
Abstract
Synthetic patches and fixed grafts currently used in the repair of congenital heart defects are nonliving, noncontractile, and not electrically responsive, leading to increased risk of complication, reoperation, and sudden cardiac death. Studies suggest that tissue-engineered patches made from living, functional cells could grow with the patient, facilitate healing, and help recover cardiac function. In this paper, we review the research into possible sources of cardiomyocytes and other cardiac cells, including embryonic stem cells, induced pluripotent stem cells, mesenchymal stem cells, adipose-derived stem cells, umbilical cord blood cells, amniotic fluid-derived stem cells, and cardiac progenitor cells. Each cell source has advantages, but also has technical hurdles to overcome, including heterogeneity, functional maturity, immunogenicity, and pathogenicity. Additionally, biomaterials used as patch materials will need to attract and support desired cells and induce minimal immune responses.
Congenital Heart Defects
Congenital heart defects (CHD) are the most common birth defects (3 per 1000 live births) and are the leading cause of birth defect-related death.1,2 CHD can range from life threatening to benign. Benign CHD, such as septal defects, the most common CHD (>50% of all CHD), are often underdiagnosed because of lack of symptoms, 3 and only an estimated 20% of patients with septal defects will require surgical intervention. 4 Other CHD, such as hypoplastic left heart syndrome (2.7% of all CHD) or Tetralogy of Fallot (4% of all CHD) have life-threatening symptoms and require surgical intervention. 3 When treating patients with CHD, surgical repairs often require patches to close septal defects or enlarge stenosed structures, for example, ~50% of Tetralogy of Fallot repairs include a patch for the right ventricular outflow tract. 5 Currently, polymers (polytetrafluoroethylene [PTFE], Dacron, etc.), fixed xenografts (eg, bovine pericardium), and fixed pericardium autografts are the most common patch materials used in CHD repairs. 6 However, because these materials are synthetic or treated to remove living cells, they do not grow with the patient nor are they electromechanically integrated; moreover, the patches may become fibrotic at the site of implantation and have mismatched mechanical properties compared to the surrounding tissue. As CHD are often treated early in development, additional surgeries are sometimes required to replace overgrown patches, and risks of arrhythmia and sudden cardiac death are increased in patients with patch implants.7,8 The 25-year survival rate of patients who underwent repair with patch implantation is 80% compared to 93% in patients who underwent repairs without patch implantation. 9 Without cells, the patch cannot grow with the patient, potentially requiring further surgeries to replace it as it is outgrown; 14% of Tetralogy of Fallot patients require reoperation on the right ventricular outflow tract. 10 The lack of contractility and mismatched mechanical properties could lead to loss of cardiac function and output. The lack of electrical conductivity could potentially cause a reentrant loop of depolarization around the patch area, leading to a self-sustained depolarization pattern, which could lead to arrhythmia, fibrillation, and further complications. The combination of these various shortcomings may contribute to the elevated risk of sudden cardiac death in patients implanted with patches, which is 25 to 100 times the norm. 11 A tissue-engineered patch that contains cardiac cells with contractility and conductivity matching the native tissue could grow with the patient and recover cardiac function.12,13 This patch could be seeded with cardiac cells or cardiac progenitor cells (CPC), potentially derived from the patient's own stem cells, in a bioengineered material designed to incorporate with native tissue and facilitate healing.
Cardiac Development and Homeostasis
Cardiomyocytes (CM) are derived from the mesodermal lineage through the interaction between several signaling factors, including activin, bone morphogenetic proteins (BMP), and transforming growth factor β (TGF-β).14,15 Through the activation of mesendoderm differentiation by activin, BMP signals, and inhibition of the canonical Wnt pathway, the cardiac lineage is derived, as indicated by the expression of Nkx2.5, an early cardiac transcription factor.14,15 Nkx2.5 further initiates the expression of another early cardiac transcription factor, GATA4, and these transcription factors upregulate the expression of key components of the CM, including cardiac actin filaments, myosin heavy chains, and cardiac troponin. 14 Myofibrillogenesis is the formation and organization of the sarcomere during CM differentiation and maturation. Beginning in the precardiac mesoderm stage, actin and myosin filaments independently form and give rise to stress fibers. Initially, these fibers are composed mostly of nonmuscle actin and myosin subunits. As differentiation occurs, cardiac-specific actin and myosin subunits begin to replace their nonmuscle counterparts. Formation of anchoring proteins as well as filament alignment takes place as the cells begin to contract. As the cells mature, the myofilaments increase in alignment and most nonmuscle filament proteins are replaced. 16
Supporting cells, including cardiac fibroblasts and connective tissues, are also important in maintaining cardiac functions. Cardiac fibroblasts comprise the largest population (>50%) of cells in the myocardium.17,18 They help stabilize cardiac tissue by maintaining extracellular matrix (ECM) homeostasis, providing electromechanical support, and participating in wound healing. 19 Fibroblasts are responsible for secreting collagen types I, III, IV, and IV, generating the collagen-rich ECM that provides mechanical stability to cardiac tissues.17,20 Fibroblasts also produce growth factors that regulate cardiac cells, including TGF-β, angiotensin II, IL-6, and IL-1β. 21 Cardiac fibroblasts provide passive resistive force to the cardiac tissue, and the electrical connection between cardiac fibroblasts and CM facilitates the propagation of action potentials throughout the cardiac tissue. 17 Cardiac fibroblasts participate in wound healing responses such as fibrosis when ischemia or other damage occurs in the heart. 22 Cardiac fibroblasts can be identified by their expression of discoidin domain receptor 2, a collagen receptor. 17 Vascular tissues such as endothelial cells (EC) and smooth muscle cells (SMC) are also found in cardiac tissue; they are a small portion of the 70% non-CM cell population in the heart.17,18 They primarily participate in the vascularization necessary to support cardiac tissues, providing a pathway for oxygen, nutrients, and signaling cytokines. 23
Application of Stem Cells for Chd Treatment
A tissue-engineered patch encapsulated with CM and support cells with properties and functions similar to native tissues would integrate with healthy tissues to restore functions while avoiding the risk of arrhythmia.24,25 However, because native CM do not normally proliferate, alternative sources for CM are needed. Several sources of CM have been investigated in current literature, including CM derived from embryonic stem cells (ESC),26,27 induced pluripotent stem cells (iPSC),28,29 mesenchymal stem cells (MSC), 30 adipose-derived stem cells (ADSC), 31 umbilical cord blood cells (UCBC), 32 amniotic fluid-derived stem cells (AFSC), 33 and CPC34–36 (Table 1). As potential sources of CM in a cardiac patch for pediatric CHD repair, the cells need to be minimally allergenic while being able to develop sufficient force generation and electrical coupling.
Summary of CM cell sources discussed, ESC, iPSC, MSC, ADSC, UCBC, AFSC, and CPC.
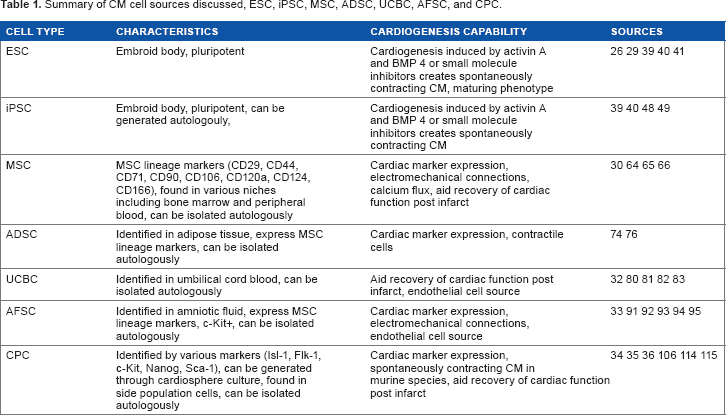
Embryonic stem cells
ESC with the capability to differentiate into all three germ layers have been heavily investigated in the last two decades.37,38 Early research had shown that spontaneously beating CM can be found in embryoid body culture of ESC with low yield (<10%). 26 Kattman et al showed that optimized treatment of activin A and BMP4 can differentiate ESC to beating CM with better yield (>50%). 39 Recently, groups have shown that small molecule inhibitors of Wnt signaling can induce highly efficient (>85%) CM differentiation in embryonic-like stem cells.29,40 However, the leftover undifferentiated ESC could form teratomas.37,38 ESC-derived CM are functionally immature; myofilaments in ESC-derived CM do not align in a preferential direction, thus do not generate the level of force found in mature CM. However, Lundy et al recently reported that late-stage (80-120 days of differentiation) ESC-derived CM exhibited improved filament organization and force generation compared to early-stage (20–40 days of differentiation) ESC-derived CM. 41 Additionally, fibrin gel containing CD15+, Isll+ ESC-derived progenitor cells is currently being tested for use in ischemic heart failure recovery in an ongoing clinical trial.42,43 Variability between different lines of ESC may also affect the differentiation efficiency of ESC and cause heterogeneity in the resulting cells. 44 Furthermore, ESC would not be autologous to the patient, resulting in risks of immune reactions. 45
Induced pluripotent stem cells
iPSC were first generated from differentiated cells by the Yamanaka group with the transfection of a combination of transcription factors, including Oct3/4, Sox2, Nanog, Klf4, and Myc.46,47 These transcription factors act to reprogram differentiated cells to revert back to a pluripotent stage resembling ESC. iPSC reprogramed from human fibroblasts have been shown to possess the ability to form teratoma containing all three germ layers when injected in mice. 47
iPSC have been shown to differentiate into CM through protocols similar to ESC-derived CM. 48 Namely, iPSC can be induced to differentiate toward mesoderm lineage through inhibiting GSK3 or embryoid body formation. Further inhibition of the canonical Wnt signaling pathway in these mesoderm fated iPSC will lead to differentiation of functional CM. Recently, highly efficient CM differentiation protocols have been identified using both growth factor Wnt pathway manipulation39,49 and small molecule inhibition of the Wnt pathway.29,40 Introducing chemically defined small molecule inhibitors of Wnt at specific time points, the Palecek lab was able to induce greater than 85% pure iPSC-CM. 40
iPSC are readily available from commercial sources, typically generated from fibroblasts. 50 The advantage of iPSC over ESC is that iPSC could be generated autologously from one's own tissue; iPSC can even be generated from cells isolated from patient's amniotic fluid.51,52 Patient iPSC derived CM can also be used as in vitro models of cardiac disorders.53,54 However, iPSC pose the same potential for teratoma formation in patients if undifferentiated cells remain with the desired differentiated ones. While there are many advantages of personalized iPSC lines, patient to patient variability may be a concern, requiring additional consideration in controlling the differentiation efficiency and heterogeneity in the resulting cells. Additionally, studies of iPSC showed different immunogenic responses in their syngeneic hosts depending on the implantation site.55–57
Mesenchymal stem cells
MSC are adult stem cells identified by the expression of CD29, CD44, CD71, CD90, CD106, CD120a, CD124, and CD166 with the presence of integrins α1, α5, and β1.58–60 MSC have been isolated from various niches, including bone marrow and peripheral blood.61–63 The ability of MSC to differentiate into CM has been disputed. Studies have observed MSC expressing cardiac lineage proteins such as cardiac troponin and GATA4 when injected in cardiac infarct models.64,65 Groups have also shown that MSC could establish electromechanical connections with native cells in host cardiac tissue in animal models.30,66 Xu et al found rhythmic calcium flux and potential electrical activities in MSC cultures treated with 5-azacytidine to prevent DNA methylation. 30 However, consistent generation of functional MSC-derived CM has yet to be shown. MSC have been shown to differentiate into endothelial and perivascular phenotypes and have been shown to improve cardiac function in animal cardiac infarct models.67–69 While MSC could be isolated autologously, the frequency and differentiation potential of MSC varies between patients and decreases with age and morbidity.70,71 While MSC were shown to be nonimmunogenic in experiments in vitro, recent studies have shown that MSC could still be immunogenic in vivo, suggesting that MSC may be less immunoprivileged than previous literatures suggested.57,72
Adipose-derived stem cells
ADSC are cells similar to MSC isolated from adipose tissue.73–75 ADSC can differentiate into various phenotypes similar to MSC. 75 Mouse ADSC have been reported to generate spontaneous beating cells expressing cardiac genes. 74 Human ADSC have been shown to upregulate cardiac genes when cultured on laminin 76 and generate small populations of beating cells when exposed to rat CM extracts. 31 ADSC could be isolated autologously. However, similar to MSC, the consistency and purity of ADSC are highly variable, and the methods for generating CM from ADSC have yet to demonstrate clinically relevant numbers.
Umbilical cord blood cells
UCBC are a group of heterogeneous cells found in the umbilical cord that have been shown to possess multipotent differentiation potential.77–79 Populations of UCBC have been shown to help recover cardiac function when implanted on infarction sites in animal models.80,81 UCBC can serve as an endothelial source.82,83 However, differentiating UCBC into CM phenotypes resulted in limited functioning CM. 32 Owing to the heterogeneic nature of UCBC, strict selection methods are required to limit variability and improve reproducibility of results. 79
Amniotic fluid-derived stem cells
Amniotic fluid (AF) provides protection for the developing embryo by dampening movements to the amniotic cavity. AF contains a mixture of several cell types from the fetus.84,85 A population of the cells found in the AF was found to express Oct-4, a transcription factor found in pluripotent stem cells.86,87 Another population of the cells found in the AF was identified to express c-kit (or CD117), a surface marker found in multipotent stem cells in adults. 88 These c-kit positive cells, referred to as AFSC, have been shown to possess osteogenic, myogenic, and adipogenic potential similar to MSC, but do not form teratomas in mice.85,89
AFSC, as a potential therapeutic component, offer many advantages, especially in congenital defect repairs. When defects are diagnosed during pregnancy, AF could be collected and AFSC could be isolated. These AFSC would be autologous and pose lower risk of immune rejection than allogeneic sources. AFSC can be isolated from as early as the first trimester and expanded to clinically relevant numbers for perinatal or neonatal surgical applications. 85 Methods of inducing CM differentiation in AFSC have been investigated, which include coculture mixing of AFSC with cardiac cells of various sources. 33 When cocultured directly with neonatal rat ventricular myocytes (NRVM), AFSC have been shown to express cardiac troponin I. 33 When cocultured indirectly with NRVM, AFSC have been shown to form functional gap junctions and improve cell-cell connectivity. 90 AFSC have yet to be shown to generate spontaneously contracting CM.
AFSC can be differentiated into EC with the commercially available media Endothelial Growth Medium 2 (EGM-2) supplemented with basic Fibroblast Growth Factor (bFGF) or Vascular Endothelial Growth Factor (VEGF).91–93 A viable method of high-efficiency AFSC endothelial differentiation has been developed by a member of our lab 94 and formed comparable vessel structures to human umbilical vein EC control in fibrin/PEG hydrogels. 95
Cardiac progenitor cells
Even though most CM are terminally differentiated in the developed heart, efforts have been made to identify and study CPC. CPC are proliferating cells with the ability to differentiate into cardiac cells when triggered by cytokine signals or other factors.96,97 Some CPC are identified by their ability to elute Hoechst dye; these cells are referred to as “side population” cells.98,99 The surface markers Isl1,100,101 Flk-1, 102 c-Kit, 36 and Nanog 103 were also used to identify CPC from human patient samples.100,101 CPC are found to be activated by electrical stimulation in vitro 104 and cardiac infarction in adult patients. 36 Promising methods of identifying and differentiating CPCs have been identified with rodent sources. 105 Cardiac cells expressing Sca-1 have been identified in cardiac niches in neonatal and adult murine hearts that can aid in regenerating cardiac function post infarction and potentially differentiate into CM.36,106–108 Groups have even generated functional beating cells from murine CPC.108,109 Additionally, studies have reported that murine CPC can help improve cardiac functions in postinfarction models. 110 However, experiments with human CPC have yet to show beating CM in vitro.
Alternatively, phase-bright cells migrating out of explants of cardiac tissue samples have also been shown to form dense colonies or cardiospheres. 111 These cardiosphere-derived cells (CDC) contained potential CPC that expressed markers of pluripotency and were able to divide in in vitro and in vivo conditions. 111 CDC were able to differentiate into multiple germ layers in vitro and even exhibited myocardial differentiation when implanted in rat heart. 34 Mishra et al have shown that CDC can be isolated from samples taken from pediatric CHD patients. 35 They noted that age correlated negatively with the amount of CDC harvest and their proliferation abilities. 35
A study has identified a population of c-kit positive cardiac cells that has shown CM differentiation in infarcted areas. 36 When isolated from human patients and injected into immune compromised mice, CPC were found to have developed connections to the native mouse CM and begin to differentiate into different cardiac lineages.112,113 Other studies with Hoechst stain eluding “side population” CPC also have shown cardiac differentiation capabilities. 105 CDC containing CPC have been shown to be multipotent in vitro. 34 Implanted CDC in a rodent cardiac infarct model have been shown to differentiate into CM, fibroblast, and vascular cells, and CDC derived CM were able to integrate with host CM. 35 Compared to control, CDC implantation led to improved cardiac tissue regeneration and functional recovery. 35 Matsuura et al have shown beating cells derived from mouse CPC cultures. 108 The presence and quality of CPC decrease with age; therefore, CPC may be a more relevant source of generating autologous cardiac cells for pediatric applications as greater amount of CPC could be isolated from younger patients. A variety of niches for potential CPC have been identified. Therefore, extensive characterization and strict selection criteria must be established prior to clinically relevant application. 114
Using Stem Cells in Conjunction with Biomaterials
Cell-loaded biomaterials
Stem cells could be incorporated into biomaterials and function as tissue replacements. Tissue-engineered systems can be tested in various in vivo models of cardiovascular defects such as the mouse angiogenesis model, 115 rat Left Ventricle (LV) ischemia model, 16 rabbit cardiac infarct model, 116 baboon infarct model, 117 and hemodialysis model. 118 Engineered tissues could have passive functions: scaffolds containing bone marrow MSC have been tested as functional vascular grafts with successful implants in ongoing clinical trials12,119 and fibrin hydrogels containing skeletal myoblasts have been shown to reduce the thinning of the ventricular cell walls postinjury in a rat LV ischemia model. 16 A vascular autograft composed of autologous fibroblast and EC has been shown to achieve 76% patency in patients in a clinical trial. 120 Tissue-engineered grafts could also provide active functions. Shin et al examined a gelatin methacrylate gel containing carbon nanotube and seeded with NRVM reported that it was able to act as a bioactuator, generating forces in vitro when stimulated with electrical signals. 121 Biomaterials loaded with stem cells could also be used as stabilizing or regenerative agents. Wang et al reported that injectable poly (ethylene glycol)-poly (caprolactone) copolymer (PEG-PCL) hydrogels containing bone marrow MSC have significantly higher vessel growth in the infarcted areas compared to control in a rabbit cardiac infarct model. 116 However, because of the multipotent nature of stem cells and because biomaterials are derived from native tissue or containing heterogeneous natural molecules, the resulting material could have variable behavior. 122 One method of reducing variability and toxicity of biomaterials is to use cells of definite phenotype or predifferentiated stem cells. 123 At a cost of plasticity and differentiation potentials, the behavior of the material could be more controllable.
Biomaterials that recruit cells
Biomaterials can also be designed to recruit native cells. By adjusting the mechanical properties, such as elasticity, compressibility, density, and degradation rate, biomaterials could be selective in the adhesion and proliferation of specific types of cells.124,125 Recently, a silk-fibronectin hydrogel has been shown to recruit native endothelial population by enhancing the bioactivity of a porous synthetic material (PDLLA) by adding silk-fibronectin, resulting in increased vascular ingrowth. 126 Biomaterials could also be designed to promote the migration and proliferation of specific types of cells through chemical means with preloaded growth factors, biomolecules, or cells.125,127 A MSC-loaded hydrogel has been shown to induce angiogenesis in vivo with significantly higher invading vessel segments compared to acellular controls when injected subcutaneously in a mouse model. 115
Biomaterials conditioned with cells
Furthermore, biomaterials could be pretreated with cells, and then decellularized. The resulting material would benefit from the remodeling and ECM deposited by the cells, improving biocompatibility and cell recruiting abilities of the material. Removing the cells reduces the concern of controlling cell behaviors postimplantation and host immune responses. Syedain et al generated arterial grafts with tubular fibrin gels conditioned with human fibroblast and underwent mechanical stretching; the authors reported that the decellularized grafts withstood burst pressures at 50% or higher of ovine femoral arteries control. 128 A study has shown that tissue-engineered fibrin heart valves conditioned with fibroblasts have similar mechanical properties compared to allograft controls and have improved recellularization potential in vitro. 129 Weber et al reported that similar fibrin gel heart valves conditioned with human fibroblast and dynamically stretched showed improved in vivo recellularization in a baboon model compared to decellularized allograft controls. 117 Dahl et al generated a tissue-engineered vascular graft with SMC cultured in PGA; after decellularization, the graft was examined in a baboon hemodialysis model and found to have 83% patency and lower T-cell adhesion compared to PTFE control after 24 weeks. 118
Future Considerations
Several stem or progenitor cell sources have been investigated for their ability to generate cardiac cells. ESC and iPSC have been shown to generate functional CM.39,40,49 MSC expressed CM phenotype markers when injected in myocardium in in vivo models.30,64,65 CPC have been shown to express a CM phenotype in in vitro experiments, and murine CPC have been shown to generate spontaneously beating cells.34,35,108,109 Other stem cell sources, such as ADSC, UCBC, and AFSC, have been able to express CM phenotype markers when induced by mature CM in vitro or in vivo.31–33 Current studies have yet to produce CM with mature phenotype capable of generating force at the level of native CM. 41 Generating CM at clinically relevant numbers remains another challenge. 18 Purity and immunogenicity of the resulting CM should also be closely monitored.44,45 With respect to CHD and pediatric patients, one should also consider the lasting effects of materials and cells that may remain in the patients for years, which may increase risk of certain side effects such as the formation of teratomas in pluripotent stem cell-derived cells37,38 or the toxicity of degradation byproducts. 130 The pediatric heart is also at a different stage developmentally compared to the adult heart, which may result in different immune responses and regenerative potentials.17,18 Material design and choice of cells will require consideration of these challenges. Designing biomaterials to improve the proliferation and function of stem cells should be considered.121,125 Conditioning biomaterials with stem cells is a different approach that has seen promising results recently.117,128,129 So far, promising results have been shown in generating cardiovascular tissues from various stem cell sources in combination with biomaterials; however, many challenges still have to be met in tissue engineering constructs for surgical repairs of CHD.
Footnotes
Author Contributions
Wrote the first draft of the manuscript: YG. Contributed to the writing of the manuscript: YG, JGJ. Agree with manuscript results and conclusions: YG, JGJ. Jointly developed the structure and arguments for the paper: YG, JGJ. Made critical revisions and approved final version: JGJ. Both authors reviewed and approved of the final manuscript.