
Abstract
Polarity protein complexes are crucial for epithelial apical-basal polarity and directed cell migration. Since alterations of these processes are common in cancer, polarity proteins have been proposed to function as tumor suppressors or oncogenic promoters. Here, we review the current understanding of polarity protein functions in epithelial homeostasis, as well as tumor formation and progression. As most previous studies focused on the function of single polarity proteins in simplified model systems, we used a genomics approach to systematically examine and identify the expression profiles of polarity genes in human cancer. The expression profiles of polarity genes were distinct in different human tissues and classified cancer types. Additionally, polarity expression profiles correlated with disease progression and aggressiveness, as well as with identified cancer types, where specific polarity genes were commonly altered. In the case of Scribble, gene expression analysis indicated its common amplification and upregulation in human cancer, suggesting a tumor promoting function.
Introduction
Most human cancers arise in the epithelial monolayers that line many organs, including the lung, colon, breast, prostate, etc. Normal epithelial cells exhibit apical–basal polarity, where the apical region faces the lumen while the basal surface is attached to the basement membrane. This intrinsic property is established and maintained by the restricted spatial localization and function of the so-called cell polarity proteins.1–4 These proteins also regulate the formation of junctional structures that interconnect neighboring epithelial cells.5,6 Specifically, tight junctions (TJs) localize at the sub-apical region where they separate apical domains from basolateral areas and serve as barriers to prevent molecules diffusing across the epithelial sheets. Adherens junctions (AJs), which are more basal to TJs, connect adjacent epithelial cells to each other and to their underlying actomyosin cytoskeleton. This well-organized architecture allows epithelial cells to function properly to precisely control cellular events in response to cues from neighboring cells and the microenvironment. In contrast, when cellular junctions or cell polarization is perturbed, epithelial cells lose their ability to tightly control their growth and migration, ultimately leading to tumor formation and cancer metastasis.7–10 Therefore, not unexpectedly, polarity proteins have been proposed to function as tumor suppressors or tumor promoters in human cancer, as reviewed recently.11,12 The involvement of these proteins in tumor progression and metastasis has been suggested in a number of studies, from the original genetic studies in Drosophila to more recent studies in mammalian cell lines and mice (see below). Unlike these studies, though, which focus on individual polarity proteins using single gene depletion or overexpression approaches, the recent explosion of cancer genomics data from patient samples provides a unique insight into the role of polarity proteins in different types of human cancer and holds the potential to significantly enhance our view of polarity protein function in tumor progression.
In the case of polarity genes, genomic studies are also important for another reason. As epithelial cells in different tissues perform unique functions, it is expected that the complement of polarity proteins they utilize is also unique. In agreement, a number of closely related paralogs as well as multiple isoforms exist for most polarity proteins, exhibiting restricted tissue distribution and specialized function.13–18 Combined with the well-established crosstalk between polarity complexes, this complexity is better appreciated using a systematic approach to examine these polarity molecules as a whole rather than studying an individual polarity protein out of context.
Here, we first briefly review the current understanding of polarity protein functions in epithelial homeostasis and then focus on their roles in tumor formation and progression. For purposes of clarity and brevity, this work is focused on protein complexes that govern the formation and maintenance of apical-basal polarity. Information for genes related to planar cell polarity in cancer can be found in other articles.19,20 Additionally, we present the expression profiles of polarity complex proteins in different cancers collected from The Cancer Genome Atlas (TCGA, http://cancergenome.nih.gov/) RNASeq database. In summary, mining this great resource suggests that 1) while polarity proteins are fundamental for epithelial function, their roles in different types of cancers are distinct and context-dependent, and 2) the expression of polarity gene profiles differentiates tumor types and correlates with cancer progression. The data suggest that polarity gene profiles can be useful as biomarkers to predict cancer progression, or to develop individualized treatments for cancer therapy.
Polarity Proteins Govern Epithelial Polarization
Establishment and maintenance of epithelial cell polarization is mainly controlled by three polarity complexes, called Crumbs, Partitioning defective (Par), and Scribble (Fig. 1). These complexes, which were originally identified in Drosophila and Caenorhabditis elegans model systems, localize at distinct epithelial membrane domains and function either in a cooperative or antagonistic manner to induce cellular asymmetry and to establish apical-basal polarity.1,21–31 Specifically, the Crumbs and Par complexes localize at the apical regions to specify the apical membrane and determine cell polarity.24–26,32–34 On the other hand, the Scribble complex localizes at the basolateral membrane domain where it excludes apical proteins. 1 The function of polarity proteins in epithelial polarity and homeostasis has been reviewed in detail recently,2–4 and is beyond the scope of this paper.
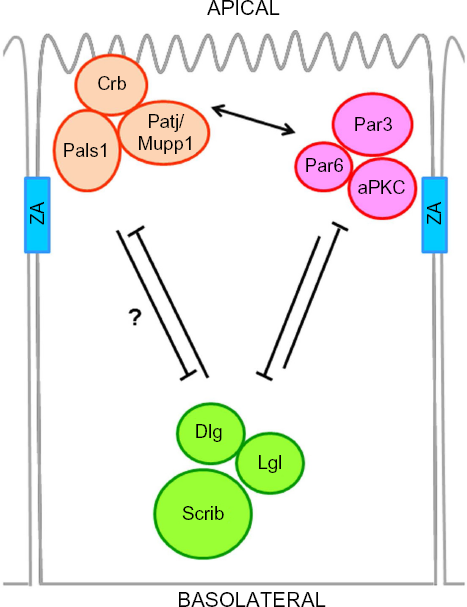
Core members and crosstalk between cell polarity complexes. Epithelial polarity is initiated and maintained mainly by three cell polarity complexes, namely the Crumbs, Par, and Scribble complexes. The Crumbs complex consists of Crb, Pals1, and Patj or its paralog Mupp1; the Par complex consists of Par3, Par6, and aPKC; and the Scribble complex is composed of Scrib, Dlg, and Lgl. A simplified view of the crosstalk between these complexes is shown. The Crumbs and Par complexes localize to the apical region of the cell and work together (arrow) to antagonize the function of the Scribble complex at the basolateral region of the cell.
Polarity Proteins and Cancer
Genetic studies in Drosophila initially established that polarity proteins play important roles in cell growth, asymmetric cell division, invasion, and metastasis.7,35 The data suggested a tumor suppressor function for polarity proteins, which was further supported by the observation that oncogenic viruses often target polarity proteins for degradation.36,37 The implication of these studies and others listed below is that loss of polarity protein function during tumor progression promotes a dysplastic phenotype that either precedes hyper-proliferation, or succeeds it, to induce neoplastic cell growth.1,38
Crumbs Complex and the Hippo Pathway
In mammalian epithelial cells, the Crumbs complex is composed of Crb3, an isoform widely expressed in epithelial tissues,
The role of Crumbs as a tumor suppressor by activating the Hippo signaling was initially proposed by studies using Drosophila tumor models.38,44,45 The Hippo pathway is a crucial, evolutionally conserved cascade that senses extracellular cues to control cell proliferation and organ size. 46 Activation of Hippo signaling results in phosphorylation and inhibition of Yap/Taz (Drosophila Yorkie) transcriptional factors, which activates the expression of genes related to cell proliferation and anti-apoptosis. Similarly, in mammalian cells, Crb3 along with Palsl, Patj/Mupp1, and angiomotins promotes the phosphorylation and cytoplasmic retention of Yap/Taz.47–49 Ultimately, this complex inhibits TGF-β/SMAD signaling and the epithelial–mesenchymal transition (EMT) program. 47 In turn, overexpression of TGF-β, or other EMT regulators such as Snail and ZEB1 prevents expression of Crb3, suggesting a role for Crb3 in inhibiting cancer progression.50,51 Indeed, using an in vivo selection for tumorigenesis method, Karp et al showed that loss of Crb3 contributes to tumor formation. 52 In this system, the tumorigenic progression of immortalized baby mouse kidney cells was accompanied by altered gene expression. Among these genes, Crb3 was found to be dramatically reduced in the acquired tumors, resulting in altered epithelial polarity and morphogenesis as well as tumor expansion. 52 It is important to note here that mammalian cells also express Crb1 and Crb2, two Crumbs members with more restricted expression patterns and important roles in development.17,53 Despite resembling Drosophila Crumbs more closely, their function in cancer initiation or progression is unknown.
The role of Patj or Mupp1 in cancer is largely unknown; both are targeted by viral oncoproteins for degradation or mislocalization, suggesting a tumor suppressor function.36,54–56 However, they also may act as tumor promoters by inducing cancer cell migration. In migrating epithelial cells, Patj was shown to localize at the leading edges, where it mediated directed migration.57,58 Furthermore, Mupp1 cooperated with the RhoGEF Syx to promote the directed migration of oligodendrocyte precursor cells 59 or glioma tumor cells. 60 Pals1 was shown to regulate mTOR and NF–κB signaling to promote cell survival of cerebral cortex and lymphocytes, respectively.61,62 Combined, these studies strongly argue for the potential involvement of Crumbs complex members in cancer.
Par Complex and Growth Factors
The core members of the Par complex are atypical protein kinase C (aPKC), Par3 (a.k.a. ASIP), and Par6, which were initially discovered in a screen for mutants resulting in aberrant asymmetric division in C. elegans.23,63–65 These three members associate with each other to form a ternary complex15,66 although the interaction of Par6 with aPKC is constitutive, and its association with Par3 is mediated and stabilized by the presence of aPKC.67,68 Association of Par3 with aPKC has been reported to inhibit aPKC activity.66,69 Upon activated Cdc42/Rac1, Par6 releases its intrinsic ability to activate aPKC, 70 which then phosphorylates Par3, resulting in the dissociation of Par3 from the Par6/aPKC complex.67,71 The Par6/aPKC complex is then distributed to the apical surface, where apical localization of Par6 is further enhanced by association with Crb3. 72 Activated aPKC also mediates the phosphorylation of lethal giant larvae (Lgl), a component of the Scribble polarity complex, which then translocates to the basolateral membrane of polarized epithelial cells, or the leading edge of migrating cells.30,31 Collectively, it is thought that the Par complex cooperates with the Crumbs complex at the apical domain and antagonizes the basolateral Scribble complex in epithelia to regulate the formation and maintenance of tight junctions and apical-basal polarity.21,22
In mammals, there are two forms of aPKC, aPKC-λ (mouse)/l(human) and aPKC-ζ, which share 70% amino acid identity. 16 Aberrant expression and activity as well as mislocalization of aPKCs are frequently found in cancer. Specifically, expression of aPKCι is upregulated in non-small-cell lung cancer (NSCLC), 73 colon cancer, 74 ovarian cancer, 75 esophageal squamous cell carcinoma (ESCC), 76 breast cancer, 77 and hepatocellular carcinoma. 78 Moreover, increased aPKCι level, which can result from amplification of the 3q26 chromosomal region, is positively correlated with stages of these cancers and poor survival in patients. In NSCLC and colon cancer cells, aPKCι functions as a downstream effector of Ras signaling to promote Rac1 activation and cell transformation.73,74,79 Using a mouse model, aPKCι was shown to promote K-ras-mediated tumorigenesis and induce the expansion and transformation of bronchioalveolar stem cells (BASCs). 80 In ovarian cancer, upregulation of aPKCι was associated with increased level of Cyclin E, a key cell-cycle regulator that is upregulated by aPKC in Drosophila. 75 Additionally, aPKCι is upregulated in basal cell skin cancer and functions as a key activator of hedgehog signaling, which mediates a positive feedback loop resulting in the transcriptional activation of aPKCι and promoting cell proliferation. 81 aPKCι has also been implicated in the trafficking of metalloproteinase MT1-MMP to promote breast cancer metastasis. 82 In glioblastoma, aPKC is involved in EGFR- and TNFα-mediated oncogenic signaling to promote tumor growth, and its expression was inversely correlated with patient survival. 83 Taken together, aPKCι is considered as an oncoprotein in many types of cancers.
Similar to aPKCι, aPKCζ is often altered in cancer. A number of studies suggest that aPKCζ mRNA or protein levels are upregulated in hepatocellular carcinoma, 84 prostate cancer, 85 bladder transition cell carcinoma (TCC), 86 pancreatic ductal cancer, 87 head and neck squamous cell carcinoma (HNSCC), 88 and breast cancer. 89 Expression of aPKCζ correlates with poor survival in patients with non-gastrointestinal stromal tumor soft tissue sarcomas (non-GIST STSs). 90 In HNSCC cells, aPKCζ is required for EGF-mediated MAPK activation and cell proliferation. 88 Moreover, aPKCζ was shown to control front-rear polarity of migrating cells and EGF-mediated chemotaxis in breast cancer cells, suggesting a role in cancer metastasis.91–93 On the other hand, there is also evidence that aPKCζ can function as a tumor suppressor. In colorectal cancer, patients with higher PKCζ level have better prognosis. 94 Nutrient deprivation induces the activation of aPKCζ, which mediates the reprogramming of tumor metabolism to impair tumorigenesis. 94 Furthermore, aPKCζ suppressed the expression of interleukin-6 to prevent cell proliferation and tumor formation in a Ras-induced lung cancer mouse model. 95 Finally, using an antibody against both aPKC isoforms, a recent study suggested that higher aPKC levels correlate with delayed tumor recurrence in patients with superficial bladder cancer. 96
Par3 may also function as a tumor suppressor or pro-tumorigenic protein in different types of cancer. Conditional Par3 knockout in mouse skin delays the formation of Ras-induced papillomas due to reduced cell proliferation and increased apoptosis, suggesting a tumor-promoting role of Par3. 97 However, the same condition predisposes skin to develop another type of cancer termed keratoacanthoma, consistent with a tumor-suppressing function for Par3. 97 In breast cancer, Par3 is downregulated compared to normal tissue, while its membrane localization is also decreased in metastatic cancer cells relative to their primary cancer counterparts.98,99 In addition to morphogenesis, Par3 plays a key role in controlling front–rear polarity, migration, and chemotaxis.100–105 These observations suggest a role for Par3 in regulating critical processes for cancer metastasis. Loss of Par3 in combination with activating Notch or Ras promotes tumorigenesis in mouse mammary glands and induces tumor cell invasion and metastasis.99,106 In these systems, Par3 depletion upregulates Rac1/JNK and aPKC/JAK/Stat3/MMP signaling pathways to circumvent apoptosis, induce cell proliferation, and promote metastasis. 106 Moreover, Par3 knockdown cooperates with ErbB2 activation to promote breast cancer cell invasion and tumor metastasis by aberrantly activating Rac1 activity, reorganizing the cytoskeleton, and suppressing cell-cell adhesion. 98 Additionally, Par3 is downregulated and correlates with cancer metastasis in ESCC tumors, 107 while deletion of Par3 is found in HNSCC and glioblastoma primary tumors. 108 In contrast, both total and cytoplasmic levels of Par3 are upregulated and associated with cancer metastasis and poor survival in human hepatocellular carcinoma 109 and renal clear cell carcinoma (RCC).110,111 Collectively, the data support an important role for Par3 in tumor progression and metastasis, although the specific effects of Par3 depletion or upregulation are context dependent.
In mammals, Par6 proteins are encoded by three genes, namely Par6α (a.k.a. PARD6A, Par-6C), Par6β (a.k.a. PARD6B, Par-6B), and Par6γ (a.k.a. PARD6G, Par-6G).14,15 A number of studies support a pro-tumorigenic role for Par6 proteins in many aspects of breast cancer formation and progression. Both the DNA copy number and mRNA level of Par6β are upregulated in breast cancer and correlate with poor survival.112–115 The Pard6α/aPKC complex was shown to mediate ErbB2-mediated cell transformation and invasion,116,117 while overexpressed Par6α promoted cell proliferation by upregulating ERK signaling following its interaction with aPKC and Cdc42. 112 Furthermore, Pard6a associates with the TGF-β receptor and is required for downstream cytoskeletal reorganization and thereby EMT. 118 Indeed, Par6's presence was highly associated with expression of EMT characteristics in human basal type breast cancer, and interfering with TGF-β/Par6 signaling blocked metastasis of orthotopic mouse mammary tumors. 119 Similarly, Par6 plays a key role in Rasmediated NSCLC tumor progression. In this context, the interaction of Par6 with aPKCι is required for activation of Rac1 and induction of cell transformation and invasion.120,121 Moreover, the TGF-β/Par6 pathway is also important for EMT induction in NSCLC cells. 122 Interestingly, the function of Par6 may not be limited to cancer cells but also affect the tumor microenvironment, as high Par6 expression in stromal cells correlates with better prognosis in NSCLC. 123
The Scrib Complex and Oncogenes
The Scrib complex includes three members; Scribble (Scrib, a.k.a. Vartul), lethal giant larvae (Lgl, a.k.a. HUGL, and LLGL), and disk large (Dlg). Although they belong to the same genetic pathway, 1 and each member has been shown to associate with another, the actual interaction of all three members within a complex remains unclear.124–126 In Drosophila, Scribble plays a crucial role in maintaining apical-basal polarity; however, it has stronger influence on planar cell polarity in mammals.127,128 One possibility is that other members of the leucine-rich repeats and PDZ (LAP) protein family, which comprises Scrib, Erbin, Densin-180, and Lano in mammals, may also play a role in apical-basal polarity. 129 Lgl, on the other hand, whose localization and function is regulated by aPKC-mediated phosphorylation, regulates epithelial polarity in both systems.29,31 Additionally, Dlg appears to play an important role in both apical-basal polarity and planar cell polarity of epithelial cells.130,131 Scribble binds the planar cell polarity proteins Vangl1 and Vangl2, which are required for proper Scribble localization and may also regulate apical-basal polarity via Scribble.125,132,133 The role of Vangl proteins in planar cell polarity and cancer has been reviewed recently and is beyond the scope of this paper. 134
Initial studies indicated that Scribble complex members play a key role in inhibiting the uncontrolled growth of Drosophila imaginal disks during development, supporting a tumor suppressor function.1,135,136 Furthermore, the polarity defects and tumor outgrowth of Scribble mutant flies were rescued by ectopic expression of the human Scribble homolog. 137 In general, later studies have supported a tumor suppressor role for Scribble in human cancer. Early studies suggested that Scribble is downregulated and mislocalized in breast cancer,138,139 although a more recent study argues that Scribble is often upregulated and mislocalized. 140 Loss of Scribble cooperates with myc oncogenes to block apoptosis and to promote cell transformation and breast cancer formation. 139 Additionally, Scribble downregulation or mislocalization induced by EMT or loss of LKB1/MARK signaling inhibited the Hippo pathway and activated Taz/Yap-mediated tumorigenesis.141,142 In prostate and lung carcinomas, loss of Scribble cooperates with activating Ras mutations to promote tumorigenesis and tumor progression.143,144 Additionally, knockdown of Scribble in combination with activation of different growth factors, cytokines, or Ras signaling promotes the migration and invasion of cancer cells.117,145 In agreement, reduced Scribble levels correlate with invasiveness in cervical cancer. 146 Conversely, overexpression of Scribble has been shown to inhibit expression of the EMT regulator ZEB-1, promoting the expression of tight junction proteins, such as Crb3. 147 In contrast to these studies, there are also reports that suggest a role for Scribble in tumor progression. Scribble has been shown to play a key positive role in the establishment of front-rear polarity and directed cell migration by controlling the localization and activity of Cdc42 and Rac1 at the leading edge of migrating cells and regulating the downstream activation of p21-activated kinases (PAKs).148–150 In the case of breast cancer, increased expression of cytoplasmic Scribble resulted in activation of the Akt/mTOR/S6K pathway to promote mammary carcinogenesis. 140 Finally, in agreement with the hypothesis that Scribble does not act as a classic tumor suppressor, its locus is amplified in many human cancers (http://www.cbioportal.org/public-portal/).151,152
Unlike Scribble, which is thought to be functionally distinct from its family members, Dlg and Lgl are expressed by several related genes in mammals, complicating their roles in cancer. The Dlg family comprises of five members in humans: Dlg1 (a.k.a. SAP97), Dlg2 (a.k.a. PSD95, Chapsyn-100), Dlg3 (a.k.a. SAP102, NE-Dlg), Dlg4 (a.k.a. SAP90, PSD95), and Dlg5 (a.k.a. lp-Dlg, P-Dlg).153–156 Among them, expression of either Dlg1 or Dlg3 is able to suppress tumor growth in Drosophila Dlg mutants
157
; importantly, several human Dlg paralogs are targeted by human virus oncoproteins for degradation.36,158–164 Dlg1 is the most studied member in cancer and has been shown to directly interact with the tumor suppressors
Consistent with a tumor suppressor function, altered localization and overall expression of Dlg is shown in cervical carcinomas, which are often related to human papillomavirus (HPV) infection.177–179 In colorectal cancers that are related to APC mutation/inactivation, the expression and distribution of Dlg is also perturbed. 180 Moreover, somatic mutations in Dlg1 have been reported in breast ductal carcinoma, 181 and Dlg3 is downregulated in gastric, 182 esophageal,174,175 and papillary thyroid carcinoma, 183 as well as glioblastoma. 176 In contrast, Dlg3 is reportedly upregulated in serous ovarian carcinomas. 184
A role of Dlg5 in cancer was suggested by the discovery that its genetic variant R30Q is associated with inflammatory bowel disease and Crohn's disease, which predispose patients to colorectal cancer.185,186 By interacting with and promoting the degradation of TGF–β receptors, Dlg5 prevents the occurrence of EMT, which, in turn, induces the downregulation of Dlg5.187–189 Loss of Dlg5 results in increased cell migration and invasion through the activation of Akt or ERK/p90 ribosomal S6 kinase signaling pathways.189,190 Finally, while loss of Dlg5 is reportedly frequent in prostate cancer, 190 Dlg5 also promotes the growth of pancreatic cells and is upregulated in diffuse gastric and pancreatic cancers.191,192
Lgl is the third member of the Scribble complex and comprises of two Drosophila orthologs in mammals (Lgl1 and Lgl2).31,193 Deregulation of Lgl's function leads to cancer formation and progression as shown by the finding of neuroblastoma formation in Drosophila lgl mutants 135 and brain dysplasia and invasion in Lgl1 knockout mice. 194 In human, loss of Lgl1 copy number correlates with poor survival of glioblastoma patients. 195 In addition, Lgl1 is frequently lost or downregulated in prostate, breast, and colon cancers, as well as malignant melanoma.196–198 Further examination revealed a role of Lgl1 in decreasing cell migration, invasion, and EMT, and showed that loss of Lgl1 correlates with advanced stages of malignant melanoma and colorectal cancer.197,198 Similarly, Lgl2 mutation in zebrafish (penner mutant) results in epidermal hyperproliferation, as well as ErbB2-mediated EMT activation.199,200 Studies in human cells demonstrated that the EMT regulators ZEB1 and Snail repress the expression of Lgl2, whose expression prevents the nuclear translocation and thus function of Snail.51,201 Finally, deregulation of Lgl2 by decreased expression or mislocalization was reported in gastric dysplasia and adenocarcinoma,202,203 and reduced Lgl2 correlated with aggressiveness in colorectal cancer. 204
Overall, the data argue that the Scribble complex acts as a tumor suppressor in most cases, and that reduced expression or mislocalization of Scribble complex components is common in cancer.
Differential Expression of Polarity Proteins in Cancer
As expected from their crucial role in regulating cell polarity, directed migration, asymmetric cell division, and growth, all polarity complex proteins are involved in cancer progression. However, the functional diversity of polarity proteins, both with the existence of multiple paralogs in the human genome and with the expression of multiple isoforms per gene, suggests that different tissues utilize a distinct set of polarity proteins to generate and maintain their differentiated state. Early studies in Drosophila have indicated that polarity proteins play a tumor-suppressing role. However, our current understanding of polarity signaling underscores the presence of polarity proteins in functionally distinct and mutually exclusive polarity complexes that specify membrane subdomains to induce cell polarization. In this scenario, the loss, overexpression, or mislocalization of polarity proteins in cancer cells could all result in impaired polarity signaling and promotion of tumorigenesis.
To start addressing the relative contributions of polarity genes in human cancer, we focused our analysis on the core members of the major complexes that regulate epithelial apical-basal polarity. Since the TCGA now provides the genomic profiling data of thousands of samples representing all major cancer types, we assessed the expression of core polarity components in these samples and generated their profiles. A detailed description of samples used and data analysis including statistical calculations are provided in the Methods section.
The Gene Expression Profiles of Polarity Proteins Differentiate Cancer Types
The expression levels of individual polarity genes from the TCGA tumor RNA sequencing data were compared among 5416 patient samples representing 15 human cancer types (Fig. 2). The polarity genes were analyzed using unsupervised hierarchical clustering analyses. Analysis was restricted to genes related to the three main polarity complexes described earlier, including their known paralogs and functional isoforms. Unexpectedly, clustering resulted in the separation of different cancer types (Fig. 3, Supplementary Table 1). In other words, the expression profiles of the polarity genes classified cancer types, which may be beneficial for distinguishing metastatic cancer versus a new primary cancer of different tissue origin. The data support the hypothesis that different polarity proteins are involved in the progression of different cancer types, and provide a first glimpse into the subset of polarity proteins that may be deregulated in each cancer type.

The number of samples from different cancer types downloaded from the TCGA and used in this study.

Expression profiles of polarity complex genes in human cancer. The heat map shows differential expression of polarity genes in 15 different cancers. mRNA level relative to the mean of zero is depicted by the blue-red intensities (red = 3.0 high; white = 0.0, mean; blue = −3.0, low). Integrative and hierarchical clusterings were detected using the Patrek genomic suite. Individual polarity genes and cancer types are listed on the x-, and y-axis, respectively.
Alteration of Polarity Proteins in Invasive Breast Cancer
While the above analysis compares polarity profiles across cancer types, it provides no information on the relative expression changes of polarity genes in tumor versus normal tissues. This type of analysis is critical for understanding the relative contribution of individual polarity genes in tumor progression; however, only limited numbers of normal samples are currently available for most cancer types. We chose breast cancer to perform this analysis because breast cancer is one of the top five leading causes of cancer death worldwide, and a large number of both cancer and normal samples have been collected and sequenced.
Using the same unsupervised clustering method to analyze both the normal and breast cancer data as above, we have clustered polarity genes according to their overall expression pattern (Fig. 4A, Supplementary Table 2). Interestingly, the expression profiles of the 20 polarity genes automatically segregated normal from tumor breast tissue samples. Similar results were obtained when analyzing other cancer types, showing the power of this analysis for differentiating normal from tumor samples and highlighting the significance of polarity genes in tumor formation (Supplementary Fig. 1). In breast cancer, normal tissue samples and a very small number of tumors clustered closely together, whereas breast cancer samples were classified into two major clusters. This finding is consistent with the presence of genomic heterogeneity in breast cancer, and further analysis will be required to assess whether this segregation is dependent on the subtype (luminal vs basal) or aggressiveness of each tumor sample. Our analysis revealed significant expression changes in polarity genes in breast cancer. Although SCRIB (Scribble) was initially thought to be downregulated in breast cancer,138,139 a recent study argued its upregulation, 140 which is consistent with our data. As mentioned earlier, several members of polarity complex proteins are expressed as multiple paralogs, often referred to as isoforms. In most cases, we know very little about the differences in the expression and function of different isoforms. Our bioinformatics analysis suggests that polarity protein isoforms exhibit differential expression patterns in breast cancer, likely reflecting isoform-specific functions. For example, while Dlg3 is upregulated in breast cancer samples, DIg2 is downregulated (Fig. 4A). Similarly, there is an upregulation of PARD6A (Par6α) and PARD6B (Par6β), but a decrease in the levels of PARD6G (Par6γ). Crb3 is upregulated in our breast cancer samples, while its paralogs Crb1 and Crb2 are downregulated (Fig. 4A, supplementary Table 2). Crb1 and Crb2 are large proteins with extended extracellular regions like Drosophila Crumbs, while Crb3 is a relatively small molecule (13 kDa) with a very short extracellular domain. 3 Despite the discrepancy, Crb isoforms possess similar cytoplasmic domains that mediate their interaction with Pals1, Patj, and Muppl.18,205,206 Moreover, in polarized epithelial cells, both Crb1 and Crb3 localize at tight junctions,18,205 although little is known about the role of Crb1 in epithelial polarity as early studies suggested lack of expression in epithelial tissues.207,208 Similar to Crb isoforms, differential expression patterns were also found for aPKC isoforms and Patj/Mupp1 paralogs (Fig. 4A, Supplementary Table 2).
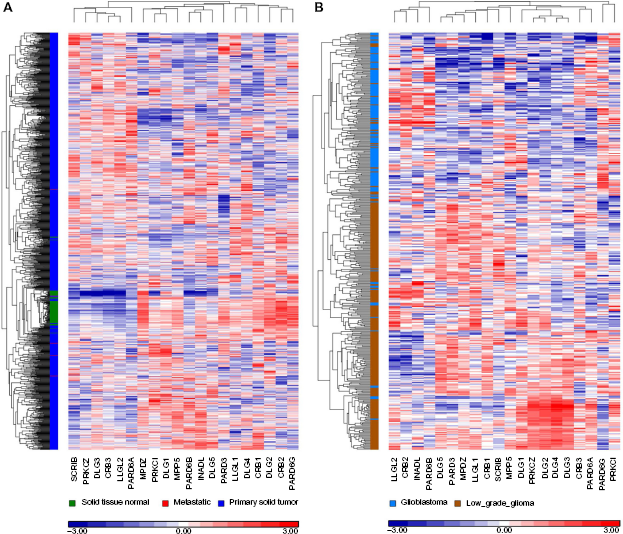
Expression of polarity complex genes in breast (
Alteration of Polarity Proteins and Cancer Aggressiveness
In addition to cancer-specific expression differences, we wanted to examine whether the expression profile of polarity proteins is altered with tumor stage and progression to aggressive disease. To test this, we investigated the expression profiles of polarity proteins in two sets of brain gliomas, namely low-grade glioma and glioblastoma multiforme (GBM), which are two related cancers with significant differences in aggressiveness and patient outcomes. Malignant glioma is the major type of brain cancer. Based on their degree of malignancy, the World Health Organization (WHO) classifies malignant gliomas from grade II to grade IV, in which grades II and III are referred to as low-grade gliomas, while grade IV is referred to as glioblastoma multiforme (GBM), the most aggressive brain tumor with a median survival of only 15 months.209,210 Therefore, we graphed the expression profiles of polarity proteins in TCGA tumor samples classified as low-grade gliomas and GBM. Based on these expression profiles, our bioinformatics analysis automatically segregated GBM tumors from low-grade gliomas (Fig. 4B, Supplementary Table 3). Interestingly, while most glioma tumors segregated with their respective tumor grades, a few did not, and showed a polarity gene profile characteristic of either GBM or low grade, despite being classified as low-grade or GBM, respectively. One interesting possibility is that these cases follow the overall survival and tumor aggressiveness of the group they segregate with, which could be tested by further bioinformatics analysis on the basis of overall survival. Finally, the low-grade glioma group exhibits significant heterogeneity of polarity protein expression with up to four distinct polarity profiles, which may reflect multiple subclasses of low-grade glioma. Therefore, while we know very little about the role of polarity proteins in brain cancer, bioinformatics approaches can uncover significant alterations in the expression of key polarity genes that are crucial for the progression of human gliomas.
Changes in Scrib Expression in Human Cancer
The power of bioinformatics can also be harnessed to assess the overall expression of a particular gene across multiple tumor types compared to normal tissues. For this analysis, we tested expression of Scrib (Scribble), as the overwhelming majority of cell line and animal data argue for a tumor suppressor function. Nonetheless, the analysis of Scrib DNA copy number indicates that it is amplified in most human cancers (Fig. 5A). Furthermore, our bioinformatics analysis shows that Scribble mRNA is upregulated in several cancers, including breast, lung adenocarcinoma, and hepatocellular carcinoma compared to normal tissues (Fig. 5B). While Figure 5A argues for a significant amplification of Scrib (Scribble) copy number in ovarian cancer, we were not able to confirm this in our analysis due to the lack of control samples. However, the Scrib gene is located on chromosome 8q.24.3, which is a common amplicon in ovarian cancer, supporting the prediction of Scrib upregulation in ovarian cancer. 152 Importantly, Scribble mRNA is not significantly altered in HNSCC, suggesting that Scribble regulation is different in different types of cancer. It is important to note here that copy number changes may not necessarily result in changes in mRNA levels, and changes in mRNA levels may not reflect the relative abundance of protein. Nonetheless, the data suggest that the most common alteration in Scribble expression in human cancer is amplification and increased expression, which will have to be further substantiated in follow-up studies.

Expression of Scrib in different human cancers. (
Conclusions
Epithelial polarity is established and maintained by core polarity proteins that interconnect with each other to specify membrane subdomains in order to regulate cellular function and to maintain tissue homeostasis. Increasing evidence suggests that loss of cell polarity results in tumor formation and cancer progression. Polarity signaling is complex, and the presence of multiple mammalian paralogs and isoforms of key polarity components further complicates the picture. Progress to date has been made largely by studying the function of single polarity genes in cell lines or animal models. However, little is known about the relative expression of polarity genes in cancer, their relative expression in each cancer type, and their involvement in the progression of particular cancer types. Here, we reviewed the current understanding of polarity protein function in tumor formation and cancer progression, and also provided the first comprehensive analysis of polarity gene expression profiles in human cancer. We used publicly available cancer genomics resources such as TCGA and CBioPortal and examined the expression of core polarity genes in multiple cancer types. The data uncover unique polarity gene profiles in different tissues and cancers, which can be used to classify cancer types. Polarity expression profiles reveal cancer-specific alterations in polarity gene expression, which correlate with aggressiveness and may impact cancer progression. In contrast to previous studies, the characterization of genomic alterations in particular cancer types provides a more comprehensive view of polarity function or dysfunction in human cancer, points to select gene alterations that may “drive” tumor progression, and may be useful as predictive or prognosis biomarkers of disease aggressiveness.
Methods
The TCGA level-3 RNA sequencing data of 15 cancer types were downloaded (Fig. 2). The raw gene counts were normalized to log2 of the read-per-million (log2 RPM). For data clustering, unsupervised hierarchical clustering of both samples and genes (Figs. 1 and 2, rows and columns, respectively) were performed using Pearson's dissimilarity metric for cluster agglomeration and average linkage for calculating distances between clusters. The rows and columns were standardized to mean of zero and scaled to standard deviation of one for the heat maps. Statistical analysis was performed using analysis of variance (ANOVA) for detection of differentially expressed genes between sample groups. The ANOVA P-values were adjusted using both Bonferroni and step-up correction methods provided in the Partek Genomic Suite software. The dot plots of individual genes were drawn using the Partek Genomic Suite.
Author contributions
Data analysis: YWA, WL. Manuscript preparation: WL, YWA, PAZ. All authors reviewed and approved of the final manuscript.
Supplementary Materials
Supplementary Figure 1
Expression of polarity complex genes in different cancers. Integrative clustering of the expression of polarity genes in different types of cancers is shown. Normal and tumor tissues from head and neck squamous cell carcinoma, hepatocellular carcinoma, lung adenocarcinoma, lung squamous carcinoma, renal clear cell carcinoma, prostate adenocarcinoma, and thyroid carcinoma are indicated. Relative mRNA level is depicted by the blue-red intensities (red = 3.0 high; white = 0.0, average; blue = −3.0, low).
Supplementary Table 1
Expression values of polarity complex genes in different cancers. The binary logarithm (log2) of the average expression values is shown. Relative expression levels of individual genes among different cancer types is shown by the blue-red intensities. Data is organized as the layout of the Figure 3 (
Supplementary Table 2
Expression value of polarity complex genes in breast cancer. The binary logarithm (log2) of the average expression values (n) is shown. Expression levels of individual genes from primary tumor samples are compared with those from normal solid tissues. Statistical analysis was performed using ANOVA alone or with Bonferroni and step-up corrections. P-Value is shown. Nonsignificant difference is highlighted in gray.
Supplementary Table 3
Expression value of polarity complex genes in glioma. The binary logarithm (log2) of the average expression values (n) is shown. Expression levels of individual genes expressed in glioblastoma multiforme (GBM) are compared to those from low-grade glioma. Statistical analysis was performed using ANOVA alone or with Bonferroni and step-up corrections. P-Value is shown. Nonsignificant difference is highlighted in gray.