
Abstract
Pharmacological differences between neurogenic sympathetic responses in rat and pig isolated pulmonary arteries were examined in strip preparations. Electrical field stimulation in the range of 0.6 to 40 Hz resulted in frequency-dependent contractions in terms of amplitude and rate of rise. Responses in the rat declined sharply from pulmonary trunk to main artery; in contrast, in the pig they continued into the third-order vessels. Contractions were inhibited in the presence of tetrodotoxin, prazosin or WB-4101 and hence neurogenic in origin. Cocaine enhanced field stimulated contractions in both rat and porcine tissues; however, the effect in the former was of significantly greater magnitude in terms of either area under the mechanogram or height of contraction. In addition, the rate of rise, time to peak and duration of peak were all increased in the rat but less so or not in the pig. Field stimulated contractions were virtually abolished by guanethidine (1×10−6 M) in rat but not in porcine pulmonary arteries in which a ten-fold higher concentration significantly reduced neurogenic contractions and abolished them in 2 out of 4 tissues tested. The effect of guanethidine (1×10−6 M) observed in blood vessels of rat exceeded about five-fold that observed in porcine tissues. Thus, neurogenic responses appear to be entirely mediated by extra-junctional α1-adrenoceptors in both species, and in contrast to the rat, pig tissues seem to have a noradrenaline re-uptake that is either less efficient or operating near saturation.
INTRODUCTION
The sympathetic innervation of the pulmonary artery varies considerably among mammalian species.[1] However, the common picture would suggest that large vessels are richly endowed with adrenergic axon terminals while smaller vessels are sparsely endowed or not innervated. Adrenergic fibers are typically located approximately 5-10 μm from the medial smooth muscle layers.[2] In porcine pulmonary arteries, adrenergic terminals were identified in the tunica media, where they were seen to run around the vessel between elastic laminae and smooth muscle cells.[3] Histochemical data reported in rats are somewhat unclear. According to Zussman,[4] large pulmonary arteries of the rat contain a dense adrenergic innervation with the terminals extending into the media and sub endothelial layer. However, a study by El-Bermani[5] supports the view that a direct adrenergic innervation may be lacking in rat pulmonary artery smooth muscle, noradrenergic fibers having been noted to pass through the smooth muscle layer of the vessel without forming varicosities. MacLean and colleagues[6] have concluded that adrenergic fibers are absent in rat pulmonary trunk and main arteries proper and occur solely in association with their vasa vasorum, which are formed by branches of bronchial arteries. Costa and Majewski[7] reported an electrically evoked release of [3H]-noradrenaline from spiral strips of rat pulmonary artery.
There is a lack of data in the current literature on vasomotor responses to sympathetic nerve stimulation in either rat or porcine pulmonary arteries. Thus it appeared useful to demonstrate and compare such neurally mediated responses in these two species, particularly in light of conflicting neurohistochemical information in the rodent. As shown in this investigation, electrical field stimulation (EFS) of pulmonary arterial strips produces neurogenic contractions in both species that have distinct pharmacological properties.
MATERIALS AND METHODS
Tissue preparations
All procedures on animals were carried out in accordance with the guidelines and approval of the Animal Care Committee, Memorial University, St. John's, Newfoundland and Labrador, Canada.
Male Sprague-Dawley rats (280-310 g) were anaesthetized with sodium pentobarbital (65 mg/kg i.p.). The pulmonary trunk and main (left and right) pulmonary arteries were isolated, dissected free of connective tissue at room temperature in Krebs buffer with the following composition (in μM): NaCl, 130; KCl, 4.0; glucose, 11; MgCl2, 1.2; CaCl2, 2.5; KH2PO4, 1.2; NaHCO3, 12.5; EDTA, 0.1. The pH of the buffer following saturation with a 95% O2: 5% CO2 gas mixture was 7.4 at 36±1°C.
Pigs of either sex (2-9 months of age) were sedated with ketamine (22 mg/kg i.m.) and xylazine (2.0 mg/kg i.m.), before being intubated and maintained on isoflurane (1.5%-1.0% in 100% O2). Specimens of pulmonary arterial trunk and of juxta-hilar lung plus main bronchi were excised from animals immediately following thoracotomy and removal of the heart. Segments of trunk, main pulmonary artery and intralobar pulmonary arteries were dissected free of connective tissue at room temperature in Krebs buffer under the same condition as above.
Experimental protocol
After isolation from adjoining structures and removal of surrounding adipose tissues, the rat pulmonary trunk and arteries were divided at the bifurcation into two corresponding segments, before being cut into helical strips approximately 7 to 10 mm in length and 1.5 to 2 mm in width. Vascular preparations made from porcine pulmonary arteries included: transverse strips of the extrapulmonary portion of the main right pulmonary artery and its second order branches and helical strips dissected from third order intrapulmonary branches (approximately 10 to 12 mm in length and 2 to 3 mm in width). Tissues were mounted on a holder armed with two platinum ring electrodes spaced 10 mm apart in organ baths (Radnoti LLC, Calif., USA) at 36-37°C. Preload tension levels employed were as follows: for rats, 0.3 g (trunk) or 0. 15 g (right pulmonary arterial strips), and for pigs, 1.0 g. Bath media were gassed continuously with a mixture of 95% O2: 5% CO2. Contractile tone was recorded by force displacement transducers (Model FT03, Grass Instruments Co., Mass., USA) connected to a polygraph (Model 7PCPB, Grass Instruments Co., Mass., USA). EFS was delivered by a Grass S88 stimulator generating trains of rectangular monophasic pulses of selected frequency (0.6 to 40 Hz), width (0.5, 1.0 or 2.0 ms) and amplitude (50 V) applied in repetitive train mode (10 s train duration; 100 s train interval) or intermittent mode with 100 or 200 pulses per train delivered at intervals of 4-5 minutes. Ascorbic acid (10−4 M) was routinely added to the bathing medium to prevent the electrochemical oxidation of noradrenaline.[8] Cocaine HCl (1.0 μM), guanethidine monosulphate (1.0 and 10 μM), tetrodotoxin (1.0 × 10−7 M), the non-selective α1-adrenoceptor antagonist, prazosin HCl (1.0 and 3.0 × 10−7 M) and, the α1A/D-adrenoceptor antagonist, WB-4101 (1.0 × 10−9 M to 3.0 × 10−8 M)[9] were added in aqueous solution to the bath fluid after the preparations had reached a reproducible and consistent magnitude of evoked contractions.
Data and statistical analysis
Traces were digitized and responses to EFS assessed by measuring contraction height and the “area under the mechanogram trace” of the evoked response. The data were normalized as percent of the respective control response in the absence of test drugs. Comparisons employed Student's t-test, analysis of variance with repeated measure and the Bonferroni test for multiple comparisons. The data are based on replications from at least 3-5 individual animals.
RESULTS
EFS consistently elicited contractions in strips obtained from rat pulmonary trunk but produced no or barely detectable (<2 mg) responses in strips of rat main pulmonary artery. Contractile tension began to rise within 10 seconds of EFS and continued to climb for another 15 to 20 seconds after its off-set before falling to baseline over the next 60 to 100 seconds (Fig. 1). Although some pulmonary trunk preparations (33%) responded at a pulse width of 0.5 ms, the majority required a pulse width of 1. ms to generate a contraction >20 mg in amplitude. In contrast, irrespective of their anatomical origin or size, transverse or helical strips from different segments of porcine pulmonary arteries invariably contracted at a pulse width of 0.5 ms. Evoked contractions were inhibited in blood vessels from rat and pig in the presence of tetrodotoxin (Fig. 1), or prazosin or WB-4101 (Fig. 2) and hence considered neurogenic in origin. The antagonism displayed by WB-4101 was equally effective at 2.5 Hz (not illustrated).
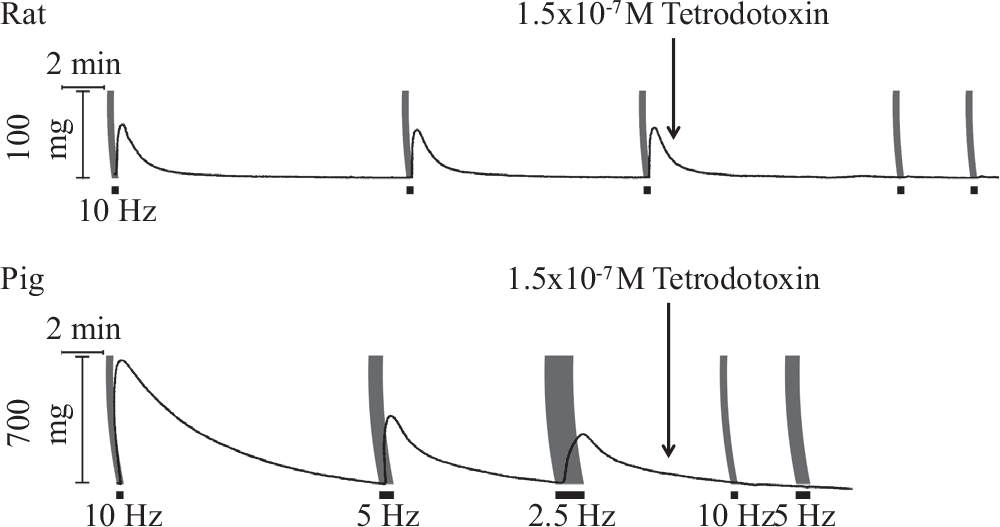
Effect of tetrodotoxin on contraction elicited by 200 pulse train electrical field stimulation (black bars) in rat pulmonary trunk (width 1.0 ms) and pig main pulmonary artery (width 0.5 ms) at frequencies indicated.
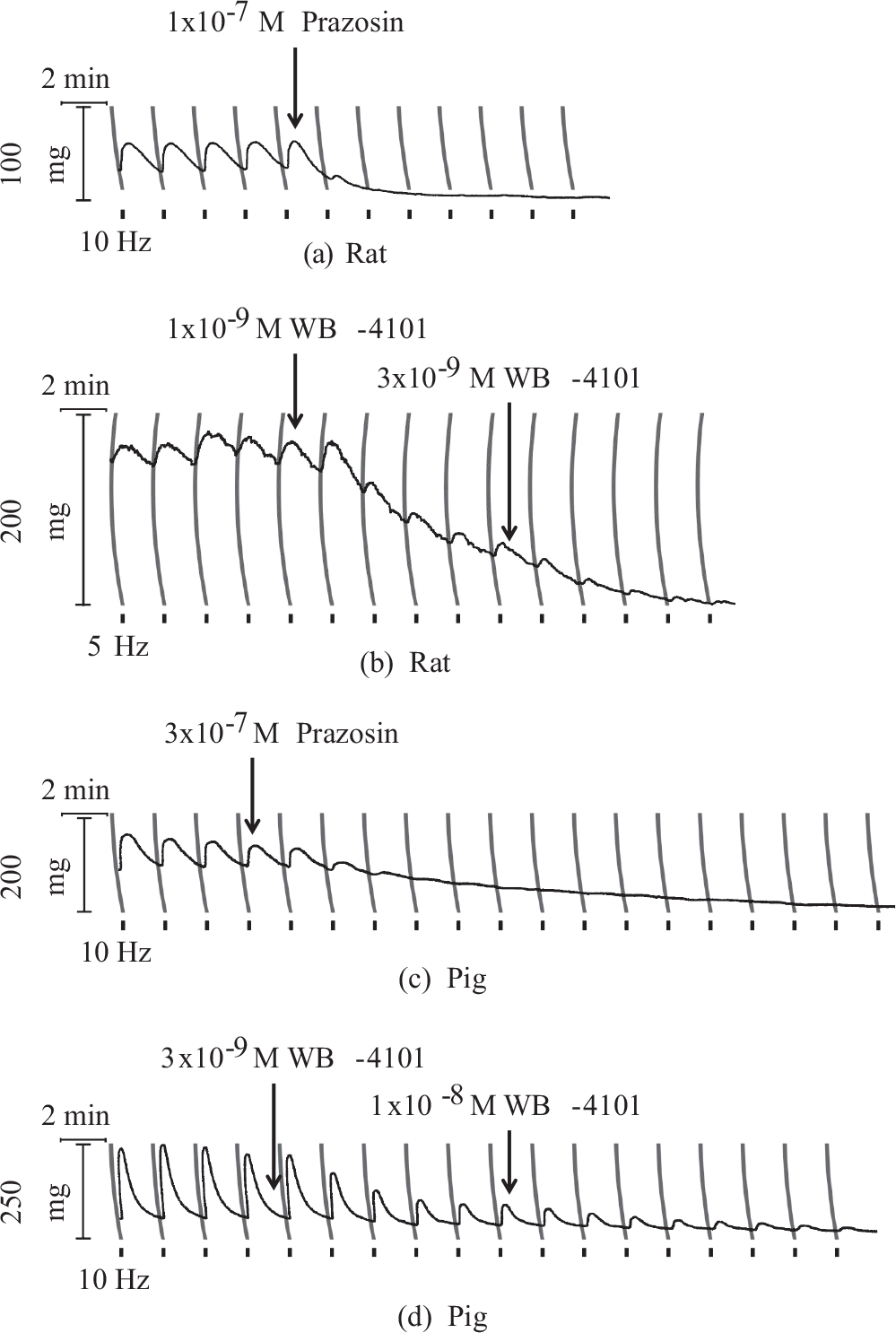
Effect of α1-adrenoceptor antagonists (prazosin and WB-4101) on field stimulated contraction induced in rat pulmonary trunk (pulse width 1.0 ms) and in pig second order pulmonary artery branch (pulse width 0.5 ms) by 100 pulse train electrical field stimulation (black bars).
With increasing pulse frequency in the range of 0.6-40 Hz, EFS resulted in contractions of increasing height and rate of rise with the threshold being apparent at 0.6 Hz and peaks being reached below and above 5.0 Hz for rat and pig tissues, respectively (Fig. 3). In the former, maximal neurogenic contractions elicited at 10 Hz amounted to 12-13% of the maximal tension induced by noradrenaline (1.0 to 10 [J.M). In the pig, the corresponding value showed proximo-distal variation with a range from 40% (main pulmonary artery and first order vessels) to 10-20% (second to third order vessels). The presence of cocaine enhanced field stimulated contractions in both rat and porcine tissues (Fig. 3); however, the effect in the former was of significantly greater magnitude in terms of either area under the mechanogram (Fig. 4) or height of contraction (not shown). In addition, the rate of rise, time to peak and duration of peak were all increased in the rat but less so or not in the pig (Fig. 3). The cocaine-induced enhancement of neurogenic contractions in rat or pig tissues did not vary significantly with increasing EFS pulse frequency in the range tested. The presence of cocaine significantly decelerated relaxation. Thus in rat tissues at frequencies of 2.5 and 5.0 Hz, half maximal relaxation time increased by 83±6% and 56 ± 16% and in the pig it increased by 59 ± 15% and 28±6% (mean ± s.e.m), respectively.
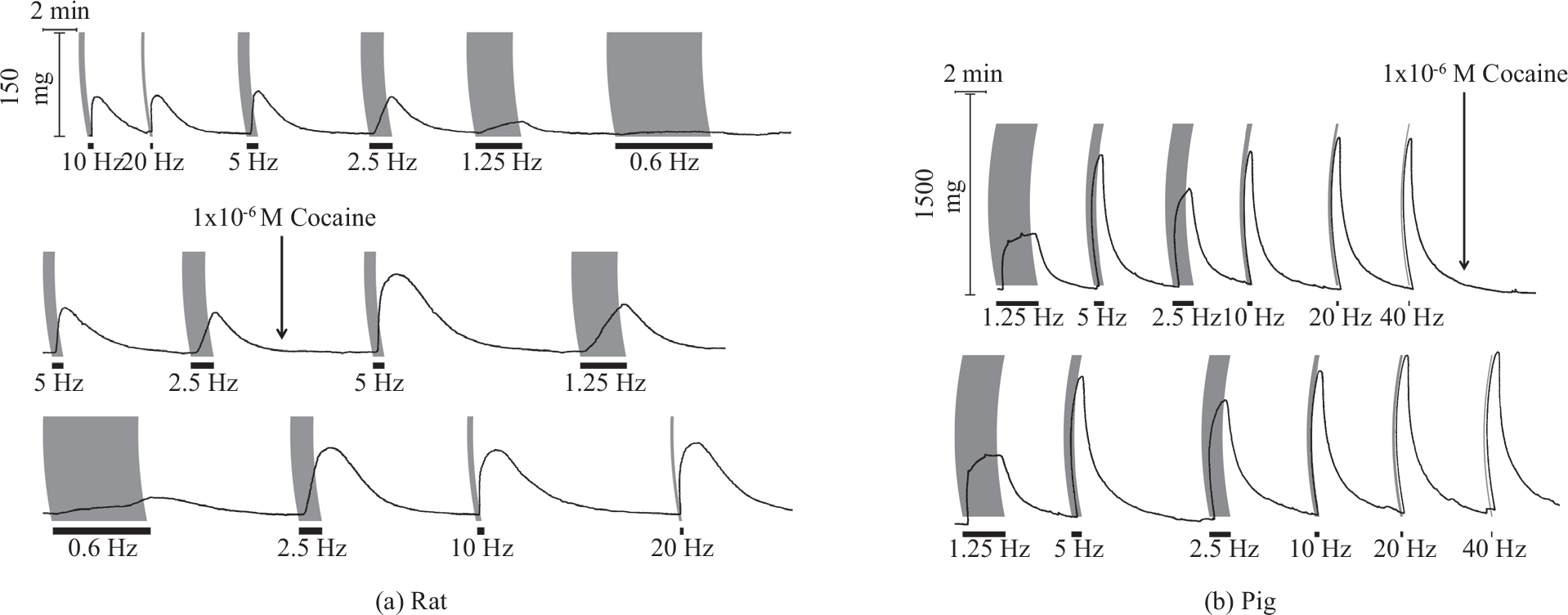
Frequency dependence of neurogenic contractions elicited by 200 pulse train field stimulation (pulse width 1.0 ms; black bars) in the absence and (a) presence of cocaine in the rat pulmonary trunk and (b) pig second order pulmonary artery branch.
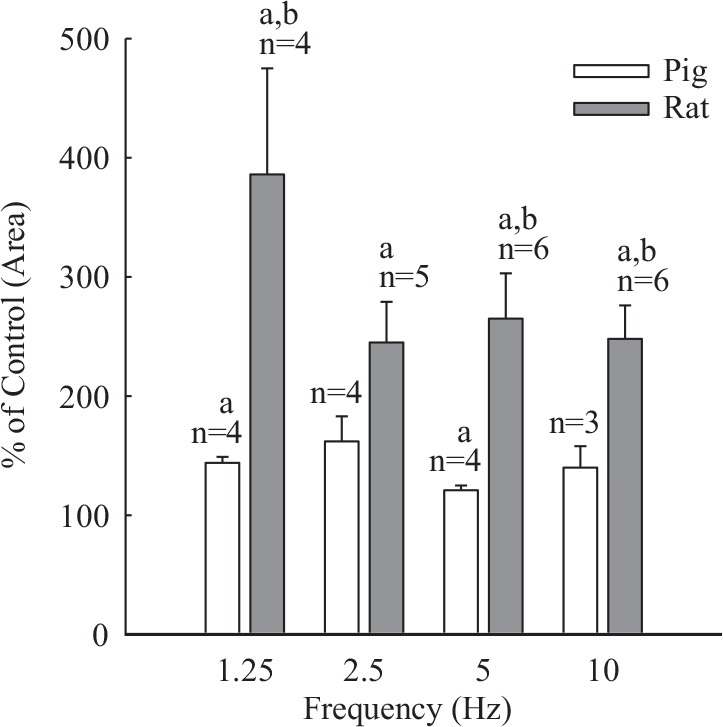
Effect of cocaine (1.0 × 10−6 M) on neurogenic contraction of rat and pig pulmonary arterial strips. Shown are histograms of area under mechanogram traces. Responses were evoked by trains consisting of 200 rectangular pulses (width 1.0 ms, rat; 0.5 ms, pig). aSignificantly different from control P<0.05; bSignificantly different from respective response in pig P<0.05.
In the presence of guanethidine (1× 10−6 M) field stimulated contractions were virtually abolished in rat but were unaffected in porcine pulmonary arteries (Fig. 5). In the latter a higher concentration of guanethidine (1×10−5 M) significantly reduced neurogenic contractions and abolished them in 2 out of 4 tissues tested (Fig. 5). The effect of guanethidine (1×10−6 M) observed in rat tissues exceeded by about five-fold that observed in porcine tissues (Fig. 6).
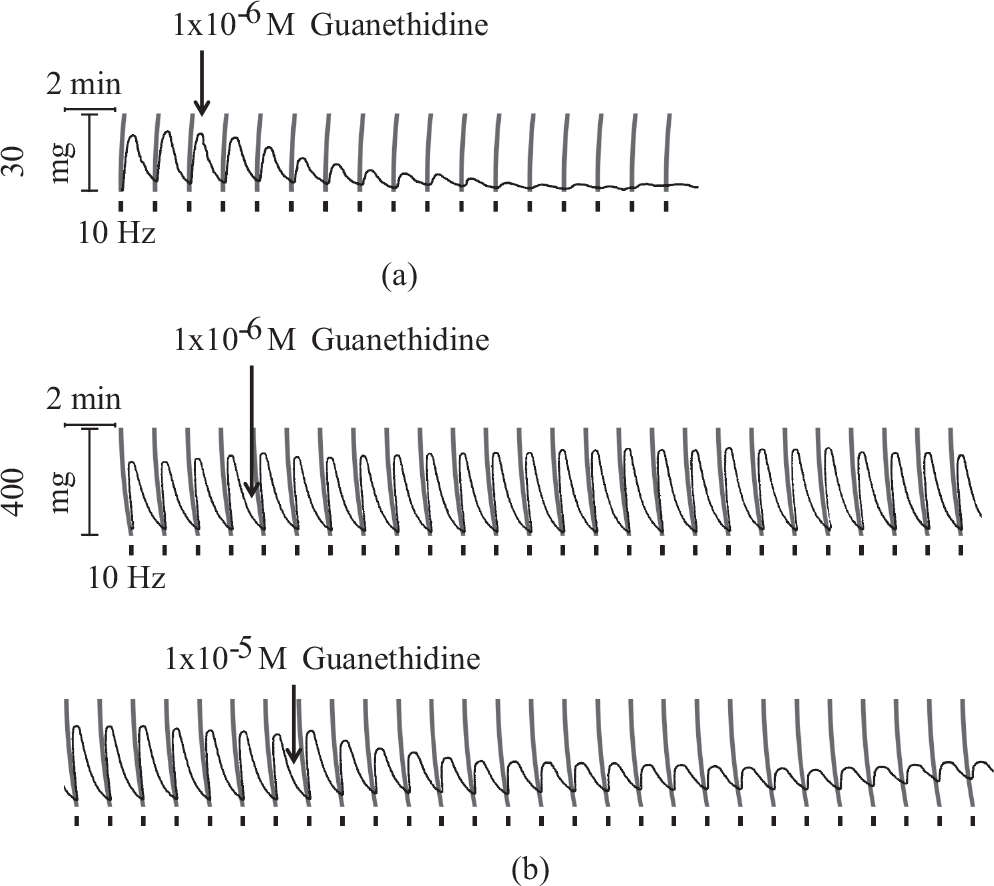
Differential effect of noradrenergic neuron blocker, guanethidine, on neurogenic contraction evoked in rat pulmonary trunk (pulse width 1.0 ms) (a) and pig second order pulmonary artery branch (pulse width 0.5 ms) (b) by 100 pulse train electrical field stimulation (black bars).
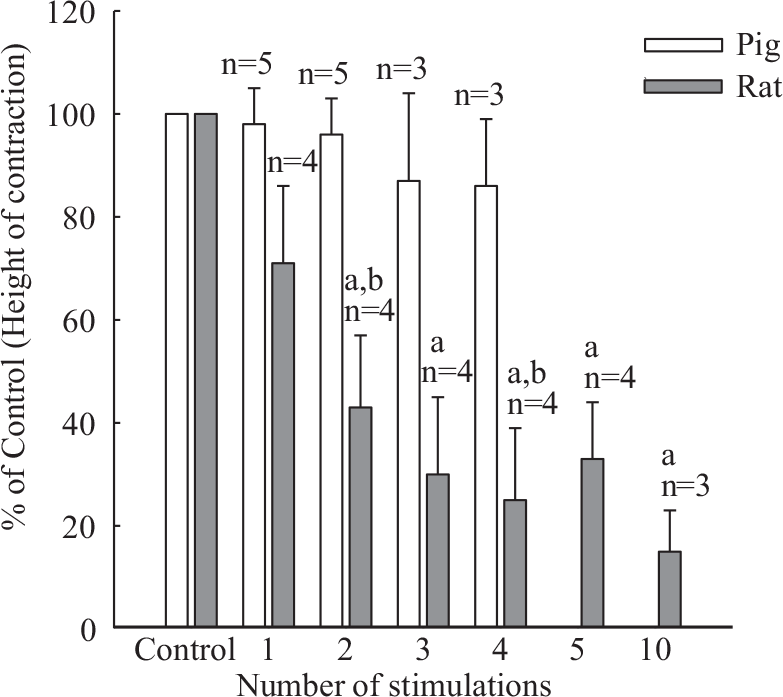
Progressive inhibitory effect of guanethidine (1.0×10−6 M) on height of contraction evoked by 100 pulse trains in rat pulmonary trunk (pulse width 1.0 ms), and lack of inhibition in pig second order pulmonary artery branch (pulse width 0.5 ms). aSignificantly different from control P<0.05; bSignificantly different from respective response in pig P<0.05.
DISCUSSION
The data presented here demonstrate that the pulmonary arteries of the rat and the pig possess a functional adrenergic vasoconstrictor innervation. Despite basic similarities, including inhibition by tetrodotoxin and α1-adrenoceptor antagonists, the observed neurogenic vasoconstrictor responses were shown to exhibit species dependent differences in (a) anatomical distribution and (b) pharmacological characteristics.
Distribution of EFS-evoked responses
In agreement with available histochemical evidence,[3] the porcine pulmonary arterial strips displayed neurogenic contractions irrespective of the vessel caliber tested; however, in the rat the response appeared to be virtually confined to the pulmonary trunk. If adrenergic terminals supplying the vasa vasorum of rat pulmonary arteries were to constitute the sole source of vasomotor innervation at this level, their involvement in neurogenic contractions would be plausible. Yet the difficulty to detect this response in the main pulmonary artery (i.e., past the bifurcation of the pulmonary trunk) would militate against this idea. In any case our observations are consistent with the presence in the rat pulmonary arterial trunk of a functional sympathetic innervation.
The density of adrenergic innervation in arteries of various species has been correlated with the efficacy of EFS in producing vasoconstrictor responses, i.e. blood vessels with low-density innervation have a shallow frequency response relationship and low relative efficacy expressed against agonist-evoked maximal contractions.[10] In this regard, the rat pulmonary trunk and porcine second to third order strips behaved like sparsely innervated tissues. It is, however, worth noting that a larger pulse width was required to evoke a response in rat as opposed to pig pulmonary arterial strips. The greater electrical excitability of porcine pulmonary constrictor nerve terminals would be consistent with a higher innervation density or transmission efficiency in this species. The possibility of the EFS-evoked responses being due to direct activation of muscle fibers can be dismissed in view of the observed full blockade obtained with tetrodotoxin in both rat and pig tissues.
The density of the vasoconstrictor nerve terminals is known to vary in the same vascular bed. For example in the rat mesenteric artery, the sympathetic innervation, as assessed by histochemical and functional criteria, gradually decreases from the principal arteries to the terminal arterioles, with the precapillary arterioles apparently being devoid of innervation.[11,12] Based on the present results, the density of adrenergic terminals in the rat pulmonary artery would seem to decrease sharply beyond the bifurcation of the trunk.
Pharmacological characteristics
Since the sympathetic neurogenic responses were abolished by the α1-adrenoceptor antagonists, prazosin and WB-4101, co-transmission by ATP appears to be absent or functionally insignificant in the pulmonary arteries of rat and pig. ATP is considered to play no role as a co-transmitter in vasomotor responses mediated via the activation of sympathetic nerves in veins and pulmonary arteries.[13] In addition, no evidence was obtained for a participation of α2 adrenoceptors at low EFS pulse rate. The latter has been demonstrated by Bao et al.[14] in the rat tail artery.
As revealed by the effect of guanethidine and cocaine, there were striking pharmacological divergences in rat and pig pulmonary arterial strips. These could be due to different transmitter kinetics i.e. release and uptake, more specifically, the regulation of the clearance of noradrenaline released neurally. Evidently, the pig pulmonary artery was relatively less sensitive to the pharmacological actions of either guanethidine or cocaine. The inhibitory actions of guanethidine are critically dependent on its uptake via the axolemmal amine transporter into the nerve endings while the potentiating actions of cocaine on neurogenic transmission are largely attributed to inhibition of neuronal uptake. Under conditions where neuronal uptake is inefficiently used or saturated, the pharmacological actions of both guanethidine and cocaine would become compromised. The latter possibility may thus apply to adrenergic neurogenic vasoconstriction in the pulmonary artery of the pig.
Neuronal uptake of noradrenaline plays an important role in controlling the extent of the functional response as well as the duration of action in mammalian tissues, and this transporter has also been cloned.[15] It is also recognized that the role of neuronal uptake for noradrenaline-induced neurogenic responses in vascular smooth muscle remains incompletely defined. Thus, in the mesenteric arteries, the density of sympathetic nerve innervation has been cited as the parameter controlling the effects of cocaine on noradrenaline- and neurally-mediated vasoconstriction.[12] In the rat caudal artery, a low concentration of noradrenaline (60 nM) exogenously applied or neurally released has been shown to be inactivated and removed by neuronal uptake.[16] However, according to Webb and colleagues[17] in the same tissue, responses to low (0.1-1.0 Hz) but not high (2.0-16 Hz) frequency stimulation were enhanced by cocaine (1.0 μM), at variance with the present findings and those of Stjärne et al.[18] Nonetheless, the apparent half time of relaxation was found not to be different at 1.0 versus 16.0 Hz in the presence of cocaine.[17] The present data obtained in both rat and pig pulmonary arterial strips are in agreement with the literature in showing that cocaine retarded rate of relaxation irrespective of impulse frequency.
A two-compartment working model was proposed for the removal of neurally released noradrenaline.[18] In that model neuronal uptake near active neuromyal junctions saturates very quickly. In addition, released noradrenaline diffuses into a compartment “surround” which rarely saturates irrespective of nerve activity. Associated with the latter compartment is a selective binding substrate “S” acting as a reservoir for sequestration of noradrenaline that can later be released once neural exocytosis has ceased and hence maintains biological function.[18] Our results obtained in pig tissues are at least partly compatible with the two-compartment model.
A quantitative description of the mechanical function in isolated blood vessels has been put forward in line with the experimental observations that the relatively slow decline of response after cessation of sympathetic nerve stimulation is mainly due to the slow rate of removal of noradrenaline from the extracellular space by neuronal reuptake and diffusion into surrounding areas.[19] Under such conditions, the effectiveness of both cocaine and guanethidine could be impaired as seems to be the case with our observations in the porcine pulmonary artery, in striking contrast to the rat pulmonary neurogenic vasoconstriction.
Perspective
Sympathetic nerve stimulation in vivo increases vascular resistance in the pulmonary arterial vasculature. Pace and colleagues[20] reported that in dogs, an increase in impedance occurs in the pulmonary arterial bed as a consequence of sympathetic nerve activation. The elevation in impedance in the pulmonary arterial bed is accompanied by an increase in oscillatory component in input to hydraulic power, and this occurs independently of either positive chronotropic or inotropic effects arising from the right ventricle. The physiological basis for this event is a stiffening of the main pulmonary artery causing augmentation of pulse pressure for the same volume of flow generated by the right ventricular contraction and a more efficient delivery of blood to the lungs.[20] Similar effects have also been reported to occur in cats due to injections of noradrenaline or sympathetic nerve stimulation.[21] Thus, it is proposed that in a state of low cardiac output, α-adrenergic activation in the pulmonary arterial bed increases the fraction of hydraulic power delivered by the right ventricle, which then allows for more efficient propulsion of blood into the pulmonary vascular bed downstream.[21] Certainly, this concept is supported by denser adrenergic innervation in the larger vessels as noted in our current investigation. Regioselective innervation of the pulmonary arterial vessels may permit a more efficient delivery of blood to the lungs with lower work load for the right ventricle: for instance, during low output or intense physical activity sympathetic neuron driven proximal pulmonary vasoconstriction could play an essential role.