
Abstract
In aquatic environments, genotoxicity results from the effects of pollution combined with the inflammatory response triggered by the immune system. Indeed, the production of nitrosylated DNA and proteins are though to arise from the production of peroxinitrite during phagocytosis and inflammation. The purpose of this study was to examine new DNA biomarkers that differentiate between immune- and pollution-mediated genotoxicity in wild clam populations. Intertidal clam populations were sampled and analyzed for gonadal DNA strand breaks, DNA nitrosylation and xanthine oxidoreductase (XOR) activity (purine salvage pathway). The clam weight-to-shell-length ratio, the gonado-somatic index (GSI), age status, lipid peroxidation, xenobiotic conjugation activity (glutathione S-transferase (GST) and phagocytic activity were examined to shed light on their relationships with the observed genotoxic endpoints. XOR activity and DNA strand breaks were generally elevated at polluted sites and correlated significantly with clam weight-to-shell-length ratios and DNA nitrosylation. DNA nitrosylation was also higher at some sites and correlated significantly with phagocytic activity and with DNA strand breaks. This study showed that DNA strand breaks were associated with both immune-and pollution-mediated effects. This suggests that there is a loss of DNA repair capacity due to the combined effects of aging, pollution and immune response in wild clam populations that are impacted by anthropogenic activity.
Keywords
Introduction
The St. Lawrence estuary, which receives inflows from the Fjord du Saguenay, is influenced by various sources of pollution, including harbours, marinas and wastewater discharges from municipal and industrial sources such as pulp and paper mills. The chemical contamination of this intertidal area is well documented and includes the usual contaminants, such as polycyclic aromatic hydrocarbons (PAHs), heavy metals, organotin compounds, and the so-called endocrine disruptors found in urban waste. 1 Clams readily accumulate PAHs by ingesting contaminated food or sediment. 2 Some PAHs, especially those in the heavy molecular class (e.g. benzo[a]pyene), are biotransformed by cytochrome P450-based enzymes in a process that can lead to oxidative stress and genotoxicity. 3 Both PAH-contaminated sediment and clams are commonly found in the Fjord du Saguenay area, especially near harbours and municipal effluent outfalls.4,5
Although DNA damage is caused by pollution, it can also result from sustained inflammation in the fight against infection.
6
During the process of phagocytosis, ingested bacteria are internalized into phagosomes, fused to lysosomes and readily destroyed by the concomitant production of nitric oxide (NO) and hydrogen peroxide, which yield the highly reactive biocide peroxynitrite.
7
The release of NO in phagocytosis was also associated with the expression of interleukin-2 (IL-2), which plays a role in the production of NO.
8
For example, exposure to bacteria in the oyster
The production of NO during an oxidative burst will not only kill bacteria and viruses; it can also damage host cells by reacting with endogenous proteins (thiol S-nitrosylation) and DNA (DNA nitrosylation). DNA can also undergo oxidative damage (7,8-dihydro-8-oxo-2′-deoxyguanosine) from the release of hydrogen peroxide and other oxidants, and this damage is therefore the result of both inflammatory and oxidative conditions, and pollution-related effects (e.g. through DNA adducts or production of reactive oxygen species).
DNA damage is usually determined by measuring DNA strand breaks and alkali-sensitive sites,10,11 neither of which can discriminate between pollution and disease-mediated effects. The development of biomarkers capable of such discrimination would be of value in determining the cumulative effects of pollution, disease and inflammatory conditions, hence in identifying the etiology of DNA damage. The basal levels of DNA strand breaks in marine invertebrates have been attributed to a large number of alkali-labile sites that are characteristic of tightly packaged DNA in cells. 12 Interestingly, strand breaks were reduced by the antioxidant Trolox, EDTA and an inhibitor of Ca2+/Mg2+-dependent nucleases. Biomarkers designed to measure the oxidation and nitrosylation of DNA, along with the breakdown or salvage of purine released by DNA strands, might provide information leading to a better understanding of the role of pollution and disease in the genotoxicity of marine invertebrates.
The purpose of the present study was to examine the contribution of DNA nitrosylation to strand-break formation and salvaging (purine catabolism) in order to provide a better understanding of the cumulative impact of the immune response and pollution on genotoxicity in feral clam populations. This goal was achieved by measuring changes in DNA nitrosylation, purine salvage metabolism and phagocytic activity in relation to the formation of DNA strand breaks in wild
Materials and Methods
Study area
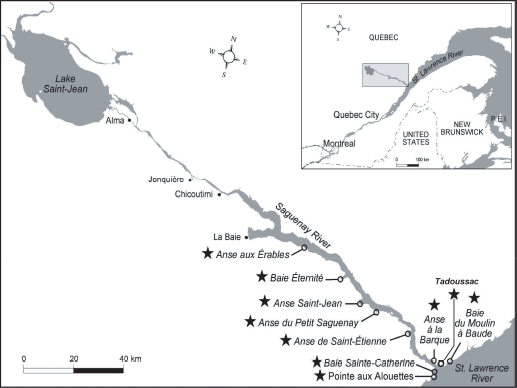
Spatial distribution of feral clam populations. Clams were collected at 10 sites (identified by stars) in June 20055. The sites at Anse de Saint-étienne (ASE) and Baie du Moulin à Baude (BAU) were used as the reference (pristine) sites, while the sites at Anse aux érables (AE), Pointe aux Alouettes (PAL), Baie éternité (BE), Anse Saint-Jean (ASJ), Petit Saguenay (PS), Anse à la Barque (BAR), Baie Sainte-Catherine (BSC) and the town of Tadoussac (TAD) were used as the pollution-impacted sites.
Characteristics of sampling sites in terms of their emplacement and potential for contamination.
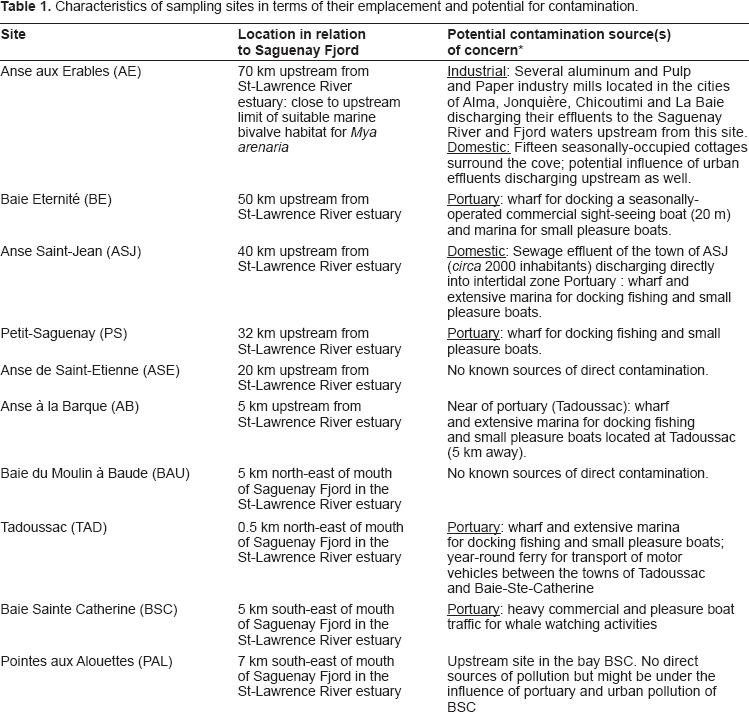
The Fjord du Saguenay sites included the Anse étienne (ASE) site, which was used as the reference site for the fjord, and was located on the south shore 20 km upstream from the fjord mouth. The Petit Saguenay (PS) site is located 32 km upstream on the south shore of the fjord mouth and has a small marina for seasonal activities and pleasure boating. The Anse Saint-Jean (ASJ) site is exposed to minimally treated (i.e. screened) urban wastewaters from a population of about 2,000 residents and is located 40 km upstream from the fjord mouth. The Baie éternité (BE) site is located 50 km upstream from the fjord mouth. It has a moderately sized marina under 10 m in length that accommodates circa 20 ships, and is used for recreational and tour boat activities. Finally, the Anse-aux-érables (AE) site, the site furthest upstream in the fjord (70 km upstream from the fjord mouth), is located downstream from aluminum smelters and pulp and paper mills, and from four towns that discharge their wastewaters into the fjord.
Clam collection and tissue preparation
Intertidal clams (N = 20) were collected at a depth between 10 cm to 20 cm, frozen on dry ice and placed in a −85 °C freezer to await analysis. Before freezing, 1 mL of hemolymph was collected from the anterior adductor muscle of each clam for on-site phagocytic determinations. Specimens of similar size (6 to 8 years of age) from each site were dissected for biomarker measurements.
The biomarker tests were performed using the gonad tissues, which are in very close contact with openly circulating hemolymph. 13 The gonad tissues were dissected out and homogenized in 10 mM of Hepes-NaOH buffer (pH 7.4) containing 125 mM NaCl, 1 mM EDTA and 1 mM dithiothreitol at 4 °C at a 1:5 weight/volume ratio. Aliquots of the homogenate were taken for total protein content, 14 lipid peroxidation (LPO), DNA damage, DNA extraction (DNA nitrosylation) and xanthine oxidoreductase activity (XOR) determinations. Before the clams were frozen, a portion of each gonad was dissected out for sex determination, maturity indexes and the presence of tissue damage from infection or parasitic infestation.
DNA from digestive glands and gonadal tissues was extracted using the method developed by Goldenberger et al. 15 Briefly, the homogenate was mixed with the extraction solution containing the detergent sodium dodecylsulfate (SDS) and 1,000 units/mL of proteinase-K and was extracted using phenol and chloroform and the aqueous phase (DNA and RNA) was precipitated with two volumes of cold ethanol (30 min at −20 °C). The resulting pellet was dried and resuspended in 10 mM Tris-acetate, pH 8.0, and 1 mM EDTA and then precipitated again in three volumes of ice-cold ethanol and resuspended in 1 mL of Tris-acetate-EDTA buffer. The extracted DNA was tested using the purity index (the A260 nm/A280 nm absorbance ratio). A 1.8 ratio for extracted DNA samples was judged satisfactory. If a sample had a ratio <1.8, then the sample was extracted again with phenol and chloroform, as described above. DNA was quantified by absorbance at 260 nm (Beckman DV 70 spectrophotometer) with standard solutions of salmon sperm DNA (Sigma Chemical Company).
Dna damage evaluation
The integrity of the genetic material was studied by examining nitrosylation and DNA strand-break assays. The level of DNA damage from the production of peroxynitrite during inflammation was determined by a novel DNA nitrosylation assay, based on the reduction of NO2 adducts to DNA using the enzyme diaphorase that catalyzes the reduction of R-NO2 to R-NH2 in the presence of NADPH. A solution containing 50 μL of DNA and 50 μL of diaphorase (10 U X mL−1 in 100 mM KH2PO4, pH 7.4) was mixed and preincubated for 5 min at 30 °C. The reaction was initiated with the addition of 50 μL of NADPH (100 μM) and the formation of NADP+ was determined by the loss of fluorescence at 340 nm excitation/460 nm emissions at 0, 5, 15, 30 and 60 min intervals. The data were expressed as the loss of NADPH fluorescence * mg−1 DNA * min−1.
DNA strand breaks were quantified by fluorescence using an alkaline precipitation assay.16,17 The levels of single- and double-stranded DNA remaining in the supernatant were assessed by mixing 50 μL of the supernatant with 200 μL of Hoescht dye at a concentration of 1 μg mL−1 in 200 mM Tris-acetate, pH 8.5, containing 300 mM NaCl and 4 mM sodium cholate. 18 Fluorescence was then measured using 360 nm (excitation) and 450 nm (emissions) filters. The blanks contained identical constituents without homogenate, and salmon sperm DNA standards were added for DNA quantification. The results were expressed as μg of supernatant DNA * mg−1 of homogenate proteins.
The elimination of purines from DNA turnover was determined by xanthine oxidoreductase activity according to the method described by Zhu et al 19 50 μL of the S15 fraction sample of mussels was added to 125 μL of assay buffer (50 mM KH2PO4, pH 7.4, containing 10 μM sodium molybdate), hypoxantine 50 μM and 10 μM dichlorofluorescein diacetate. The formation of fluorescein was measured at 0, 10, 20, 30 and 40 mn, at 30 °C, using 485 nm (excitation) and 516 nm (emissions) filters. The results were expressed as relative fluorescence units/(min-mg protein).
Phagocytosis, oxidative stress and xenobiotic conjugation
Phagocytic activity, hemocyte viability and cell adherence were evaluated as described elsewhere. 20 For phagocytosis, 25 μL of fluorescein-labelled bacteria (corresponding to 5 × 107 bacteria/well) were added to the wells and left to incubate for 2 hr at room temperature. The wells were then washed once with PBS as described above and resuspended in 100 μL of 0.125 mg/L of Trypan Blue, pH 4.4 (50 mM sodium-citrate), to quench the fluorescence of any traces of remaining bacteria adhering to the cell walls. 21 Fluorescence was measured at 485 nm for excitation and 520 nm for emission (Chameleon II, Bioscan, USA). Phagocytic activity was expressed as relative fluorescence units/mg proteins, as determined, by the fluorescamine method. 22 Briefly, 130 μL of 30 mM NaOH was added to the plates for 30 min to lyse the cells. Then 70 μL of 500 μg/mL fluorescamine (in 100% acetonitrile) was added and the plate was left to stand for 10 minutes. Fluorescence was measured at 400 nm for excitation and 450 nm for emissions. Standards of fluorescein (phagocytosis and cell viability) and bovine serum albumin (proteins) were used for calibration.
The level of thiobarbituric reactive substances (TBARS) resulting from oxidative lipid peroxide breakdown (LPO) was evaluated with the thiobarbituric method. 23 Blanks and standards of tetramethoxypropane (stabilized form of malonaldehyde) were prepared in the presence of the homogenization buffer. Fluorometric analysis was performed at 540 nm (excitation) and 600 nm (emissions). The results were expressed as μg of TBARS:mg–1 of homogenate protein.
Glutathione S-transferase (GST) activity was determined by the method of Boryslawskyj et al 24 using 2,4-dichloronitrobenzene as the substrate in a microplate format. The assay was performed on the 15,000 × g supernatant at 0.1 mM of the substrate, and absorbance readings were made at 340 nm. Data were expressed as the change in absorbance/min/mg.
Data analysis
Ten clams were collected from each site for biomarker analyses. Homogeneity of variance and normality of the data were verified using Levene's test. The data were log-transformed when significant deviations from normality occurred. The data were analyzed by a factorial two-way analysis of variance (two-way ANOVA) with gender and site as the factors. For inter-site correlation analysis of individual morphology and biomarker responses, a Pearson-moment correlation test was used. A factorial analysis was also performed using the principal components extraction procedure to determine the relationships among the biomarker data. The residual value of DNA strand breaks were calculated against DNA nitrosylation to extract the component of DNA strand breaks that was not explained by DNA nitrosylation (i.e. from inflammation). The residual is defined as the absolute difference between the predicted the levels of DNA strand breaks from the linear relationship with DNA nitrosylation and the actual measured value. Significance was set at p < 0.05.
Results
The morphological characteristics of clams collected in the Fjord du Saguenay and St. Lawrence estuary was examined (Fig. 2). In the St. Lawrence estuary, the weight-to-shell-length ratio (condition factor) was significantly reduced at sites subject to anthropogenic activity (Fig. 2A). A two-way ANOVA revealed that site location but not gender was a determining factor. Clams at the TAD and BAR sites were the most severely affected, with the weight-to-shell-length ratio decreasing 1.6 times. The ratios for clams in the Fjord du Saguenay at the ASJ and AE sites were generally less affected, decreasing by factors of 1.6 and 1.2 respectively. The gonado-somatic index (GSI) was also affected at the study sites with no apparent sex-related effects (Fig. 2B). In the St. Lawrence estuary, the GSI was reduced at all polluted sites, especially the BSC and TAD sites, with reductions by factors of 2.7 and 3 respectively, compared with the reference site (i.e. BAU). In the Fjord du Saguenay, GSI increased by a factor of 1.4 at the AE site and decreased by a factor of 1.9 at the ASJ site. Correlation analysis (Table 2) revealed that the weight-to-shell-length ratio was significantly correlated with GSI (r = 0.46; p < 0.01). Analysis of clam age distributions revealed that age status varied from 5 to 9 years of age and was significantly correlated with GSI (r = 0.46; p = 0.001).
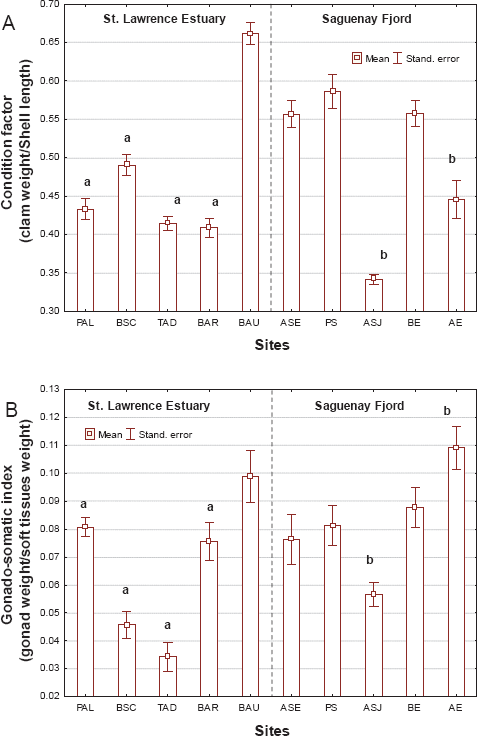
Change in morphological characteristics of
Correlation analysis of the biomarker data.

The physiological health status of the clams was also determined by tracking the extent of GST activity, and LPO and phagocytic activity (Figs. 3A to 3C). GST activity was readily induced at the polluted TAD and BAR sites in the St. Lawrence estuary (increasing by a factor of 1.4 at the BAR site), with no apparent gender-related interactions (two-way factorial ANOVA: p < 0.01 for site and p > 0.1 for gender) (Fig. 3A). In the Fjord du Saguenay, GST activity was readily induced at all impacted sites, increasing by a factor 2.7 at the site receiving municipal wastewater (i.e., site ASJ).
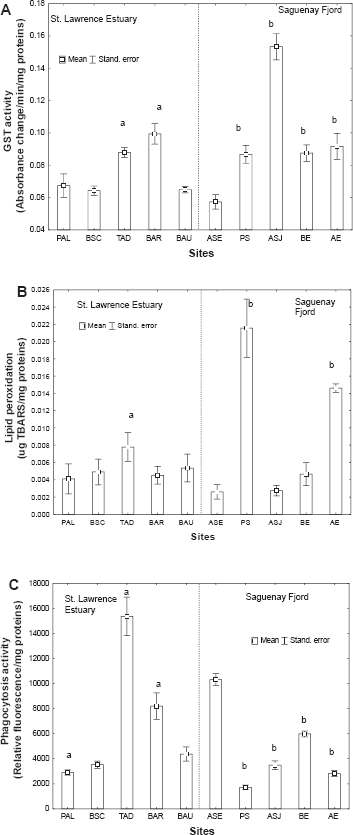
Xenobiotic metabolism, oxidative stress and phagocytic activity in feral
The extent of gonad LPO was also examined in the wild clams (Fig. 3B). A two-way ANOVA revealed that sites were a significant factor but gender was not. LPO increased significantly at TAD (circa 2-fold increase) in the clams from the St. Lawrence estuary. In the Fjord du Saguenay, LPO was readily increased at most sites (PS, ASJ and AE), rising by a factor of 1.6 at the site furthest upstream (i.e., the AE site). Phagocytic activity was affected by site location, with no gender-related effects (two-way ANOVA: p < 0.01 for site and p = 0.18 for gender) (Fig. 3C). In the St. Lawrence estuary, phagocytic activity was reduced at the PAL site, but it increased significantly at the TAD and BAR sites. In the Fjord du Saguenay, phagocytic activity was reduced at all sites (i.e. the PS, ASJ, BE and AE sites). Correlation analyses revealed that GST activity was significantly correlated with the weight-to-shell-length ratio (r = −0.52; p = 0.001) and GSI (r = −0.26; p = 0.01), suggesting that xenobiotic conjugating activity (pollution effects) occurs at the expense of clam condition and gonad development. LPO in gonads was correlated with the ratio of weight-to-shell-length (r = 0.29; p = 0.01), GSI (r = 0.69; p < 0.001), GST activity (r = –.73; p = 0.01) and phagocytosis (r = −0.34; p = 0.01). Phagocytic activity was significantly correlated with weight-to-shell-length ratio (r = −0.2; p = 0.04) and GST activity (r = 0.65; p = 0.01), suggesting that enhanced phagocytosis is detrimental to the condition factor and that increased phagocytosis occurs at sites where pollution stress also occurs.
DNA integrity was examined in the gonadal tissues of clams by tracking changes in DNA strand breaks, DNA nitrosylation and the purine salvage pathway by xanthine oxidoreductase (XOR) activity (Figs. 4A to C). The levels of DNA strand breaks were significantly affected by site location but not by gender (two-way ANOVA p < 0.01 for site and p > 0.05 for gender) in both sectors (Fig. 4A). In the St. Lawrence estuary, DNA strand breaks were increased about 3-fold at the BSC and TAD sites. In the Fjord du Saguenay, the levels of DNA strand breaks were significantly reduced at the PS and AE sites, with no significant changes at the other sites. The levels of DNA nitrosylation were also affected by site location but gender had no significant effect (Fig. 4B). The extent of nitrosylation was readily elevated at the most impacted sites (BSC and TAD) in St. Lawrence estuary clams. In the Fjord du Saguenay, the levels of nitrosylation dropped at polluted sites, reaching a 2-fold reduction at the site furthest upstream (i.e. AE). DNA nitrosylation was marginally correlated with the weight-to-shell-length ratio (r = −0.24; p = 0.05), GSI (r = 0.84; p = 0.01), phagocytic activity (r = 0.32; p = 0.02) and DNA damage (r = 0.41; p = 0.004). In an attempt to highlight the contribution of DNA nitrosylation (resulting from inflammation) to DNA strand breaks, the residual values (absolute value) of DNA strand breaks were calculated to identify any site-specific changes in DNA strand breaks not explained by DNA nitrosylation (i.e. DNA damage not produced by inflammation). The extraction of residuals for DNA strand breaks against DNA nitrosylation (Fig. 3A) revealed that DNA strand breaks were reduced in the St. Lawrence estuary but that the breaks nevertheless increased in the Fjord du Saguenay, suggesting that pollution-related DNA damage was still occurring there. It is noteworthy that the residual DNA strand breaks dropped significantly at the reference site in the fjord (i.e. site ASE).
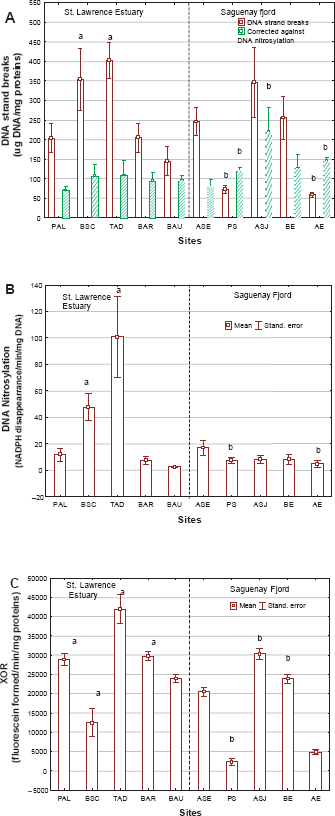
DNA lesions in feral
XOR activity of was significantly influenced by site location only, with no gender-related interactions (Fig. 3D). XOR activity was significantly induced at nearly all the St. Lawrence estuary sites, with the exception of the BSC site, where it was significantly reduced (Fig. 4C). In the Fjord du Saguenay, XOR activity was significantly induced at the ASJ site and reduced at the PS and AE sites. Correlation analyses revealed that XOR activity was negatively correlated with the weight-to-shell-length ratio (r = −0.3, p = 0.02), GSI (r = −0.51; p = 0.01), phagocytosis (r = 0.67; p < 0.001), DNA nitrosylation (r = 0.4, p = 0.007), GST (r = 0.93; p < 0.001) and DNA strand breaks (r = 0.42, p = 0.007). An analysis of covariance of XOR activity with site location, DNA strand breaks and DNA nitrosylation revealed that site location (F = 37) and DNA strand breaks (F = 3.3) were the most significant variables. Interestingly, GST was not significantly correlated with any of the above genotoxicity parameters, with the exception of XOR activity (r = 0.93; p < 0.001), but DNA strand breaks were negatively correlated with LPO (r = −0.5; p < 0.001), GSI (r = −0.68; p < 0.05) and positively correlated with phagocytic activity (r = 0.3; p < 0.01).
Discussion
Clams collected at sites affected by harbours and marinas show evidence of DNA damage in gonad tissues. A recent study correlated an increase in the formation of DNA strand breaks with changes in the energy expenditure associated with reduced clam-bed density profiles.
25
Harbours are known to be contaminated with fuel-based products such as PAHs, which are well-known DNA-damaging agents.
26
Exposure of
In a previous study of
In another study, total PAH levels were significantly correlated with levels of DNA adducts in blue mussels in Iceland. 28 PAHs were also recognized to increase GST activity in mussels, 29 which was correlated with XOR activity, a purine salvage pathway. In this study, DNA strand breaks were associated with DNA nitrosylation, which is considered an immune- or disease-mediated type of damage. Peroxinitrite released during phagocytosis reacts to DNA, which can lead to nitrosylation and deamination of cytidine residues. 30
At the St. Lawrence estuary sites, the levels of DNA strand breaks, when corrected against DNA nitrosylation (i.e. the residuals) were not significantly different from those at the BAU reference site, suggesting that DNA damage seems the result of an immunological response rather than through pollution effects in the gonad. The occurrence of red algal tides, which occurs in the estuary but not in the Fjord du Saguenay, could stimulate phagocytosis and lead to increased DNA damage. Clearly, more research will be needed to determine whether algal blooms can stimulate the immune systems in bivalves and contribute to DNA nitrosylation.
In the Fjord du Saguenay, DNA strand breaks at the ASE reference site were mostly affected by DNA nitrosylation. This suggests that clams at this site were under stress from disease or infection–-a finding corroborated by histological analyses of gonads that revealed that this site had the highest incidence of trematode (parasite) infestations (results not shown). This was a sporadic observation, but a systematic analysis of the infestation of clams was carried out in the present study. Conversely, the residual DNA strand breaks were higher at the AE site–-the site furthest upstream and known to be contaminated by multiple sources of pollution, indicating site-specific pollution effects.
During phagocytosis, microorganisms are killed by the concomitant production of nitric oxide and hydrogen peroxide, leading to the formation of the highly cytotoxic agent peroxinitrite during the so-called oxidative burst of immunocytes.
31
As mentioned earlier, NO production is also stimulated by the proinflammatory cytokine IL-2 in mussels. Clams and mussels exposed to β-glucans (cell wall components of various infectious microbes) also increased the production of nitric oxide in hemocytes.
32
Moreover, clam hemolymph treated with several doses of β-glucans inhibited the growth of 3 potentially pathogenic bacteria:
In another study, blue mussels exposed to untreated sewage effluent increased phagocytic activity, NO and hydrogen peroxide production after 14 days of exposure,
7
indicating that sustained production of these mediators of inflammation might be harmful to the host. This was corroborated by the significant relationship of phagocytosis activity with DNA nitrosylation, XOR activity and DNA strand breaks in the present study. The production of NO in hemocytes of
Sustained phagocytosis and the release of mediators of inflammation such as IL-2 could increase the concentrations of hydrogen peroxide and NO, which lead to the formation of peroxinitrite and reactions to genomic DNA. Indeed, NO leads to the formation of base nitration (nitrosylation) in nucleic acids (8-diazoguanine). The reduction of nitrosylated bases to amino bases underpins the measurement by NADPH-dependent diaphorase activity. The diazo bases and amino bases can form hydrogen bonds with each other and produce conformational changes in DNA, which can increase the propensity for DNA adducts and trigger a greater number of mutations. This would suggest that organisms battling infections are more susceptible to genotoxic compounds, since their DNA is already damaged by an increased number of nitrosyl adducts. This hypothesis was also consistent with the observation that age correlates negatively with DNA nitrosylation (r = −0.24; p = 0.05), owing perhaps to the weakened immune system and the generally less efficient DNA repair capacity that comes with age. In conclusion, DNA strand breaks seem to follow both pollution- and immune-mediated damage in the Fjord du Saguenay and the St. Lawrence estuary. DNA nitrosylation was significantly correlated with both the immune response (phagocytosis) and DNA damage suggesting that genotoxicity was aggravated by sustained inflammation and pollution.
Disclosure
The authors report no conflicts of interest.
Footnotes
Acknowledgments
This work was funded by the postdoctoral fellowship program of the Spanish ministry of Education and Science. The authors would like to thank Chantal André, Melanie Douville and Sophie Trépanier for their assistance with the experimental approach and biochemical determinations. We also thank Patricia Potvin and Heather Ferguson of Environment Canada for the English editing.