
Abstract
Human pluripotent stem cells (PSCs) are a promising cell source for cardiac tissue engineering and cell-based therapies for heart repair because they can be expanded
Introduction
Heart failure is the leading cause of death in most industrialized countries and a growing cause of death in emerging countries. 1 The most common cause of heart failure is myocardial infarction (MI), which is a loss of myocardium because of oxygen and nutrient deficiency and replacement with non-contractile scar tissue. A primary challenge in developing therapies for cardiac muscle repair is the limited regenerative capacity of the myocardium. 2 While recent research has established that the adult heart contains a sub-population of multipotent stem cells capable of heart regeneration, 3 only 1% at most of the cardiomyocytes are replaced annually and thus cannot repair the significant cardiac damage that occurs during MI. Currently, the only long-term treatment available to restore cardiac function after Mi-related heart failure is heart transplantation. 2 However, in the United States, <2,500 heart transplantations are performed annually because of limited organ availability and ~10% of patients die while waiting for a donor heart. 4 Further, there are a large number of patients who do not qualify for heart transplant but could still benefit from heart repair, which could be a significant portion of the ~290,000 people who die in the USA each year because of heart disease. 4
Cardiac tissue engineering and cell-based therapies, including direct cell injection, are potential approaches for treating cardiac disease. Cell injection is a straightforward method that directly injects cells into the target tissue to restore its function. Cardiac tissue engineering seeks to engineer myocardium
Human cardiomyocytes derived from pluripotent stem cells (PSCs) are the only viable source for new cardiomyocytes currently available. The reason is that adult cardiomyocytes are non-proliferative, and adult stem cells are difficult to differentiate into cardiomyocytes. The development of human embryonic stem cells (ESCs) first enabled the generation of cardiomyocytes in large numbers for cell-based therapies and tissue engineering applications. 13 However, ESC-derived cardiomyocytes are allogeneic with the potential for immunogenicity and there are also ethical concerns regarding the use of human embryos as a cell source. 14 Both of these points have been addressed by the development of induced pluripotent stem cells (iPSCs), derived from somatic cells by reprograming their transcriptional profile into a pluripotent state. 15 Patient-specific cardiomyocytes can easily be derived by reprograming adult somatic cells into iPSCs and then differentiating these into cardiomyocytes.
ESCs and iPSCs have the potential to form a teratoma if injected directly into the heart, 16 and thus must be differentiated into cardiomyocytes prior to implantation. Cardiomyocyte differentiation protocols were first developed for ESCs 17 and then adapted to iPSCs using established ESC protocols. 15 Since that time, PSC differentiation protocols have undergone substantial improvements, making the process cheaper, faster, more reliable, and more efficient. Here we review the most recent advances in differentiating human PSCs toward cardiomyocytes with particular focus on PSC monolayer-based approaches that have proved to be the most efficient, consistent, and scalable techniques currently available. Further, we will discuss current approaches to promote maturation of the PSC-derived cardiomyocytes and the primary challenges related to the use of these cells for cell-based therapies and tissue engineering.
Comparison of iPSC and ESC Differentiation
Conceptually, the differentiation of ESCs into terminally differentiated cardiomyocytes is no different than that of iPSCs. The differentiation of both PSC types relies on modulating signaling pathways that guide embryonic development
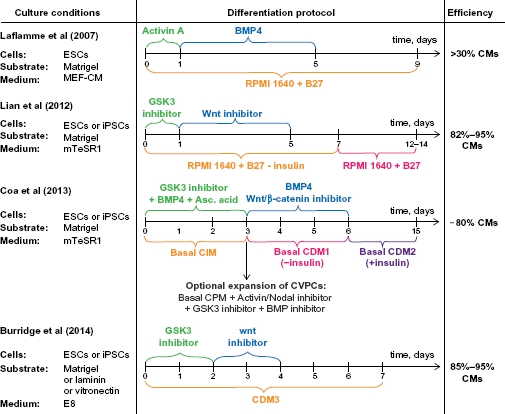
Timelines of PSC monolayer-based differentiation methods. Culture conditions are shown in the left column and differentiation efficiency in the right column. Timelines show information about media used for differentiation (below time axis) and the additional components added to it (above time axis). Information about inhibitors used in each method can be found in Table 1. The last time mark indicates the beginning of spontaneous contractions of the derived cardiomyocytes.
Interestingly, there are studies showing that the differences in transcriptional profiles between ESCs and iPSCs are minimal and insignificant, 21 while other studies indicate the opposite. 22 This would suggest that cardiomyocytes derived from ESCs and iPSCs may show differences in phenotype because of differences in gene expression. Supporting this, Zhang et al. 23 found that ESC-derived cardiomyocytes have a faster beat frequency and a larger percentage of spontaneously beating cells. They also reported that depending on the method used for the iPSC induction, iPSC-derived cardiomyocytes can have residual expression of pluripotency transgenes OCT4 and NANOG, which may cause undesired effects, including teratoma formation and dedifferentiation. Another study by Lee et al. 24 showed that Ca2+ handling of human ESC-derived cardiomyocytes is much closer to the adult phenotype compared to more immature Ca2+ handling of human iPSC-derived cardiomyocytes. However, in contrast, Gupta et al. 25 reported that despite differences in transcriptional profiles between ESCs and iPSCs, transcriptional profiles of both types of PSC-derived cardiomyocytes were surprisingly similar. It was also shown that both types of PSC-derived cardiomyocytes have comparable sarcomere organization, responsiveness to chemical stimuli, and ability to differentiate into cardiomyocyte subtypes (eg, ventricular, atrial, and nodal). 23 Finally, it was shown that iPSC differentiation efficiency as well as the maturity of the iPSC-derived cardiomyocytes can be significantly influenced by the somatic cell type that those iPSCs were derived from. 26 In particular, iPSCs derived from cardiomyocytes tend to differentiate more efficiently into cardiomyocytes, forming more mature cells compared to iPSCs derived from other tissues. 27 Thus, history of the iPSC-derived cardiomyocytes needs to be taken into account when comparing cell phenotypes or gene expression profiles. These results suggest that iPSCs could successfully replace ESCs as a cell source for cardiac tissue engineering and cell-based therapies; however, the possible risks of using iPSC-derived cardiomyocytes, such as de-differentiation, trans-differentiation into the initial donor cell type, and risk of teratoma formation, still need to be further studied.
Co-Culture- and Embryoid Body (EB)-Based Methods of PSC Differentiation into Cardiomyocytes
Current methods for PSC differentiation into cardiomyocytes can be divided into three categories: (i) differentiation via co-culture with mouse visceral endoderm-like (END-2) stromal cells, (ii) EB differentiation in suspension, and (iii) 2D monolayer differentiation. 18 The first method requires culturing PSCs and END-2 stromal cells together or culturing PSCs in END-2-conditioned medium and relies on the signaling molecules produced by END-2 stromal cells to direct differentiation. While it was one of the first methods developed, it is limited in utility by a low cardiomyocyte yield (less than 10%, but usually around 1%) 28 and an immature cardiomyocyte phenotype. An additional limitation is that the mechanism of differentiation is poorly characterized, as it is driven by partially unknown chemical factors produced by the END-2 cells. However, in 2007, Graichen et al found that the cardiomyocyte yield can be increased to >20% upon addition of p38 MAPK inhibitor, making this method much more efficient than EB differentiation. 29 Another advantage of human PSC differentiation by END-2 cell co-culture is its simplicity and inexpensiveness compared to the EB-based methods. 30
EB differentiation relies on a combination of physical and chemical cues to modulate cell signaling pathways and direct PSCs into cardiomyocytes. EBs are formed by growing PSCs as small, spherical aggregates that mimic early embryonic development. Depending on the protocol used and the variability between the experimental repeats, culture of the EBs overtime results in differentiation of PSCs toward various cell types where 5%-70% of EBs contain beating cardiomyocytes. 31 An advantage of the EB approach is the option of differentiating PSCs under fully defined conditions. Disadvantages of the EB method include inconsistent results between experiments in terms of the number of EBs that contain beating cardiomyocytes, 31 low cardiomyocyte yield (often <1%), 32 and immature cardiomyocyte phenotype. For these reasons, researchers have been exploring alternatives to EB differentiation, specifically looking to PSC monolayer differentiation techniques.
PSC Monolayer-Based Differentiation
Monolayer differentiation of PSCs, much like EB differentiation, is based on the use of growth factors and small molecules added to the medium to induce differentiation down the cardiac lineage. The monolayer technique, however, differs in that instead of forming 3D EBs, PSCs are grown in a 2D monolayer, which allows for the scalable differentiation of a large quantity of cells. Additionally, the monolayer method produces more mature cardiomyocytes that show signs of subtype specification (ventricular, atrial, or nodal cardiomyocytes). 33 Over the past three years, a number of 2D monolayer differentiation techniques have been developed that can be applied to a large variety of PSC lines, with cardiomyocyte yields as high as 85%-95% (Fig. 1) and a reduction in cost by reducing the number of media components.
Differentiation of human PSCs into cardiomyocytes from a confluent monolayer was first reported for human ESCs (cell line H7) in 2007. 32 In this example, ESCs were cultured on Matrigel in MEF-conditioned medium until a high-density cell monolayer was formed and then switched to chemically defined RPMI-B27 medium. Over the first 24 hours, the medium was supplemented with human recombinant Activin-A, followed by a four-day supplementation with human recombinant BMP4, until finally cultured in pure RPMI-B27 for two to three weeks. Spontaneous beating of cardiomyocytes was typically observed on the 12th day of differentiation, and the final cell population consistently contained >30% cardiomyocytes (Fig. 1). At the time of development, this technique was the most efficient one, outperforming EB-based techniques in terms of efficiency by an order-of-magnitude. However, later studies found that this monolayer method produced inconsistent results for different cell lines and experimental repeats. 34
In 2010, it was found that the mechanism of the Activin/BMP4 differentiation is dependent upon endogenous Wnt/β-catenin signaling. 34 Particularly, Wnt/β-catenin signaling is stimulated in the beginning of differentiation and then inhibited after the formation of mesoderm. Furthermore, suppressing this signaling at early stages or stimulating it after the mesoderm formation significantly reduced cardiomyocyte yield. Understanding the role and timing of Wnt/β-catenin signaling has led to improved monolayer differentiation protocols. Lian et al.35,36 demonstrated in 2012 that stimulation of Wnt/β-catenin signaling through the addition of the GSK3-inhibitor CHIR99021 in the beginning, and its suppression (either chemically or by β-catenin shRNA expression) after the mesoderm formation, is sufficient to consistently generate cardiomyocytes. Additionally, Lian et al showed that insulin, which is a part of B27 medium supplement, suppresses cell differentiation during the first five days. After insulin was removed from the differentiation medium, the consistency of the results increased dramatically. Importantly, an improvement in this technique was that both culture and differentiation of the human PSCs occurred under fully defined conditions (Table 1) and were validated for three lines of ESCs and three lines of human iPSCs with cardiomyocyte yields of 82%–95% (Fig. 1). A major advantage of this approach was consistent differentiation of cardiomyocytes in large quantities with high purity. The authors also demonstrated the importance of understanding the role of media components used for differentiation. However, this approach still contained some media components in the B27 supplement with unknown effects on differentiation efficiency and phenotype of the derived cardiomyocytes.
Comparison of PSC monolayer-based methods used for cardiomyocyte differentiation.
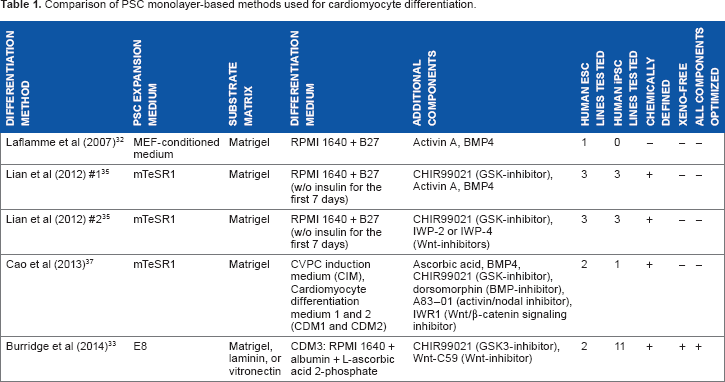
The differentiation of PSCs to cardiomyocytes has also resulted in investigation of intermediate states of specification and their use in monolayer-based protocols. Cao et al. 37 used human ESC and iPSC lines to generate cardiovascular progenitor cells (CVPCs), which were expanded up to 15 passages and then further differentiated into cardiomyocytes (Fig. 1). CVPCs were generated using a GSK3-inhibitor CHIR99021, BMP4, and ascorbic acid and then maintained in the progenitor state by blocking BMP, GSK3, and Activin/Nodal pathways with the corresponding inhibitors (Table 1). Differentiation of CVPCs into cardiomyocytes was achieved by adding BMP4 and the Wnt/β-catenin signaling inhibitor (IWR1) and removing insulin from the medium. This approach allows for cell expansion at the progenitor stage, cryopreservation of CVPCs for future use, and differentiation into three CVPC-derived cell lineages: cardiomyocytes, endothelial cells, and smooth muscle cells. Furthermore, the CVPCs were shown to be non-tumorigenic, and thus have the potential to be used directly for cellbased therapies. However, this technique is more labor intensive and time consuming compared to Lian et al.35,36 and requires the use of the B27 supplement, where the effects of some chemical factors on differentiation remain poorly defined.
Recent work has focused on using fully defined media with known components in order to reduce the number of and understand the effect of each component on PSC differentiation to cardiomyocytes. The goal is to improve differentiation efficiency across different PSC lines into specific cardiomyocyte subtypes. In 2014, Burridge et al. 33 reported development of E8 media containing only eight ingredients to replace the more expensive mTeSR1 media and development of CDM3 differentiation media containing only three components to replace the previously used RPMI 1640 + B27 media (Table 1). The differentiation protocol was similar to Lian et al. 36 ; cells were first exposed to a GSK-inhibitor (CHIR99021) for two days, followed by a Wnt-inhibitor (Wnt-C59) for the next two days, and then CDM3 media alone for the remainder of the process (Fig. 1). The researchers also investigated the role of the substrate extracellular matrix protein coating on human PSC culture and differentiation and found that growth factor-reduced Matrigel, laminin-511, and laminin-521 achieved reliable pluripotent growth and long-term attachment during differentiation. Vitronectin was also found to be a cheaper substitute to the laminins, but cells needed to be passaged at least every 15 days to prevent detachment. This difference is explained by cell adhesion to laminin-based matrices using α6β1 integrin compared to αvβ5 integrin used predominantly for vitronectin. Importantly, this differentiation protocol was successfully applied to 11 human iPSC lines and 2 human ESC lines from passage 20 to 80, with cardiomyocyte purity consistently more than 85%. Further, gene expression and electrophysiology of the derived cardiomyocytes at 30th–35th days of differentiation revealed that ~57% of cells express a ventricular-like phenotype. The overall result is a reliable, efficient (85%–95% purity), and relatively inexpensive human PSC cardiomyocyte differentiation protocol using E8 and CDM3 media with predictable maturation and subtype specification. Other known alternatives to mTeSR1 include StemPro (Invitrogen), 38 Pluripro (Cell Guidance Systems),39–41 PluriSTEM (Millipore),41,42 and Nutristem (Stemgent)41,43; however, not all of these media have been tested for cardiac differentiation.
Maturation of PSC-Derived Cardiomyocytes
Cardiomyocytes derived from PSCs can begin contracting after seven days of differentiation, 33 but have an immature phenotype. To date, the most direct way to mature PSC-derived cardiomyocytes is to culture them for prolonged amounts of time (1+ year). 44 It has been determined that PS-derived cardiomyocytes >35 days old, though it depends on the differentiation technique used, show significant maturation. Specifically, their phenotype compared to immature cardiomyocytes includes (i) loss of proliferative ability, (ii) elongation, (iii) action potential profile becoming subtype specific (eg, atrial, ventricular, and nodal), (iv) change in gene expression profile, and (v) increase in beat frequency. 11 In addition to time, maturation of PSC-derived cardiomyocytes has also been achieved through electromechanical stimulation,45,46 treatment with tri-iodo-L-thyronine, 47 transgenic expression of cardiac-specific proteins such as calsequestrin, 48 and co-culture with non-cardiomyocytes. 49
Yet despite the advances in maturation, even the most mature PSC-derived cardiomyocytes are more similar to fetal than to adult cardiomyocytes. Robertson et al. 11 have compiled a comprehensive review, comparing the changes in structural and functional properties between embryonic, fetal, neonatal, and adult cardiomyocytes. Summarizing its key conclusions, PSC-derived cardiomyocytes have a fetal-like phenotype because they have a smaller length-to-width aspect ratio (3:1 compared to 15:1), are mononuclear, have fewer mitochondria, and have poor sarcomere organization. Also, the global gene expression profile of PSC-derived cardiomyocytes is closer to embryonic than adult cardiomyocytes. Finally, functional characteristics of PSC-derived cardiomyocyte have altered Ca2+ handling, low Ca2+ capacity of sarcoplasmic reticulum, low beat rates (~40 BPM), immature action potential characteristics, abnormal levels of ionic currents, negative force-frequency relationships, and abnormal expression of sarcoplasmic reticulum proteins, all indicative of a fetal phenotype. Thus, the challenge moving forward is developing strategies, not just to generate PSC-derived cardiomyocytes, but also to mature them to adult phenotypes so that they can be used for cellular therapies and engineering cardiac tissues with the potential to integrate with native heart tissue and demonstrate physiologic contractile function.
Future Directions
PSC-derived cardiomyocyte differentiation techniques have undergone a number of significant improvements. Earlier methods aimed at mimicking cell environment
However, there are still many challenges that require further research before therapeutic application of these cells can be realized. First, human PSC-derived cardiomyocytes have an immature phenotype. Therefore, factors that can direct cardiomyocyte maturation still need to be further investigated and validated. While some progress has already been made, a full mechanistic understanding of cardiomyocyte maturation has yet to be determined. Second, PSC-derived cardiomyocytes are believed to have higher risk of de-differentiation and teratoma formation compared to primary cells or adult stem cells. 19 This is particularly a concern for iPSCs, where the pluripotency induction method used can result in unwanted transgene expression in the cells. More studies are needed to determine which iPSC induction methods result in clinically safe, differentiated cells. Thus, while differentiation of human PSC-derived cardiomyocytes has achieved a number of major advances, there is still significant work to be done to elucidate all the possible risks related with using them in human cell therapies and developing approaches to eliminate or mitigate these.
Footnotes
Author Contributions
Wrote the manuscript: IB, AWF. All authors reviewed and approved the final manuscript.