline (before the carotid sinuses were isolated), the central venous pressure was higher and HR was lower in the MI than in the NC group, while the mean AP did not differ significantly between the two groups (Table 1).
Biventricular weight and baseline hemodynamics.

Figure 1 shows typical time series of CSP, SNA, AP, and HR obtained from an NC rat (A) and an MI rat (B). In both rats, the staircase-wise increase in CSP decreased SNA, AP, and HR. Vagotomy was performed 1 minute after the completion of S3. In the MI rat, an artifact in SNA was observed due to vagal nerve manipulation at the time of vagotomy. In both the NC and MI rats, SNA disappeared after hexamethonium (C6) administration.
Figure 2 compares the baroreflex static characteristics of the NC (blue) and MI (red) rats estimated from the data shown in Figure 1. In the NC rat, vagotomy increased SNA and HR throughout the four input sequences after vagotomy (S4-S7). Although AP also increased during the first input sequence after vagotomy (S4), the increase subsided during the last three input sequences (S5-S7). In the MI rat, the magnitudes of SNA, AP, and HR responses to CSP were smaller than those observed in the NC rat. Vagotomy did not change the SNA or AP response significantly in the MI rat.

Open-loop static characteristics of the total reflex arc (top row), heart rate (HR) control (second row), neural arc (third row), peripheral arc (fourth row), and baroreflex equilibrium diagram (bottom row) derived from the same data shown in Figure 1. The plots derived from the normal control (NC) rat are shown in blue, and those from the rat with myocardial infarction (MI) are shown in red.
The results of multiple linear regression are summarized in Table 2. Open-loop static characteristics of the carotid sinus baroreflex averaged for the NC and MI groups are illustrated in Figures 3 and 4, respectively. The logistic function and the regression line fitted for the data in S3 are shown in light blue, and they are duplicated in the plots of the other input sequences for comparison. The red lines in Figure 3 indicate parameters showing significant differences compared with S3 by post-hoc Dunnett's test.
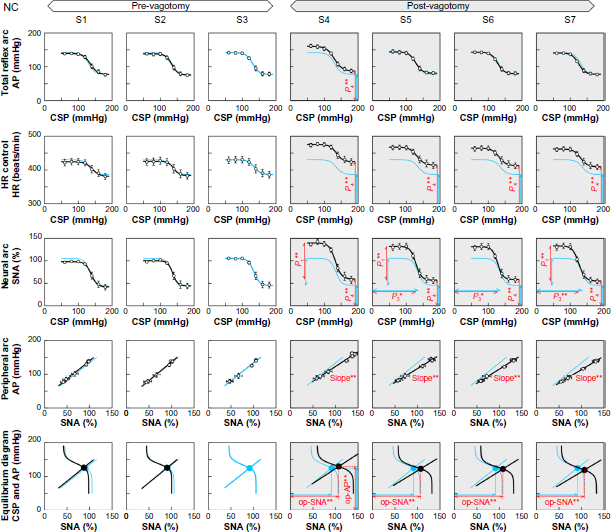
Open-loop static characteristics of the total reflex arc (top row), heart rate (HR) control (second row), neural arc (third row), peripheral arc (fourth row), and baroreflex equilibrium diagram (bottom row) averaged for the normal control (NC) group (n = 10). The characteristics of the total reflex arc, HR control, and neural arc approximate an inverse sigmoid curve. The characteristics of the peripheral arc approximate a straight line. For each row, the sigmoid curve fitted for sequence S3 is duplicated in all other sequences for comparison. Significant changes in parameters are shown in red lines.

Open-loop static characteristics of the total reflex arc (top row), heart rate (HR) control (second row), neural arc (third row), peripheral arc (fourth row), and baroreflex equilibrium diagram (bottom row) averaged for the myocardial infarction (MI) group (n = 10). The characteristics of the total reflex arc, HR control, and neural arc approximated an inverse sigmoid curve. The characteristics of the peripheral arc approximated a straight line. For each row, the sigmoid curve fitted for sequence S3 is duplicated in all other sequences for comparison. While the open-loop static characteristics in S4 apparently deviated slightly from those in S3, Dunnett's test detected no significant differences in the fitted parameters in S4 compared to those in S3.
Results of multiple linear regression analyses on the parameters of carotid sinus baroreflex function.

indicate P < 0.05
P < 0.01, respectively.
For the total reflex arc, the significant positive/negative bMI values (Table 2) indicated that the response range (P1) was narrower, the slope coefficient (P2) was greater, the midpoint pressure (P3) was lower, the minimum AP (P4) was higher, and the maximum gain (Gmax) was lower in the MI than in the NC group. The significant positive bVX for P4 indicated that vagotomy increased minimum AP as an overall effect. The significant negative bTime for P4 indicated a trend of decrease in minimum AP, but the magnitude was small (-2.5 mmHg per sequence). A within-group analysis in the NC group (Fig. 3, top row) revealed that minimum AP in S4 was significantly higher than that in S3. In contrast, minimum AP did not change significantly across the input sequences in the MI group (Fig. 4, top row).
For HR control, the bMI values (Table 2) indicated that the response range was smaller, the slope coefficient was greater, the midpoint input pressure was lower, and the minimum HR was lower in the MI than in the NC group. Vagotomy increased the response range and minimum HR as an overall effect. These effects of vagotomy were significantly attenuated in the MI group because bINT was significant and negative for P1 and P4. A small decreasing trend in minimum HR was observed. A within-group analysis in the NC group (Fig. 3, second row) revealed that minimum HR was significantly higher in sequences S4 through S7 than in S3. In contrast, minimum HR did not change significantly in the MI group (Fig. 4, second row).
For the neural arc, the bMI values (Table 2) indicated that the SNA response range was narrower, the slope coefficient was greater, the midpoint input pressure was lower, and the minimum SNA was higher in the MI than in the NC group. Vagotomy increased the response range, minimum SNA, and maximum slope. The effects of vagotomy on the response range and minimum SNA in the MI group were not as great as those in the NC group because bINT were significant and negative for P1 and P4. A within-group analysis in the NC group (Fig. 3, third row) revealed that the SNA response range was greater in sequences S4, S5, and S7 than in S3; the midpoint pressure was lower in sequences S5 through S7 than in S3; and the minimum SNA was higher in sequences S4 through S7 than in S3. In contrast, these parameters did not change significantly in the MI group (Fig. 4, third row).
For the peripheral arc, the bMI values (Table 2) showed that the slope was significantly smaller and the AP-intercept was significantly higher in the MI than in the NC group. Vagotomy significantly reduced the slope but increased the AP-intercept. The significant positive bINT for the slope (0.20 mmHg/%) had an absolute value similar to bVX (-0.23 mmHg/%), indicating that vagotomy did not reduce the slope in the MI group. There was a small trend of decrease in the AP-intercept (-2.4 mmHg per sequence). A within-group analysis in the NC group (Fig. 3, fourth row) revealed that the slope was significantly smaller in sequences S4 through S7 than in S3. No significant changes in the slope of the peripheral arc were detected within the MI group (Fig. 4, fourth row).
Baroreflex equilibrium diagrams can be obtained by plotting the neural and peripheral arcs on a pressure–SNA plane.2,3 The intersection between the two arcs provides the operating point of the carotid sinus baroreflex. Operating-point SNA and AP were lower in the MI than in the NC group (Table 2). Vagotomy increased the operating-point SNA, AP, and gain. Since bINT were significant and negative with absolute values comparable to bVX for operating-point SNA and gain, the effects of vagotomy on these parameters were almost canceled in the MI group. A within-group analysis in the NC group (Fig. 3, bottom row) indicated that operating-point SNA was significantly higher in sequences S4 through S7 than in S3, and operating-point AP was significantly higher in S4 than in S3. These parameters did not change significantly within the MI group (Fig. 4, bottom row).
Discussion
The present study examined the effects of vagotomy on the open-loop static characteristics of the carotid sinus baroreflex in NC and MI rats. Vagotomy significantly increased AP, HR, and SNA in the NC group. While AP was significantly elevated only during the first input sequence after vagotomy (S4), SNA and HR remained elevated until the end of the protocol. Vagotomy also significantly decreased the slope of the peripheral arc in the NC group. These effects of vagotomy on the carotid sinus baroreflex function were blunted in the MI group.
Characteristics of carotid sinus baroreflex function in MI rats
A previous study using Wistar rats indicated that the left ventricular weight normalized to body weight decreased after MI during the period of scar formation and then recovered toward the pre-MI value by 106 days after MI due to compensatory hypertrophy of the remaining myocardium. 22 The right ventricle showed significant hypertrophy by 106 days after MI only in rats with large MI. Consequently, normalized biventricular weight increased in rats with large MI. We have also found that significant biventricular remodeling occurs only in rats with large MI, 15 and adopted normalized biventricular weight of 2.5 g/kg (approximately mean plus 2 standard deviation of the NC group) as a criterion for successful creation of large MI. 14
The response ranges of SNA and AP were smaller in the MI than in the NC group (Table 2, negative bMI for P1) confirming that the carotid sinus baroreflex is depressed in the MI rats. 14 The minimum AP of the total reflex arc was significantly higher in the MI than in the NC group (positive bMI on P4), which is consistent with a canine model of pacing-induced heart failure. 7 Insufficient sympathetic suppression during high CSP and fluid retention associated with low cardiac output likely increased the minimum AP in the MI group.
The slope coefficient of the total reflex arc was greater (positive bMI for P2), but the maximum gain was smaller (negative bMI for Gmax) in the MI than in the NC group. Since the slope coefficient is inversely related to the size of the input range of the sigmoid curve, 20 the greater slope coefficient implies a narrower input range of AP regulation in the MI group. The reduced input range may increase the vulnerability of AP to exogenous disturbance. For instance, noncompliance with salt and water restriction easily leads to acute decompensation even in patients with stable heart failure. 23 Baroreflex failure impairs volume tolerance and increases the risk of pulmonary edema even in the absence of left ventricular dysfunction. 24
Effects of vagotomy on the neural arc and total reflex arc
The vagal afferents transmit signals from cardiopulmonary baroreceptors that respond to volume expansion. 25 In a study by Wang et al. 7 , however, vagotomy did not affect the CSP–renal SNA or CSP–AP relationship in anesthetized dogs with or without pacing-induced heart failure, suggesting that vagal afferent function cannot be observed under anesthetized conditions unless vagal afferents are experimentally stimulated. On the other hand, in a study by DiBona and Sawin, 13 vagotomy immediately increased renal SNA and AP in anesthetized rats, and the magnitude of SNA and AP elevations was attenuated in rats with MI compared with sham-operated rats. While the contradiction between these two studies may be attributed to species difference, the present results suggest that time-dependent hemodynamic changes after vagotomy may partly explain the discrepancy. Vagotomy increased SNA and AP during the first input sequence after vagotomy in the NC group (Fig. 3, S4). While the increase in SNA persisted until the last input sequence, the increase in AP subsided in sequences S5 through S7. This time-dependent recovery of AP may be a reason for no significant effect of vagotomy on the CSP–AP relationship reported by Wang et al. 7 because they waited 30 minutes after vagotomy to assess the baroreflex function. An increase in AP after vagotomy and its time-dependent recovery toward baseline AP have also been reported in anesthetized dogs with or without carotid sinus baroreceptor denervation. 26 On the other hand, sustained SNA elevation after vagotomy in the present study contradicts the lack of significant effect of vagotomy on the CSP–renal SNA relationship reported by Wang et al. 7 Further studies are needed to reconcile the diverse effects of vagotomy on the neural arc.
In the present study, vagotomy shifted the neural arc toward higher SNA in the NC group (positive bVX for P1 and P4), probably reflecting the elimination of vagally mediated tonic sympathoinhibitory effect. Given that volume expansion induces cardiopulmonary baroreflex responses 25 and that the MI rats had higher central venous pressure (Table 1), the vagally mediated tonic sympathoinhibitory effect was expected to be enhanced in the MI rats. However, vagotomy did not increase SNA in the MI rats (Fig. 4), suggesting impairment of the vagally mediated sympathoinhibitory effect. This finding is consistent with a previous study showing impairment of the cardiopulmonary mechanoreflex in a canine model of pacing-induced heart failure. 12 The cardiopulmonary baroreflex possibly resets to higher cardiac filling pressure in the MI rats 13 , and the resulting rise in cardiac filling pressure may contribute to the maintenance of cardiac output in heart failure. 27
Effects of vagotomy on peripheral arc characteristics
To the best of our knowledge, this study demonstrated for the first time that vagotomy decreases the slope of the peripheral arc in NC rats (Fig. 3, fourth row). This result was not anticipated because electrical stimulation of either the afferent 10 or the efferent 28 vagal nerve did not significantly change the peripheral arc. Since native vagal activity and electrical vagal stimulation had entirely different discharge patterns and activated fiber components, the intact vagal nerves may modulate SNA in a more complex manner than what is simply deduced from the electrical stimulation experiments.
Several factors can be proposed to explain the decrease in the slope of the peripheral arc after vagotomy in NC rats. First, we recorded the multifiber activity of the splanchnic sympathetic nerve. If vagotomy increases the activities of nerve fibers that do not contribute to AP elevation, the slope of the peripheral arc may be reduced after vagotomy. Second, SNA exhibits regional differences,29,30 and the effects of vagotomy may differ among neural districts. If vagotomy predominantly increases splanchnic SNA but not SNA related to other neural districts, the slope of the peripheral arc may also decrease. Such regional difference in the effect of vagotomy can be found in an earlier study in anesthetized dogs, 5 in which vagotomy significantly increased celiac, mesenteric, and renal artery resistance but not femoral artery resistance. Finally, exposure to a higher maximum AP during the first input sequence after vagotomy could facilitate pressure diuresis, restoring AP toward the pre-vagotomy baseline level. Further studies are required to identify the mechanisms by which vagotomy reduces the slope of the peripheral arc in NC rats.
Effects of vagotomy on HR control
The HR response range was narrower in MI than in NC rats (Table 2, negative bMI for P1), which is consistent with our previous study using rats that survived 100-200 days after MI. 14 The reduced HR response to CSP may be partly explained by the desensitization of beta-adrenergic receptors in the heart in response to sustained sympathetic drive in heart failure. 31 Vagotomy significantly increased minimum HR in the NC group (Fig. 3, second row), reflecting interruption of vagal efferent-induced bradycardic effect. However, vagotomy did not narrow the HR response range, suggesting that vagal nerves contributed little to the expansion of the HR response range. The fact that HR was higher than that observed under conscious conditions (280-300 beats/min) 15 also implies a vagolytic effect of anesthesia in the present study. Notwithstanding the limitation related to the use of anesthesia, vagotomy did not significantly affect the parameters of HR control in MI rats (Fig. 4, second row), confirming withdrawal of the efferent vagal nerve activity in heart failure.
Limitations
As noted above, due to the use of anesthesia, the results may not be directly extrapolated to interpret baroreflex function under conscious conditions. In the present study, direct comparison of absolute levels of SNA between the NC and MI groups was impossible because SNA was normalized in each animal. As a possible solution, AP responses to graded doses of pressor agents may be used to calibrate SNA and standardize the abscissa of the baroreflex equilibrium diagram for both NC and MI groups. 17 A similar strategy may be adopted to obtain a baroreflex equilibrium diagram with putative absolute SNA.
Conclusion
Open-loop static characteristics of the carotid sinus baroreflex were depressed in 8-week post-MI rats. In the NC group, vagotomy caused a sustained increase in SNA, suggesting the presence of a tonic sympathoinhibitory effect before vagotomy. While AP also increased after vagotomy in the NC group, the elevation of AP was transient, contrary to the sustained elevation of SNA. The dissociation of SNA and AP responses after vagotomy in the NC group may explain some of the discrepancies among previous reports regarding the effects of vagotomy on baroreflex-mediated sympathetic AP regulation. The effects of vagotomy on SNA and HR were not obvious in MI rats, confirming withdrawal of both afferent and efferent vagal nerve activities in heart failure. The present results may add evidence supporting the potential use of electrical or pharmacological vagal activation for the treatment of heart failure.15,16,32
Author Contributions
Conceived and designed the experiments: TK, MS. Performed the experiments: TK, ML, CZ. Analyzed the data: TK. Wrote the first draft of the manuscript: TK. Contributed to writing of the manuscript: ML, CZ, MS. Agree with manuscript results and conclusions: TK, ML, CZ, MS. All authors reviewed and approved of the final manuscript.
Footnotes
Acknowledgement
Part of this study was published in a preliminary form in an article in Japanese that appeared in Igakunoayumi. 2012;243:466-74.