
Abstract
The lymphatic system has several physiological roles, including fluid homeostasis and the activation of adaptive immunity by fluid drainage and cell transport. Lymphangiogenesis occurs in adult tissues during various pathologic conditions. In addition, lymphangiogenesis is closely linked to capillary angiogenesis, and the balanced interrelationship between capillary angiogenesis and lymphangiogenesis is essential for maintaining homeostasis in tissues. Recently, an increasing body of information regarding the biology of lymphatic endothelial cells has allowed us to immunohistochemically characterize lymphangiogenesis in several lung diseases. Particular interest has been given to the interstitial lung diseases. Idiopathic interstitial pneumonias (IIPs) are characterized by heterogeneity in pathologic changes and lesions, as typified by idiopathic pulmonary fibrosis/usual interstitial pneumonia. In IIPs, lymphangiogenesis is likely to have different types of localized functions within each disorder, corresponding to the heterogeneity of lesions in terms of inflammation and fibrosis. These functions include inhibitory absorption of interstitial fluid and small molecules and maturation of fibrosis by excessive interstitial fluid drainage, caused by an unbalanced relationship between capillary angiogenesis and lymphangiogenesis and trafficking of antigen-presenting cells and induction of fibrogenesis via CCL21 and CCR7 signals. Better understanding for regional functions of lymphangiogenesis might provide new treatment strategies tailored to lesion heterogeneity in these complicated diseases.
Introduction
The idiopathic interstitial pneumonias (IIPs) are a heterogeneous group of acute and chronic disorders in which inflammation and fibrosis result from damage to the lung parenchyma, the etiologies of which remain to be fully elucidated. IIPs include idiopathic pulmonary fibrosis (IPF), nonspecific interstitial pneumonia (NSIP), cryptogenic organizing pneumonia (COP), acute interstitial pneumonia (AIP), respiratory bronchiolitis-associated interstitial lung disease, desquamative interstitial pneumonia, and lymphocytic interstitial pneumonia. 1 The former four IIPs have been of deeper concern because of their relatively higher frequencies and worse prognoses among the seven conditions. The interstitium includes the space between the epithelial and endothelial basement membranes and is the primary site of injury in the IIPs. 2 However, these disorders frequently affect not only the interstitium but also the airspaces, peripheral airways, and vessels, together with their respective epithelial and endothelial linings. 2 The inflammation and fibrosis in IIPs are characterized by spatial and/or temporal heterogeneity as typified by IPF/usual interstitial pneumonia (UIP), and these complexities have inhibited understanding of the pathophysiology in IIPs.3,4 Particularly, intraluminal fibrotic lesions called fibroblastic foci, which are present at the leading edge of fibrogenesis during the development of IPF, have been associated with poor prognoses in IPF.5–11
Lymphatic vessels form an elaborate vascular system throughout the skin and in most internal organs and play important functions in interstitial fluid drainage, absorption of lipids and small molecules, and trafficking of antigen-presenting cells into regional lymph nodes in immune responses. 12 Lymphangiogenesis occurs in adult tissues during pathologic conditions such as inflammation, wound healing, and tumor metastasis. 13 Lymphatic sprouting is mediated by vascular endothelial growth factor (VEGF)-C, VEGF-D, and VEGF receptor (VEGFR)-3, which is a receptor for both VEGF-C and VEGF-D.14–18 Lymphangiogenesis is closely associated with capillary angiogenesis.19–21 Lymphangiogenesis generally occurs after capillary angiogenesis, and lymphatic vessels grow to interdigitate blood vessels, followed by their regress in reparable conditions. 21 The balanced interrelationship between the two types of angiogenesis is considered to be essential for maintaining homeostasis. 22 Lymphangiogenesis can be either a beneficial or a detrimental phenomenon in pathologic conditions, depending on the pathophysiology of the underlying disease. Lymphangiogenesis worsens immune diseases such as the rejection response after transplantation.23–26 In contrast, lymphangiogenesis facilitates the resolution of tissue edema and the mobilization of inflammatory cells in acute and chronic inflammatory settings.27,28 Excessive lymphangiogenesis could be a cause of dehydration in interstitial tissues and can facilitate scar formation and the maturation of fibrosis. 29 Conversely, impaired lymphangiogenesis invites lymphedema,30–33 and Crohn's disease can be caused by a failure of the lymphatic vessels to transport inflammatory cells and lipids. 34
Lymphangiogenesis has been investigated in several lung diseases including airway and interstitial lung diseases, and potential roles are emerging for the measurement of lymphatic endothelial cell markers in making diagnoses, assigning prognoses, exploring the pathogenesis, and developing new treatments. 35 Particularly, growing interest has been generated regarding the relationship between IIPs and lymphangiogenesis since lymphangiogenesis is deeply linked with tissue repair. The pulmonary fibrosis associated with IIPs is hypothesized to be an aberrant healing process for tissue injury. This review provides an overview of the immunohistochemical characterization of lymphangiogenesis, the quantification of lymphangiogenic factors in IIPs and other related diseases in comparison to normal lungs, and recent advances in animal models. The possible roles of lymphangiogenesis in the pathogeneses of these conditions are discussed.
Application of Lymphatic Markers for Immunohistochemistry
The growing body of information about the expression of various markers by lymphatic endothelial cells allows the immunohistochemical identification of lymphatic vessels. Major markers for the detection of lymphatic vessels and the cells other than lymphatic endothelial cells that can express those markers are summarized in Table 1.
Lymphatic markers expressed in various types of cells in adult human and mouse lung tissues.

Podoplanin is a transmembrane mucoprotein (38 kDa), a partial epitope of which is recognized by the D2-40 monoclonal antibody. 36 Podoplanin induces the aggregation of platelets via its receptor C-type lectin-like receptor 2 (CLEC-2). 37 Podoplanin is expressed in alveolar epithelial cells, pleural mesothelial cells, and fibroblasts in the lungs of human and mouse, in addition to lymphatic vessels.36,38-41 Mice lacking podoplanin show abnormal lung development during late gestation with narrower airspaces and thicker mesenchyme. These mice fail to expand their alveolar sacs at birth, resulting in perinatal respiratory failure and mortality. 42 Mice deficient in CLEC-2 also show reduced airspace in the lung alveoli and fluid present in the larger lung airways and fail to inflate their lungs normally at birth. 42 Podoplanin is too strongly expressed by murine epithelial cells to be used as a specific marker for the detection of lymphatic vessels in lungs. 43 In human lung specimens, its expression can be varied. Anti-D2-40 antibody has been used alone for the detection of lymphatic vessels in many morphologic studies using human lung specimens because the degree of podoplanin expression does not appear to disturb the utility of anti-podoplanin antibody in lung tissues. As D2-40 is not necessarily specific to lymphatic vessels in human lungs, the identification of lymphatic vessels appears to have often been made with the assistance of information about the shape and distribution of the structures stained. Because the primary antibody against D2-40 allows robust detection of lymphatic vessels in human lung specimens, this antibody is widely used at the moment. When the detection of lymphatic vasculatures in human lungs is performed, the identification of lymphatic vessels using other markers in addition to podoplanin, such as lymphatic vessel endothelial hyaluronan receptor 1 (LYVE-1), VEGFR-3, and prospero-related homeobox transcription factor (PROX-1), is strongly recommended.29,44-46 Nevertheless, it might be not necessarily easy to determine the optimal antigen retrieval of VEGFR-3 and PROX-1 in human tissues, depending on the methods of tissue fixation.
LYVE-1 is a hyaluronan receptor that was identified through its homology with the inflammatory leukocyte homing receptor CD44. 47 The physiological roles of LYVE-1 in lymphatic vessels have not been clarified fully. It has been reported that LYVE-1 may regulate lymphangiogenesis via a stimulation driven by low molecular hyaluronic acid and the interaction with basic fibroblast growth factor (FGF-2).48,49 The expression of LYVE-1 was found to be decreased in inflammatory conditions. 50 The expression of LYVE-1 is detectable in lymphatic vessels, blood endothelial cells, and macrophages in normal human and murine lungs.43,46,51-53 In normal murine lungs, LYVE-1 is much more strongly expressed in blood vessels ranging from larger diameter vessels to capillaries, although its expression in capillaries can be decreased in inflammatory conditions. It seems to be difficult to differentiate lymphatic and blood vessels primarily by anti-LYVE-1 antibody staining in the lung parenchyma. Thus, the two types of vessels appear to be confused in murine parenchymal models, even if the expression of LYVE-1 is locally confirmed by other lymphatic markers such as VEGFR-3 and PROX-1. 43
VEGFR-3, a receptor of the lymphangiogenic factors VEGF-C and -D, is initially expressed by embryonic blood vessels during development, but its expression is subsequently restricted to lymphatic vessels once they are committed to that lineage and express additional lymphatic markers.54,55 VEGFR-3 expression can also be detected in cells of the monocyte lineage.23,52 As a detection marker for lymphatic vessels in lung models, VEGFR-3 allows the clear discrimination of lymphatic vessels from blood capillary vascularities.43,56 Based on the currently available evidence, VEGFR-3 should be chosen as the primary lymphatic marker in parenchymal models of mice. Figures 1 and 2 show VEGFR-3+ lymphatic vessels in intra-alveolar fibrotic lesions in a bleomycin-induced mouse model of pulmonary fibrosis in our study (unpublished data). LYVE-1 was robustly expressed in VEGFR-3− blood vessels and capillaries in addition to VEGFR-3+ lymphatic vessels.

VEGFR-3+ lymphatic vessels in a bleomycin-induced mouse model.
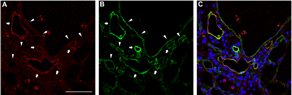
VEGFR-3+ LYVE-1+ lymphatic vessels and VEGFR-3− LYVE-1+ blood vessels in a bleomycin-induced mouse model.
PROX-1 is a transcription factor specific to lymphatic endothelial cells. PROX-1 can be expressed by the progenitor cells of monocytic lineages.24,52 Recently, PROX-1 was reported to be associated with obesity and neurogenesis.57,58 In immunohistochemistry, PROX-1 staining is detected in the nucleus. A limited number of studies have used a primary antibody against PROX-1 in morphometric analyses, 24 whereas it has often been immunohistochemically stained for the confirmation of other lymphatic markers in human and murine models.
Structure and Distribution of Lymphatic Vessels in Normal Lungs
Unlike blood vessels, lymphatic vessels have a discontinuous or fenestrated basement membrane and lack tight interendothelial junctions and are therefore permeable to interstitial fluid and cells. Through specialized anchoring filaments, lymphatic vessels stay open when the tissue pressure rises. Compared with blood vessels, lymphatic vessels are a low-flow and low-pressure system. In normal lungs, lymphatic vessels had classically been considered to be distributed in the pleura, the interlobular septa, and around the bronchovascular bundle. El-Chemaly et al. 59 reported that in normal alveolar spaces, small CD34+ capillaries were found, but no immunoreactivity for podoplanin was detected. Kambouchner and Bernaudin 60 explored the distribution of lymphatics within lobules in normal regions of human lungs using anti-D2-40 antibody. They showed the presence of lymphatic vessels not only in the connective tissue surrounding the respiratory bronchioles but also in the interlobular region, detecting lymphatic vessels with diameters ranging from 10 to 20 μm in the interalveolar walls. Sozio et al. 61 reported that most of the intralobular lymphatic vessels in normal lungs were in close contact with a blood vessel, either alone or within a bronchovascular bundle, whereas 7% were associated with a bronchiole and <1% were not connected to blood vessels or bronchioles. They additionally demonstrated an association between the shapes and sizes of lymphatic vessels and their anatomical locations. 61 The shapes of peribronchiolar and interlobular lymphatic vessels were characterized as having a marginally increased roundness and circularity and greater elongation, respectively, compared with the other type of vessel. Intralobular lymphatic size progressively decreased from bronchovascular lymphatic vessels through to the peribronchiolar, perivascular, and interalveolar lymphatics. Very limited information is available regarding the presence of the lymphatic capillaries that lie together side-by-side like beads along the alveolar septa in the intralobular region and construct an intertwined structure with blood capillaries. In this context, Ebina et al. 62 reported that lymphatic vessels were barely observed in the alveolar septa. Based on the studies that used D2-40 as a lymphatic marker, intralobular lymphatic vessels in normal lungs appear to start from the alveolar septa, possibly near venules, although the exact anatomical starting points of the lymphatic vessels in the distal site of the parenchyma have not been determined.
The amount of information available regarding the distribution of lymphatic vessels in the lungs of mice has been limited because reliable and realizable methods to recognize the lymphatic vessels have rarely been used. Baluk et al. 43 explored the anatomical distribution of lymphatic vessels in mice, mainly using PROX-1-enhanced, green fluorescence protein transgenic mice with fluorescent green lymphatic vessels and staining with anti-VEGFR-3 antibody. Their study found that most lymphatic vessels in normal lungs were located around large bronchi and major blood vessels near the hilum, while a seemingly separate lymphatic network surrounded the pulmonary veins. Lymphatic vessels accompanied airways and blood vessels into the lung parenchyma. The lymphatic network extended further distally along the branches of pulmonary veins than it did along pulmonary arteries or airways of comparable diameter. The smallest lymphatic vessels were detected on the terminal branches of pulmonary veins. Even near alveoli and visceral pleura, lymphatic vessels were typically associated with airways and blood vessels. Few lymphatic vessels were located within or beneath the visceral pleura. Kretschmer et al. 63 reported similar results in their analyses of lymphatic vessels visualized by immunostaining using anti-CD90/Thy-1 primary antibody.
Lymphangiogenesis in IIPs
Diffuse Alveolar Damage
Lymphangiogenesis has been explored in AIP/diffuse alveolar damage (DAD), IPF/UIP, NSIP, and COP. Lymphangiogenesis and capillary angiogenesis of DAD were characterized in three consecutive stages defined on the basis of histopathologic evidence, such as exudative, defined by the formation of a hyaline membrane and edema of the alveolar wall; proliferative, defined by the presence of intra-alveolar fibrosis; and fibrotic, defined by a shrunken interstitium and neighboring dilated air spaces (honeycomb formation). 64 In our study, lymphangiogenesis was characterized in the three stages of idiopathic DAD mainly using antibodies specific to podoplanin. 29 Although lymphangiogenesis was barely increased in the exudative stage, newly formed lymphatic vessels were found in the proliferative stage, primarily in the intra-alveolar fibrotic lesions. Three-dimensional images using serial sections demonstrated that some of the lymphatic vessels in the intra-alveolar fibrotic lesions had no connection to the existing ones, thus representing de novo lymphangiogenesis. Macrophages, which can potentially transdifferentiate into lymphatic endothelial cells, were expected to form de novo lymphangiogenesis by lymphovasculogenesis.24,45,52,58 In contrast, capillaries were absent from the intra-alveolar fibrotic lesions. Lymphatic vessels were not found abundantly in the alveolar septa in the exudative and proliferative stages of DAD, whereas there was an active capillary angiogenesis during these stages in the same area. Lymphatic vessels persisted in the fibrotic stage. Collectively, a remarkable unbalance was observed between the two types of angiogenesis during the development of DAD. It was speculated that lymphangiogenesis plays a role in the maturation of fibrotic tissue, facilitating tissue remodeling via fluid drainage. 29 In organizing pneumonia of acute respiratory distress syndrome-associated DAD, Mandal et al. 65 reported that the diameters of lymphatic vessels in survivors were larger than those in nonsurvivors. They speculated that the need for a greater capacity of lymphatic vessels to clear alveolar and interstitial fluids may be associated with the expansion of peribronchial lymphatics and that the obstruction of lymphatic flow might lead to unfavorable consequences through the development of intra-alveolar edema. However, it was unclear whether lymphatic density could predict prognoses in a homogeneous patient population.
IPF/UIP and NSIP
IPF is heterogeneously characterized by chronic interstitial pneumonia, intraluminal fibrosis with small aggregates of myofibroblasts and fibroblasts (termed fibroblastic foci), dense collagen, and honeycomb histological changes. 4 Fibroblastic foci consist of an extracellular matrix, an overlying epithelium, and a small aggregate of fibroblasts expressing procollagen proteins. 4 It has been considered that fibroblastic foci represent a leading edge of fibrogenesis during the development of IPF.5–9 Fibroblastic foci were reported to be a predictor of poor prognosis in patients with IPF, although conflicting data have been reported.10,11,66 Massive fibrosis is predominant in subpleural and interlobular lesions of the periphery rather than the central intralobular region. 4 On the other hand, NSIP shows a relatively good clinical response to immunosuppressive therapy, and patients with NSIP typically have a better prognosis than those with IPF. In particular, NSIP is pathologically distinguished from IPF/UIP by dense or loose interstitial fibrosis with a uniform appearance, lung architecture is frequently preserved, and fibroblastic foci are less frequently observed in NSIP than in IPF/UIP. 67 Several papers are available regarding lymphangiogenesis in IPF and/or NSIP.59,62,68-70 Lymphatic vessels have been found to be absent from fibroblastic foci and mainly detectable in the fibrotic interstitium, including fibrotic lesions around fibroblastic foci.59,68 Lymphangiogenesis has been increasingly associated with the severity of fibrosis.59,69,70 In this context, Lara et al. 70 showed the absence of definable differences between IPF/UIP and (possibly fibrosing) idiopathic NSIP, suggesting that lymphatic vessels are a unifying mechanism for the development of fibrosis in these fibrotic lung diseases. Some reports have suggested that increased lymphangiogenesis can actively facilitate fibrogenesis in IPF/UIP, although the degree of the contribution by lymphangiogenesis in whole signals associated with fibrogenesis of this condition has remained to be elucidated.59,70 CCL21 expressed in the lymphatic endothelium induces migratory and proliferative responses in CCR7+ fibroblasts.71,72 In the study by Lara et al. 70 , it was noted that the results of morphometric analyses showed considerable differences between the length and volume densities of lymphatic endothelium.
On the other hand, Ebina et al. 62 performed the characterization of lymphatic vessels among healthy controls and patients with IPF/UIP, cellular nonspecific interstitial pneumonia (cNSIP), and COP, particularly focusing on the subpleural and interlobular lesions. Using 3D images reconstructed from serial sections, they detected the destruction of subpleural lymphatics by fibrosis in IPF/UIP but not in the normal controls. In morphometric analyses, subpleural and interlobular lymphatic area and length densities and intralobular number density were decreased in IPF/UIP relative to cNSIP and COP, suggesting less lymphangiogenesis in IPF/UIP. They speculated that less lymphangiogenesis in subpleural lesions may be associated with poor prognoses in IPF/UIP via impaired alveolar clearance.
A comparative analysis of lymphangiogenesis among the subtypes of IIPs, including DAD, COP, cNSIP, and IPF/UIP, was also reported by Parra et al. 69 They estimated the amount of lymphangiogenesis by measuring the number density of lymphatic endothelial cells. DAD showed less lymphangiogenesis relative to the other IIP subtypes, although detailed information regarding the stages of DAD development was missing. On the other hand, no differences were observed among the remaining three subtypes. Although it is very important to give differential diagnoses of IPF/UIP from NSIP in clinical practice, the immunohistopathologic analyses of lymphangiogenesis generally appear to have a limited contributive value for the differentiation.
COP and cNSIP
COP has a much better prognosis than other IIPs. 73 Most patients with COP recover either spontaneously or after treatment. Organizing pneumonia is histologically characterized primarily by the involvement of alveolar ducts and alveoli with a patchy distribution.64,73 There is a mild associated interstitial inflammatory infiltrate, type II cell metaplasia, and an increase in alveolar macrophages, some of which may be foamy. There is a relatively good preservation of the background lung architecture.64,73 cNSIP is one of the three major groups described by Katzenstein and Fiorelli 67 and primarily manifests as a pure interstitial pneumonia, occasionally accompanied by organizing pneumonia. Clinically, cNSIP also shows a relatively good prognosis, similar to that of COP.
Conflicting data have been reported regarding the relative amount of lymphangiogenesis among IIP subtypes. Ebina et al. 62 showed increased lymphangiogenesis in COP and cNSIP relative to that found in IPF/UIP, whereas Parra et al. 69 reported no differences among the three subtypes. Both reports showed the appearance of new lymphatics in intraluminal fibrosis. The collective data regarding lymphangiogenesis in IIPs are summarized in Table 2.
Summary of the current reports regarding lymphangiogenesis in IIPs.

Expression of VEGF-C and -D and CCL21 in IIPs and other Conditions
VEGF-C and -D are ligands for VEGFR-3, and these signals induce lymphangiogenesis via lymphatic sprouting. Macrophages are a major source of these growth factors in inflamed tissues and fibrotic conditions. 45 An increasing body of recent information has revealed that lymphatic endothelial cells express a variety of chemokines and cell surface markers. CCL21, also known as the secondary lymphoid chemokine, which is exclusively secreted by lymphatic endothelial cells in afferent lymphatic vessels, attracts activated antigen-presenting cells that express the CCL21 receptor CCR7, and CCL21/CCR7-mediated signaling can function as an active regulator of immune responses.74,75 Information regarding the bronchoalveolar lavage fluid (BALF) levels of lymphatics-related mediators in patients with IPF was first reported by El-Chemaly et al. 59 BALF VEGF-C was significantly decreased in patients with IPF relative to that of healthy controls, while BALF VEGF-D showed no difference between the patients with IPF and healthy subjects. On the other hand, BALF CCL21 was significantly increased in patients with IPF. These results were generally confirmed by our studies. 76 El-Chemaly et al. reported that VEGF-C was immunohistochemically expressed in the airway epithelium, alveolar epithelial cells, and alveolar macrophages. In our immunohistochemical analyses, VEGF-C was remarkably expressed in alveolar macrophages near lymphatic vessels in the intra-alveolar fibrotic lesions of DAD, suggesting an important role for the participation of macrophages in the lymphangiogenesis of fibrotic diseases. 45 CCL21 expression was exclusively detected in lymphatic vessels in IPF but not in DAD. 45 In contrast, CCL19 expression was observed in lymphatic endothelial cells in DAD. 45 Janér et al measured VEGF-C levels in tracheal aspirate fluid in 54 preterm infants and reported the highest levels during the first two postnatal days. Decreased level or the immunohistochemical absence of VEGF-C in preterm infants was associated with physiologic dysplasia of the lymphatic system in the lungs. 77
In conditions other than IIPs, diagnostic values of serum and/or BALF VEGF-D have been recently determined. Some papers have reported that serum VEGF-D reflects disease activity and is a predictor of prognosis in lymphangioleiomyomatosis (LAM).78,79 LAM is an extremely rare multisystemic disease and has recently been regarded as a neoplastic disease characterized by self-multiplication of lymphatic endothelial cells. Another review provides detailed information regarding LAM and lymphangiogenesis. 80
Lymphangiogenesis in Animal Fibrosis Models
Whether lymphangiogenesis directly reflects the fibrogenesis in IIPs remains to be elucidated. Teles-Grilo et al were the first to report the existence of lymphangiogenesis in the bleomycin-induced pulmonary fibrosis rat model. After the intratracheal administration of bleomycin, new blood vessels were formed and new lymphatics were morphologically detected in the lesions associated with collagen deposition on day 21 after bleomycin exposure. 81 Meinecke et al. 82 reported that LYVE-1+ vessels exhibited ectopic mural cell coverage by smooth muscle actin-positive cells in the intraperitoneally administrated bleomycin-induced murine model. The abnormal lymphatic vascular patterning in fibrotic lungs was driven by the expression of platelet-derived growth factor (PDGF) B in lymphatic endothelial cells and signaling through PDGF receptor β in associated mural cells. They speculated that the drainage capacity of pulmonary lymphatics is a critical mediator of fibroproliferative changes. Wilder et al. 83 characterized LYVE-1+ vessels in radiation-induced murine fibrosis models. The LYVE-1+ vessel density was estimated at 1, 4, 8, and 16 weeks after bleomycin exposure and was decreased in the bleomycin-exposed animals relative to that of the controls.
Several Issues in Exploring the Relationship between IIPs and Lymphangiogenesis
Several issues should be discussed regarding the characterization of lymphangiogenesis in IIPs and its pathogenic roles. The first is the possibility of methodologic bias in morphometric analyses, as mentioned above. 70 It was noted that considerable differences between the length and volume densities of lymphatic endothelium were observed in severe fibrosis. Lymphangiogenesis persisted in dense or severe fibrosis as represented in idiopathic DAD and IPF/UIP, while capillary angiogenesis regressed.29,84 Active lymphangiogenesis beyond capillary angiogenesis would induce a decrease of interstitial pressure in the tissues, resulting in the collapse of lymphatic vessels even in active lymphangiogenic conditions. Because the volume density of the lymphatic endothelium likely creates bias for the estimation of lymphangiogenesis via the influence of interstitial pressure, length density should also be measured at the same time. In addition, these densities should be standardized by interstitial density because the deviation of airspace can create bias in the measurement of these densities (Table 2).
Furthermore, an unbalance between capillary angiogenesis and lymphangiogenesis might be linked to the prognosis of pulmonary fibrosis. Several reports showed the association between the presence of lymphangiogenesis without capillary angiogenesis and prognoses in IIPs.62,65,68 However, as summarized in Table 3, it is difficult to determine whether lymphangiogenesis in the alveolar septa and intraluminal fibrosis gives a fair or biased insight into pulmonary fibrosis associated with interstitial pneumonia. Lymphangiogenesis was active in intraluminal fibrosis of DAD but inactive in IPF/UIP. The two conditions commonly show poor prognoses. In addition, it was also active in COP, which generally has a good prognosis.
Heterogeneous characteristics of lymphangiogenesis in principal fibrotic lesions of IIPs.

In fibrosis in the alveolar septa, similar inconsistency was also observed between IPF/UIP and DAD. On the other hand, several reports are available regarding capillary angiogenesis in IIPs. Blood vessels were reported to be absent in the fibroblastic foci of IPF, 85 whereas new capillaries were found to appear in the same lesions in the later pathologic stage, albeit to a lesser degree than in healthy tissue. 68 In contrast, capillary density was increased in mild interstitial fibrotic lesions but decreased with the advancement of fibrosis. 86 In DAD, the capillary density was increased in the alveolar septa in the exudative stage, followed by regression in the fibrotic stage, and blood vessels were absent from the intra-alveolar fibrotic lesions in the proliferative stage. 29 The imbalance between capillary angiogenesis and lymphangiogenesis is common to both IPF and DAD. In contrast, both blood and lymphatic vessels are detectable in intraluminal fibrosis in COP.69,87 The discrepancy between the two types of angiogenesis might be related to poor prognoses of IIPs based on the results provided by immunohistochemistry, although capillary angiogenesis remains to be explored in interstitial lesions in COP and NSIP. This hypothesis would need to be confirmed by morphometric analyses.
The heterogeneities in the affected area and pathologic changes inhibit understanding of the roles of lymphangiogenesis in IIPs. Lower levels of lymphangiogenesis appear to be related to undesirable events in inflammatory lesions because of dysfunctional fluid drainage, and excessive lymphangiogenesis is also evidently associated with massive fibrosis. IIPs are occasionally accompanied with lymphoid hyperplasia in addition to inflammation and fibrosis, and dendritic cells (DC) have been reported to be associated with IPF. 88 Recent reports have demonstrated an association between bronchus-associated lymphoid tissue and lymphangiogenesis. 43 Lymphangiogenesis also has the potential to contribute to the immune response in IIPs. Although animal models are needed to clarify the general roles and define new functions of the lymphatic system in fibrogenesis, they might be inadequate for determining the localized roles of lymphangiogenesis in these heterogeneous and complex fibrotic diseases in humans. In the future, the development of useful methods for regionally investigating the function of lymphatic vessels might help to achieve a better understanding of the various roles of lymphangiogenesis in IIPs. Schematic images regarding the possible roles of lymphangiogenesis in IIPs are shown in Figure 3.

A schema explaining the possible roles of lymphangiogenesis in IIPs.
Lymphangiogenesis in Granulomatous Lung Diseases
Sarcoidosis
Sarcoidosis is a systemic granulomatous disorder of unknown etiology involving multiple organs. Inhalation of some kind of airborne antigens has been assumed to be the cause for this disease because >90% of the studied sarcoidosis cases have been found to involve the lungs with presentation of small, reticular, nodular shadows or bilateral hilar lymphadenopathy in chest radiographs. 89 Noncaseating sarcoid granulomas are an organized collection of macrophages and epithelioid cells and are frequently covered by a ring of fibrosis. Particularly in the lungs, sarcoid granulomas are usually distributed along lymphogenous routes such as the alveolar septa and the bronchovascular bundle, in which existing lymphatic vessels are generally located.89,90
Two studies have reported findings on lymphangiogenesis in patients with sarcoidosis.79,91 Kambouchner et al. 91 reported the important information that lymphatic vessels were observed in approximately 68% of intralobular sarcoid granulomas in lung specimens obtained from patients with pulmonary sarcoidosis, implying that sarcoid granulomas were more strongly associated with lymphatic vessels than with blood vessels. In our study, consequently, the serum and BALF concentrations of VEGF-A and VEGF-C were significantly increased in patients with pulmonary sarcoidosis relative to those of healthy controls and patients with IPF/UIP. 79 The results of immunohistochemical analyses demonstrated that VEGF-A and -C were expressed in sarcoid granulomas. Immunostaining with anti-podoplanin antibody for the detection of lymphatic vasculatures showed the presence of usual lymphatics and atypical tubular structures around sarcoid granulomas. Atypical tubular structures were characterized by a thin membrane, with weak expression of podoplanin and a membrane deficit in a part of the borderline. The structures were observed in ∼58.6% of 193 granulomas, whereas usual lymphatic vessels were limited in 15.6% of those granulomas. A portion of the tubular structures was connected to CD73+ afferent lymphatic vessels. 79 Two types of cells were observed within the lumens of atypical tubular structures: one type with positivity for S100 protein, a pan DC marker, and the other type with positivity for CD1a, a marker of a subset of myeloid DCs. It was speculated that atypical tubular structures are involved in the trafficking of antigen-presenting cells and the antigen itself into regional lymph nodes through a connection to the usual lymphatic vessels.
Lymphangiogenesis in Airway Lung diseases
Bronchial Asthma
Asthma is a chronic inflammatory disease of the airways that occasionally presents as a life-threatening condition. Ebina 92 characterized the lymphatic vessels in airway walls using specimens obtained from autopsied patients with fatal asthma. The distribution of lymphatic vessels, as estimated by the area density, was decreased in the airway walls consisting of muscle bundles and fibroconnective tissues. They speculated that the augmentation of these tissues in airway walls disrupts lymphatic vessels, resulting in the impairment of airway clearance and the acceleration of mucosal edema. Thus, the dysfunction of lymphatics causes the refractory status of these patients. However, patients with near-fatal asthma often fall into generalized dehydration. 93 The possibility that the decreased area density of lymphatic vessels in fatal asthma is a result of generalized dehydration was not excluded.
Chronic Obstructive Pulmonary Disease
Chronic obstructive pulmonary disease (COPD) is a chronic, inflammatory lung disease with high mortality and morbidity. 94 Long-term smoke exposure is the primary cause for developing COPD. Several reports are available regarding COPD and lymphangiogenesis.46,95 Mori et al rigorously reported the characteristics of lymphangiogenesis in COPD. In their report, the severe stages of COPD were associated with increases in the standardized lengths and number densities of alveolar lymphatic vessels. 46 Increased numbers of CCL21+ and D6+ lymphatics were observed in the alveolar parenchyma of subjects with advanced COPD compared with those in controls. It was hypothesized that lymphangiogenesis was involved in the trafficking of distal lung immune cells in advanced COPD. It would be interesting to know how this increase in the number of lymphatic vessels in the advanced stages of COPD is linked to the comorbidity caused by this condition. It was noted that the anti-D6 antibody used in this study might be a useful marker for immunohistochemically evaluating the function of lymphatic vessels in human lungs. Shiels et al. 96 reported that the serum levels of soluble VEGFR-3 (sVEGFR-3) in current and past smokers were significantly increased relative to the levels detected in those who have never smoked, although the pathogenic relevance of this finding has been unclear.
Experimental Obliterative Bronchiolitis
Obliterative bronchiolitis (OB) is the pulmonary manifestation of chronic transplant rejection and remains the leading cause of morbidity and mortality after lung transplantation.97,98 Krebs et al. 26 reported a role for lymphangiogenesis during the development of experimental OB in rat tracheal allografts. Adenovirus vectors were used in this study to overexpress and inhibit VEGF-C. VEGF-C overexpression in tracheal allografts induced intense inflammation followed by enhanced lymphangiogenesis and the development of OB. In contrast, the inhibition of VEGF-C activity inhibited lymphangiogenesis and reduced the infiltration of CD4+ T-cells and the development of OB. Lymphangiogenesis was linked to T-cell responses during the development of OB, and VEGF-C/VEGFR-3 signaling modulated innate and adaptive immune responses in the development of OB in this model, consistent with the previous reports related to transplantation. 25
Animal Infectious Models
Several reports are available regarding airway infection and lymphangiogenesis in mouse models, although there is limited information on this topic in humans. Baluk et al studied the roles of lymphangiogenesis in the airways using mouse models of chronic respiratory tract infection with Mycoplasma pulmonis by adenoviral transduction of airway epithelium with sVEGFR-3. Lymphangiogenesis robustly occurred in the infected airways. Inhibition of VEGFR-3 signaling by the intravenous adenovirus delivery of sVEGFR-3 completely prevented the growth of lymphatic vessels as estimated using anti-LYVE-1 and anti-VEGFR-3 antibodies. Lack of lymphatic growth exaggerated mucosal edema and reduced the hypertrophy of draining lymph nodes. 27 It was noted that after antibiotic treatment of the infection, the inflammation and remodeling of blood vessels quickly subsided, but those of lymphatic vessels persisted. Xu et al. 99 reported protective roles of cathepsin L in reducing severity in acute and chronic airway infection models and its involvement in active lymphangiogenesis.
On the other hand, Nihei et al. 56 reported a relationship between lymphangiogenesis and worsened inflammation in aspiration pneumonia models. In their model, food suspended in phosphate-buffered saline that was adjusted to low pH (pH 1.6) using pepsin was given together with lipopolysaccharide as an intranasal challenge daily 5 days a week, for a maximum of 28 days. Lung inflammation and active lymphangiogenesis were increasingly observed during the 28-day challenge period. The administration of inhibitors of lymphangiogenesis including axitinib and SAR131675 decreased the lymphatic area density, as estimated by anti-VEGFR-3 antibody in lungs, and improved oxygenation in an aspiration pneumonia model relative to that of controls. It was speculated that increased lymphangiogenesis worsened the inflammation in this model. The results appear to be inconsistent with those provided by the previous two reports in terms of the causal relationship between lymphangiogenesis and inflammation, which remains to be fully elucidated.
Conclusion
Lymphangiogenesis plays various roles in lung diseases, including interstitial fluid drainage, absorption of small molecules, clearance of inflammatory cells, and trafficking of antigen-presenting cells into regional lymph nodes in immune responses. In IIPs, lymphangiogenesis is likely to have different types of localized functions within each disorder, corresponding to the heterogeneity of lesions in terms of inflammation and fibrosis. In the future, it is expected that new assessment tools, potentially including staining for CCL21 and D6, can help to define the localized functions of lymphatic vessels. In addition, research on lymphangiogenesis in IIPs might allow the heterogeneity of these complicated diseases to be better understood in terms of the regional functions of lymphatic vessels.
Author Contributions
Conceived the concepts: MY. Analyzed the data: MY. Wrote the first draft of the manuscript: MY. Contributed to the writing of the manuscript: MY. Agree with manuscript results and conclusions: MY. Developed the structure and arguments for the paper: MY. Made critical revisions and approved final version: MY. The author reviewed and approved of the final manuscript.