
Abstract
Curcumin, a phytochemical isolated from curcuma plants which are used as coloring ingredient for the preparation of curry powder, has several activities which suggest that it might be an interesting drug for the treatment or prevention of cancer. Curcumin targets different pathways which are involved in the malignant phenotype of tumor cells, including the nuclear factor kappa B (NFKB) pathway. This pathway is deregulated in multiple tumor entities, including Hodgkin's lymphoma (HL). Indeed, curcumin can inhibit growth of HL cell lines and increases the sensitivity of these cells for cisplatin. In this review we summarize curcumin activities with special focus on possible activities against HL cells.
Introduction
The name curcumin has been used for different unrelated dyestuffs. Today, the name is used more or less exclusively for 1,7-bis(4-hydroxy-3-methoxyphenyl)-1,6-heptadiene-3,5-dione, the yellow dye from Curcuma longa (turmeric) and other curcuma plants.1,2 The name curcuma is based on the oriental name of the plant, which refers to its similarity with saffron (see the Hebrew word for saffron: 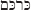 ).
3
Approximately 2000 years ago, Plinius described a plant with similar features which is considered to be turmeric (est et per se Indica herba, quae cypira vocatur, zingiberis effigie; commanducata croci vim reddit.
4
[furthermore, an Indian plant exists which is called cypira and which has a similar appearance as ginger; while chewing it constitutes the effect of saffron]). Hybridization of C. longa with other species results in marked differences in the curcumin content between different plants.
5
C. longa is a member of the family Zingiberaceae (ginger family). This family contains several plants which are widely used as spices and/or officinal plants, for example Aframomum spec. (alligator pepper, false cardamom), Alpinia officinarum (Chinese ginger), Amomum subulatum (hill cardamom), Boesenbergia rotunda (Chinese keys), C. aromatica (wild turmeric), C. xanthorrhiza (temulawak), C. zedoaria (zedoary), Eletteria cardamomum (cardamom), Kaempferia galangal (galangal), Zingiber officinale (ginger), and Z. mioga (Japanese ginger).
).
3
Approximately 2000 years ago, Plinius described a plant with similar features which is considered to be turmeric (est et per se Indica herba, quae cypira vocatur, zingiberis effigie; commanducata croci vim reddit.
4
[furthermore, an Indian plant exists which is called cypira and which has a similar appearance as ginger; while chewing it constitutes the effect of saffron]). Hybridization of C. longa with other species results in marked differences in the curcumin content between different plants.
5
C. longa is a member of the family Zingiberaceae (ginger family). This family contains several plants which are widely used as spices and/or officinal plants, for example Aframomum spec. (alligator pepper, false cardamom), Alpinia officinarum (Chinese ginger), Amomum subulatum (hill cardamom), Boesenbergia rotunda (Chinese keys), C. aromatica (wild turmeric), C. xanthorrhiza (temulawak), C. zedoaria (zedoary), Eletteria cardamomum (cardamom), Kaempferia galangal (galangal), Zingiber officinale (ginger), and Z. mioga (Japanese ginger).
Recently, the complete transcriptome of C. longa rhizomes was analyzed.
6
For a long time, the lipidsoluble yellow compound which can be isolated from the curcuma plants was used for cooking, cosmetics, and textile dying.
7
In the 13th century, turmeric was introduced into Europe by Arab traders.
8
Today, turmeric and the isolated curcumin are used as colorant in many food products. Turmeric is one of the important ingredients of curry powder. The color of curcumin depends on the pH and the presence or absence of other substances like boron (Fig. 1). With boron, curcumin forms rosocyanine, a reddish color (Fig. 1) which can be used for the detection and colorimetric quantification of boron.
9
curcumin and curcuma. A) Curcumin is the yellow dyestuff from turmeric (the structure of curcumin was drawn with C-Design 3.0f; http://www.ch.tum.de/oc1/EFontain/C-Design/). B) Curcumin forms colored complexes with boron. Turmeric powder (1 g) was extracted with 96% ethanol (10 mL, 37oC, 2 h), centrifuged and steril filtered. In the presence of 0.1 M NaOH, a 1:20 dilution of this extract in water is colored orange. After adition of 0.05 M boric acid the colore turns into deep red. C) Turmeric powder (1.3 g) was extracted with 96% ethanol (18 mL, 37oC, 4 h) in the absence or presence of boric acid (1 g), centrifuged, steril filtered, and dried on filter paper circles. In the presence of boron, the yellow color changed into red.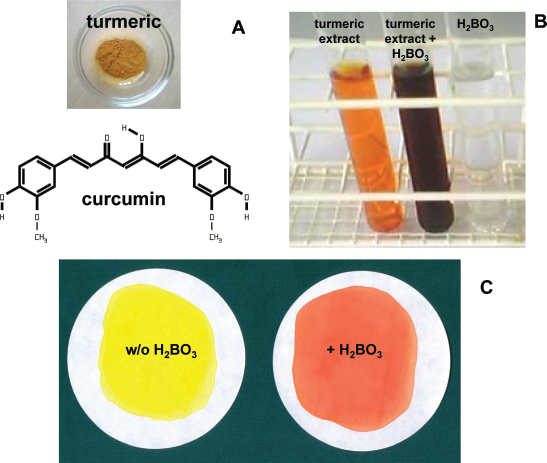
Multiple biological effects have been documented for curcumin. Curcumin has direct antibacterial 10 and anti-inflammatory 11 activity. The anti-inflammatory activity might be mediated in part by the strong antioxidant activity of its caffeic acid moiety. 12 Curcumin influences multiple signaling pathways.13,14 In addition to the anti-cancer related activities described below, curcumin is used for the treatment of a plethora of other diseases including pulmonary, neurological, liver, metabolic, and autoimmune diseases. 15
Anti-Cancer Related Activities of Curcumin
In the last decades, there has been growing interest in curcumin and curcumin derivatives for treatment or prevention of cancer as indicated by the increasing number of curcumin-related publications in the last years (Fig. 2). The basic biological activities of curcumin has been summarized in an excellent review.
16
Increasing number of publications about curcumin and cancer. The PubMed database (http://www.ncbi.nlm.nih.gov/pubmed/) was searched with the queries “curcumin” or “curcumin cancer”. Presented are the absolute numbers of found publications per puplication year.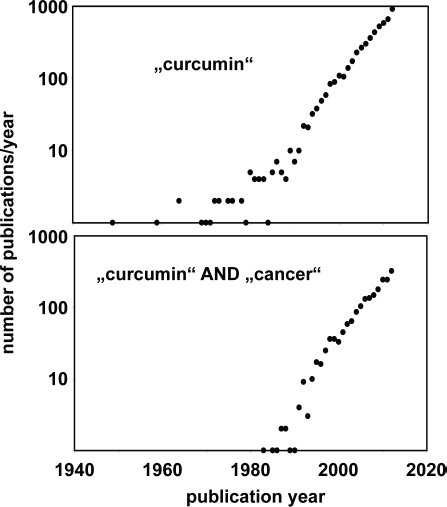
Early observations suggested that curcumin can inhibit growth of tumor cells, 17 can inhibit chemical carcinogenesis,18,19 and can be used for the treatment of patients with cancer. 20 Since then, few clinical trials suggest that curcumin might be a drug of interest for treatment of cancer21,22 or for the prevention of different forms of cancer.23,24
Examples for growth inhibitory activities of curcumin and curcumin derivatives on tumor cells.

Another interesting activity of curcumin is the inhibition of tumor cell motility, invasion, and metastasis.26,42,92–100 This activity is mediated in part by the down-regulation of matrix-metalloproteinases. A key step in tumor metastasis is the process of epithelial mesenchymal transition (EMT) usually accompanied by loss of epithelial cadherin (CDH1) expression. Inhibition of EMT and up-regulation of CDH1 in tumor cells by curcumin has been described.101,102 One inductor of cell motility in cancer cells is lysophosphatidic acid (LPA). LPA receptors are often expressed in tumor cells and co-expression of LPA receptors and LPA-producing phospholipases in tumor cells have been observed. 103 Curcumin can also inhibit LPA-induced cancer cell motility. 101 Taken together, these observations indicate that curcumin does not only inhibit tumor growth but also metastasis formation. However, enhanced metastasis has been described in a Lewis lung carcinoma model. 104
In addition to the death inducing activities of curcumin on established tumor cells, curcurmin and curcumin derivatives can inhibit or delay tumor formation in different tumor models. 92 105–113 Curcumin significantly inhibited the formation of preneoplastic lesions in animal models for chemical colon carcinogenesis.92,102 The combination with fluorouracil or carnitine enhanced this effect. Curcumin inhibited ultraviolet radiation-induced or chemically induced skin carcinogenesis in mice. 106 107 Furthermore, curcumin inhibited the development of gastrointestinal tumors,108,109 mammary tumors, 110 and hepatocellular tumors111,112 in models for chemical carcinogenesis. The anti-inflammatory activity of curcumin seems to be responsible at least partially for such cancer preventive activities. 113 In addition to the direct activities on tumor cells and pre-malignant cells, the inhibition of the growth of carcinogenic microorganisms such as Helicobacter pylori may partially account for tumor prevention by curcumin. 114 Very infrequently, enhanced tumor formation in experimental models has been described. 115 Such observations indicate that the biology of the used experimental models has not been fully elucidated and that further investigations are required in order to understand the effects of curcumin in the different models.
The Nuclear Factor Kappa B (NFKB) Pathway as Target for Curcumin
Avian reticuloendotheliosis defines a group of diseases that are observed in turkeys, broiler chickens, ducks, among other birds. One of the interesting features of reticuloendotheliosis is the development of lymphoid malignancies. The causative agent of reticuloendotheliosis is a gamma-retrovirus that transforms diverse avian cells. Analysis of the T strain of this virus (reticuloendotheliosis virus strain T; REV-T) led to the identification of the REL oncogene.116–118
Thereafter, the human REL homologue was found on chromosome 2.
119
REL is a member of a gene family that includes in the human genome at least 5 members: REL, RELA,120,121 RELB,
122
nuclear factor of kappa light polypeptide gene enhancer in B-cells 1 (NFKB1),
123
and NFKB2 (Fig. 3).
124
This family can be divided into two sub-families (Fig. 3) and usually one member of the NFKB1/2 subfamily forms a heterodimer with a member of the REL subfamily. The NFKB pathway is a highly conserved signaling pathway in eukaryotes. Members of the NFKB family are involved in chromosomal rearrangements that have been detected in cancer cells and gene fusions involving REL125,126 or NFKB2124,127 have been described.
Known human proteins of the REL/NFKB family. Reference sequences of the indicated human proteins were used for a phylogenetic tree analysis using the clustalW algorithm (http://www.genome.jp/tools/clustalw/). For human RELA (protein accession numbers NP_001138610, NP_001230913, NP_001230914, and NP_068810), human NFKB1 (NP_001158884, NP_003989) and human NFKB2 (NP_001070962, NP_001248332) different protein isoforms with highly similar sequences have been identified. For REL (NP_002899) and RELB (NP_006500) no additional human isoforms are in the databases. The family can be separated into two sub-families. A complete NFKB transcription factor consists of a heterodimer of a member of the NFKB1/2 subfamily and a member of the REL subfamily.
The NFKB pathway is often deregulated in cancer cells. One example is Hodgkin's lymphoma (HL), one of the most frequent lymphomas in Western countries. 128 The etiology of HL is unclear, but immunological and molecular properties suggest that in most cases HL cells are derived from B cells.129–131 HL cells have a characteristic gene expression profile that discriminates these cells from other normal and transformed hematopoietic cells. 132 With the combination of radio- and chemotherapy the majority of patients, with HL can be cured. However, the established therapy is associated with the induction of secondary malignancies, cardiac toxicities, and treatment-related infertility.133–135 In addition, up to 10% of patients still cannot be cured with current therapy regimes, which represent a significant number.136,137 Thus, there is a clear demand to search for new treatment options and also for optimization of current treatment strategies by identifying potential treatment resistance mechanisms.
An involvement of the NFKB pathway in HL has been suggested by the high expression of NFKB family members in HL. 138 Mutations in the NFKB inhibitors NFKB inhibitor alpha (NFKBIA) and tumor necros factor alpha-induced protein 3 (TNFAIP3) have been observed in a high percentage of HL.139,140 Curcumin inhibits NFKB activity in different cell types (Table 1) and the inhibition of the NFKB pathway can be seen by decreased expression of NFKB. The activity of NFKB is regulated by inhibitors that bind NFKB and release NFKB only after degradation. Curcumin can inhibit NFKB activity indirectly by stabilization of NFKBIA. 40
In addition to constitutive activation of NFKB, in some tumors NFKB expression and activation is induced by treatment with cytotoxic drugs or cytokines.40,141,142 Curcumin and curcumin derivatives can prevent such induced activation. Curcumin derivatives, for example, have been shown to decrease the TFR-induced expression of NFKB in melanoma and neuroblastoma cells. 40 In some cases the anti-NFKB activity of curcumin requires the presence of additional treatment elements. In pancreatic tumor cells, for example, curcumin inhibits DNA binding-activity of NFKB only in combination with aspirin and sufuraphane. 54 As the final consequence of NFKB inhibition, expression of NFKB target genes is suppressed. These target genes include anti-apoptotic factors and the suppression of these factors can increase tumor cell death. In addition, the crosstalk between NFKB and other signaling pathways can lead to complex alterations of the phenotype of tumor cells after inhibition of NFKB.
The activation of the NFKB pathway in HL makes curcumin an interesting drug for the treatment of HL. Indeed, curcumin induces cell death in HL cells. As shown in Figure 4A, incubation with curcumin leads to decreased cell viability in all HL cell lines tested whereas normal peripheral blood mononuclear cells (PBMC) are not affected by the used concentration of curcumin. Similar results were found by Mackenzie et al who showed that curcumin leads to cell cycle arrest in G2-M and reduced cell viability in Hodgkin and Reed-Sternberg cells.
143
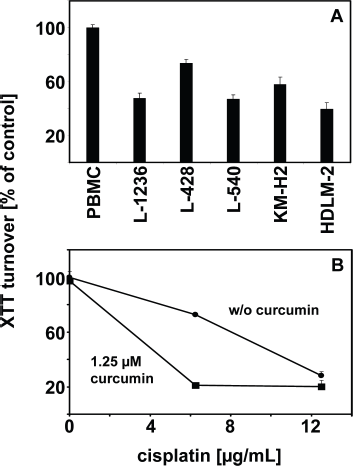
In order to determine whether the combination of curcumin with conventional chemotherapy might have synergistic effects, HL cells were incubated either without curcumin or with a low dose (1.25 μM) of curcumin and varying concentration of cisplatin. The results are shown in Figure 4B. With the used concentration of curcumin, only marginal effects on cell viability were observed. In contrast, the combination of curcumin with cisplatin had a strong cytotoxic effect on the cells. However, inhibition of the Fanconi anemia (FA) pathway might be an important factor for the synergistic effect of cisplatin and curcumin on HL cells. Ubiquitination of the FA complementation group D2 (FANCD2) protein in response to DNA damage is an important step during repair of cross-links in the DNA, caused by cisplatin. Chirnomas et al 144 showed that curcumin inhibits the FA pathway by inhibition of ubiquituation of FANCD2. This inhibition sensitized ovarian and breast tumor cells for cisplatin. 144 It remains to be shown whether similar mechanisms are responsible for increased cisplatin sensitivity of curcumin treated HL cells. The HL cell lines used in our study outlined above were established from patients with refractory disease and are not perfect models for the situation in most patients in vivo. However, the increased cell death of these highly chemotherapy resistant cell lines after treatment with curcurmin seems to be encouraging.
Curcumin shares the NFKB-inhibitory activity with another interesting dietary phytochemical: capsaicin. 145 Curcumin can bind the vanilloid receptor TRPV1 (transient receptor potential cation channel, subfamily V, member 1), the same receptor that is activated by capsaicin. 145 However, curcumin binding to TRPV1 did not result in the same signaling. TRPV1 is not the only receptor for curcumin and interestingly, curcumin can bind the aryl hydrocarbon receptor AHR. 146 Many carcinogenic polycyclic aromatic hydrocarbons are carcinogens only after in vivo activation. Cytochrome P450 family member A1 (CYP1A1) is an enzyme which can catalyze this activation. Expression of CYP1A1 is induced after binding of AHR to polycyclic aromatic hydrocarbons. It was shown that curcumin can inhibit the CYP4A1 inducing activity of the AHR. 147 Binding to AHR and competition with aryl hydrocarbon derivatives might explain the chemopreventive activity of curcumin. 147 Probably more important for the cancer cell death inducing activity of curcumin is the third curcumin receptor, the vitamin D receptor.148,149
Vitamin D Receptors (VDR) and Curcumin
Anti-neoplastic activities of vitamin D have been described, but epidemiological data also suggests that for some cancer types sunlight can reduce cancer risk independent from vitamin D. 150 Curcumin increases apoptosis and differentiation of vitamin D-treated tumor cells.151,152 Direct binding of curcumin to the vitamin D receptor (VDR) was demonstrated. 148 This binding allows VDRs to heterodimerize with the retinoic X receptor and translocate to the nucleus. The complex can then activate gene transcription of vitamin D target genes. 148
Recently, high expression of the VDR has been observed in HL biopsies.
153
HL cell lines also express high levels of VDR (Fig. 5). Lymphoma cells can synthesize vitamin D
154
and increased vitamin D levels have been repeatedly reported in HL patients.
155
In addition, seasonal fluctuations of the prognosis of HL patients have been observed and discussed as being caused by fluctuations of endogenous vitamin D levels.
156
Vitamin D can probably activate the NFKB pathway,157,158 but in most systems, vitamin D inhibits NFKB activation.159–162 In Jurkat cells with constitutive expression of VDR, no inhibition of NFKB by vitamin D was observed,
163
suggesting that some tumor cells have inactivated this pathway. Interestingly, NFKB can inhibit VDR signaling.
164
This cross-inhibition might explain the simultaneous high constitutive expression of VDR and activated NFKB in lymphoma cells. Stimulation of VDR by curcumin might shift this balance to the inhibitory activity of VDR.
Expression of VDR in HL cell lines. RNA was isolated from HL cell lines and PBMC, reverse transcribed and used as template for polymerase chain reaction with specific primers for VDR (5'-gcc ttt ggg tct gaa gtg tc-3' and 5'-cag gct gtc cta gtc agg aga t-3'). The used primers recognize all three transcript variants of the human VDR (accession numbers NM_000376, NM_001017535, NM_001017536). PCR products were sparated by agarose gel electrophoresis in the presence of ethidium bromide. NTC: no template control.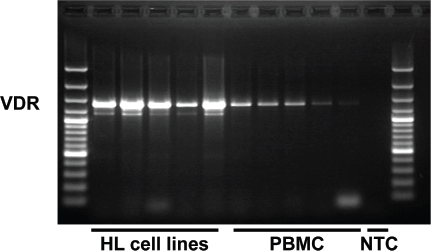
Indirect Effects of Curcumin: Immunomodulation
Curcumin can reduce the number of myeloid-derived suppressor cells, 165 an important immuno-suppressive cell population in tumors. Tumor patients, especially HL patients, often have a tumor-mediated dysfunction of the immune system. Experimental data suggest that curcumin is able to restore the immune function in tumor-bearing hosts. 166 On the other hand, induction of regulatory T cells might be increased in the presence of curcumin, 167 and the direct induction of apoptosis in activated T cells has been described. 168 As an anti-inflammatory agent, curcumin can suppress T cells.169,170
Such effects, however, might negatively affect anti-cancer immune responses. The balance between immunostimulation and -suppression might be dependent on the concentration of curcumin. 171 In the case of HL, the immune system is a double-edged sword. On the one hand, cytotoxic T cells can kill HL cells; on the other hand, T cells can provide survival signals for HL cells. The very low number of tumor cells and the presence of a high background of “normal” cells in HL indicate the importance of the interaction between tumor cells and the stroma. Whether curcumin can shift the balance between tumor promoting activities and anti-tumor activities of T cells in HL into the direction of tumor destruction must be analyzed. HL is a disease with features of chronic inflammation and the accumulation of stroma cells by HL derived cytokines is an important factor for HL biology.172,173 The anti-inflammatory activities of curcumin may be able to counteract this inflammatory stimulus.
Other Curcumin Activities Related to HL Biology
Another interesting activity of curcumin is the suppression of histone deacetylases (HDAC).174,175 Reports describe the inhibition of histon acetyltrasferases by curcumin without HDAC inhibition.176,177 In addition, curcumin has DNA methylation inhibiting activity. 178 Epigenetic regulation of gene expression by DNA methylation or histone deacetylation plays an important role in HL biology. Treatment of HL cells with HDAC inhibitors can increase the sensitivity of HL cells to cytotoxic drugs. 179 Treatment of HL cells with DNA methylation inhibitors can increase the expression of tumor antigens which might increase the recognition by the immune system.180,181 However, increased expression of factors involved in chemotherapy resistance might also occur after the treatment of HL cells with such drugs. 181
Interestingly, curcumin has been shown to inhibit transcription initiated by human immunodeficiency virus (HIV) type 1 long terminal repeats (LTR). 182 The activation of LTRs from endogenous retroviruses (ERV) in HL has been demonstrated. This activation can lead to the aberrant expression of oncogenes in HL. 183 Whether curcumin can modulate the activity of ERV promoters in tumor cells is under investigation. If ERV activity is essentially involved in the pathogenesis of HL, such ERV inhibition might reverse the malignant phenotype of the tumor cells.
A high percentage of HL carries copies of the Epstein Barr virus (EBV) in the tumor cells. Activation of the NFKB pathway by EBV seems to be involved in the pathogenesis of EBV positive HL. Curcumin can inhibit B cell immortalization by EBV.184,185 EBV-immortalized B cells undergo pronounced apoptosis in the presence of curcumin which may inhibit the outgrowth of cell lines. 185 In addition, it seems that oxidative stress promotes B cell immortalization by EBV. 186 The anti-oxidant activity of curcumin might inhibit this effect in a similar way as it was shown for vitamin E. 186 Whether this activity can interfere with the EBV driven HL pathogenesis requires further investigations.
Additional HL related activities of curcumin have been described. For example, HL cells express the anti-apoptotic BCL-XL isoform of the B cell leukemia/ lymphoma 2 (BCL2)-like 1 (BCL2L1) oncogene.187,188 Recently, a link between expression of BCL-XL and survival of HL patients has been suggested. 189 Curcumin can down-regulate BCL-XL 190 Furthermore, curcumin is a topoisomerase II inhibitor with a similar activity as etoposide, a drug which is used for treatment of HL. 191 In addition to the NFKB pathway, curcumin inhibits the STAT pathway (Table 1). This pathway is similarly important for HL cell proliferation and survival as the NFKB pathway and inhibitors of this pathway can induce apoptosis in HL cells.143,189,192–200 Constitutive activation of the STAT pathway by rearrangements of the JAK2 have been described in HL. 201 Taken together, curcumin targets several pathways in HL cells. The scarcity of the tumor cells in the tumors hampers the analysis of larger numbers of ex vivo isolated living cells. The number of established HL cell lines is very low and these cell lines are not perfectly representative for the tumor cells in vivo. Whether curcumin or curcumin derivatives with better bio-availability can be used for the treatment of HL patients requires further investigation.
Examples for DNA microarray experiments investigating the gene expression after treatment of cells or animals with curcumin or curcumin derivatives.
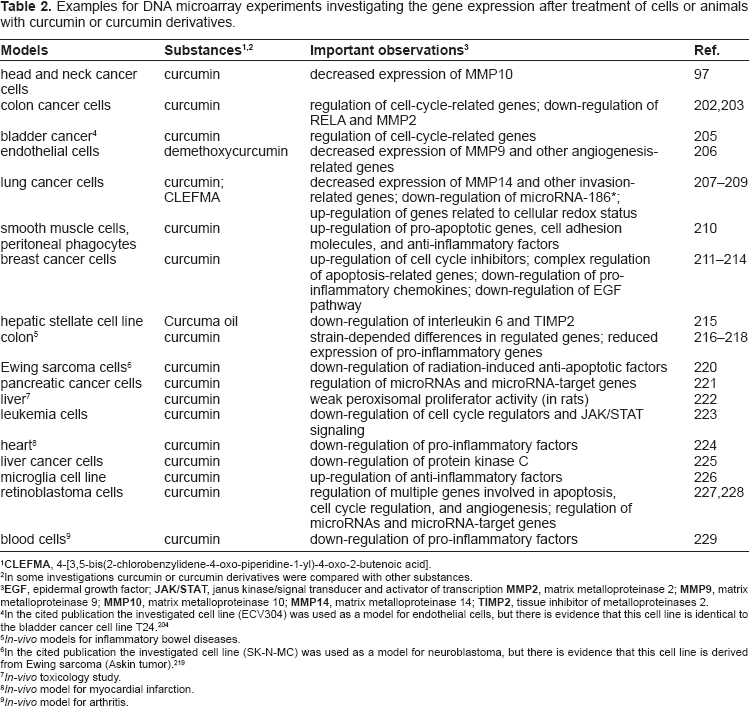
In some investigations curcumin or curcumin derivatives were compared with other substances.
In the cited publication the investigated cell line (ECV304) was used as a model for endothelial cells, but there is evidence that this cell line is identical to the bladder cancer cell line T24. 204
In-vivo models for inflammatory bowel diseases.
In the cited publication the investigated cell line (SK-N-MC) was used as a model for neuroblastoma, but there is evidence that this cell line is derived from Ewing sarcoma (Askin tumor). 219
In-vivo toxicology study.
In-vivo model for myocardial infarction.
In-vivo model for arthritis.
Future Directions
The bio-availability of curcumin is limited by the rapid metabolism and the low water solubility of this phytoceutical. Curcumin is rapidly conjugated with sulfate or glucuronic acid in the gut and liver. 230 In addition, reduction to tetrahydrocurcumin and hexahydrocurcumin occurs. 230 The resulting curcumin sulfate, curcumin glucuronide, and reduced curcumin derivatives can be found in the feces of curcumin treated patients. 231 The curcumin metabolites have some biologic activities, for example tetrahydrocurcumin can inhibit the activity of multi-drug resistance (MDR) transporters. 232 However, enhanced excretion of curcumin metabolites and the shortening of the half life of curcumin after conjugation are not desired. Inhibition of glucuronidation by piperine (the alkaloid from black pepper) has been shown to increase the bio-availability of curcumin. 233
In addition to inhibition of glucuronidation, piperine can inhibit the activity of MDR transporters. 234 On the other hand, increased expression of these transporters after exposure to piperine has been observed. 235 Together with the inhibitory effect of curcumin on MDR transporter expression and function,88,236 inhibition of MDR transporter function by piperine might counteract the effect of piperine on MDR transporter expression. In order to increase the solubility of curcumin, conjugation of curcumin with proteins or cyclodextrins have been tested.236,237 In addition, different types of nanoparticles, liposomes, and self-assembling polymers have been used for encapsulation of curcumin.238–240 Such formulations might finally overcome the bio-availability problem of curcumin.
Another interesting question that remains to be addressed is whether the effects of turmeric are only mediated by curcumin or whether additional turmeric ingredients are involved. Early studies suggest that curcumin-free turmeric extracts also have cancer preventing activities.241,242 Indeed, differences between turmeric and curcumin in the regulation of pro-inflammatory genes have been described. 243 The characterization of the factors that are responsible for biological activities of curcumin-free turmeric requires further investigations.
Author Contributions
Conceived and designed the experiments: MSS, SK. Analyzed the data: SK, IV, MSS. Wrote the first draft of the manuscript: SK, MSS. Contributed to the writing of the manuscript: SK, IV, MSS. Agree with manuscript results and conclusions: SK, IV, MSS. Jointly developed the structure and arguments for the paper: SK, IV, MSS. Made critical revisions and approved final version: SK, IV, MSS. All authors reviewed and approved of the final manuscript.
Funding
This work was supported by a fellowship from the Konrad-Adenauer-Stiftung (SK).
Competing Interests
Authors disclose no potential conflicts of interest.
Disclosures and Ethics
As a requirement of publication the authors have provided signed confirmation of their compliance with ethical and legal obligations including but not limited to compliance with ICMJE authorship and competing interests guidelines, that the article is neither under consideration for publication nor published elsewhere, of their compliance with legal and ethical guidelines concerning human and animal research participants (if applicable), and that permission has been obtained for reproduction of any copyrighted material. This article was subject to blind, independent, expert peer review. The reviewers reported no competing interests. Provenance: the authors were invited to submit this paper.
Footnotes
Acknowledgements
We thank Vera Marks (communication skills) for critically reading and copy editing the manuscript.