
Abstract
Microparticles are submicron vesicles shed from a variety of cells. Peter Wolf first identified microparticles in the midst of ongoing blood coagulation research in 1967 as a product of platelets. He termed them platelet dust. Although initially thought to be useless cellular trash, decades of research focused on the tiny vesicles have defined their roles as participators in coagulation, cellular signaling, vascular injury, and homeostasis. The purpose of this review is to highlight the science leading up to the discovery of microparticles, feature discoveries made by key contributors to the field of microparticle research, and discuss their positive and negative impact on the pulmonary circulation.
Over the past decade, microparticles have rapidly made their way into the headlines of journal articles and seminars concerning all areas of medical research. Microparticles are submicron vesicles shed from various cell types and released into the extracellular environment.[1] Increases in total numbers of microparticles are associated with disease states such as atherosclerosis, diabetes, cancer, sepsis, and pulmonary hypertension.[2] Yet, we know there are constitutively circulating microparticles that contribute to overall vascular health and homeostasis in coagulation.[3]
The focus of the emerging field of microparticle research is centered upon characterizing the phenotypes of these novel biopackages, investigating the mechanisms of their formation from parental cells, and determining their effects on vascular cells particularly in pathological conditions.[4] While many questions remain unanswered, numerous findings within the past 40 plus years have allowed the field of microparticles to progress dramatically from the original discovery of “platelet dust” by Peter Wolf in 1967. We now appreciate that microparticles are not simply inert products of cellular debris. These tiny vesicles take center stage in literature concerning numerous diseases, but microparticles of healthy origins also play an essential role in intercellular communication and coagulation events that contribute to the overall homeostasis of the body.
As the significance of these signaling packages emerges, it is important to remember their humble beginnings. Platelet microparticles are the most widely studied and the most abundant microparticles found in the normal, healthy circulation.[5–7] Thus, our review begins with a focus on the initial questions left unanswered by the description of coagulation. The contributions of platelet microparticles to coagulation will be described. To date, the cell types reported to release microparticles either constitutively or when stimulated include platelets, blood cells, endothelium, epithelium, and various cancer cells, and the list continues to grow as does our understanding of microparticle constituents and signaling capacities.[1,8,9] We will also highlight some of the more recent findings that suggest important roles for microparticles in both the homeostasis of the pulmonary circulation and pulmonary pathologies.
HISTORICAL HIGHLIGHTS OF BLOOD COAGULATION
The discovery of microparticles can be traced back to initial research conducted on blood coagulation. Major findings in blood coagulation research spanning from the late 1600s to the mid-1900s are broken up into three eras-The preclassical theory era (1666–1904), the classical theory era (1904–1934), and the prothrombin era (1934–1943). In 1666 the Italian physician, Marcello Malpighi noticed that fiber filaments remained in a blood clot after it was thoroughly washed. Malpighi's observation sparked curiosity concerning the mechanisms of blood clotting.[10] Some 100 years later with his development of a novel method to prevent blood coagulation ex vivo, William Hewson[11] resolved that the coagulation factors of blood were present extracellularly in what he termed “coagulable lymph.” In 1875, Zahn reported that an injured blood vessel was eventually plugged by a fibrin-associated white thrombus. This observation led to the discovery that platelets are responsible for contributing fibrin in the blood coagulation process.[12] By 1904, collaborative efforts of scientists led to the establishment of the classical theory, which describes the conversion of prothrombin to thrombin and the subsequent production of fibrin.[10]
The classical theory era saw the emergence of conflicting theories about blood coagulation. This led to a halt in forward progress during the times between 1904 and 1934. Thus, the discoveries of the preclassical theory era were carried over into the prothrombin era.[10] In 1935, Dam was investigating chicks that developed an excessive bleeding disorder in response to synthetic diets. He found that bleeding did not occur if their synthetic chow was replaced with one fortified with a specific vitamin. Dam labeled this antihemorrhagic agent “vitamin K” and established its essential role in normal blood coagulation.[13] One of the great developments in the field of blood coagulation during this third era was a procedure to monitor prothrombin activity within blood. Prothrombin activity assays enabled physicians to characterize patients' bleeding disorders on the basis of clotting time and generate an appropriate therapy for each condition.[10] The emergence of this assay led to significant findings beyond the scope of the three historical eras concerning patients with bleeding disorders, such as hemophilia.
In 1962 at an International Committee on Blood Clotting Factors conference, scientists Robert Gwyn Macfarlane of Oxford and Oscar Ratnoff of Cleveland came to a consensus. They proposed that the coagulation process involved an enzyme acting on its substrate to make it an active enzyme, which subsequently acted on its own distinct substrate. This type of enzymatic activation continued down a line of substrates before coagulation was achieved. The two scientists published their findings independently after departing the meeting in Stockholm.[14] In 1964, MacFarlane's “coagulation cascade” was published in
One of the numerous contributors to the eventual identification of the coagulation cascade was British physician Peter Wolf. Wolf spent over a decade analyzing various events of blood coagulation. The articles he published between the years of 1953 and 1965 focused predominantly, but not exclusively, on an antihemophilic factor.[18] While coagulation factors were a hot topic due to interest in sorting out the individual components of the coagulation cascade, scientists were still intrigued by the broad picture of blood coagulation. Taking a few steps back to 1946, Erwin Chargaff and Randolph West discovered that platelet-free plasma exhibited clotting properties. This contradicted the major dogma of preceding times, which was that platelets served necessarily as the blood coagulation contributor. Chargaff and West also found that after a high-speed centrifugation of 31,000 ×
THE IDENTIFICATION OF “PLATELET DUST”
Wolf's clarification came in the form of his 1967 article, “The Nature and Significance of Platelet Products in Human Plasma”, published in the
“Platelet dust” was the term Peter Wolf coined to describe the subcellular coagulant material he observed. This material was reported to be present in both plasma and serum, rich in phospholipids, and originating from platelets. Relative to the accounts of Chargaff and West, “platelet dust” was sedimented by ultracentrifugation. Wolf also found that during the storage of platelet-rich citrated plasma, there was an overall increase of sedimentable “platelet dust.” Therefore, he expanded Hougie's theory of “platelet coagulation activity” of stored plasma and instead attributed this phenomenon to the coagulation activity of “platelet dust.” Furthermore, Peter Wolf concluded that “platelet dust” obtained from serum and citrated plasma possessed coagulation properties identical to Platelet Factor 3, a coagulation factor derived from platelets responsible for converting prothrombin into its active form, thrombin. Wolf felt that this explained the source of O'Brien's “platelet-like activity of serum”.[18] In conclusion, in his 1967 publication, Peter Wolf revolutionized previous blood coagulation literature with his discovery of “platelet dust”, which was subsequently replaced by the term, microparticles.
THE EVOLUTION OF MICROPARTICLE ROLES IN COAGULATION
Subsequently, but not based on Wolf's findings, other scientists captured additional electron microscopic images of subcellular vesicles released from circulating platelets. In an article published by Webber and Johnson in 1970, platelets activated by thrombin were observed releasing small vesicles into the blood plasma. Webber and Johnson[21] found that platelet alpha granule contents are encompassed into vesicles, which come together to form a membrane complex at the surface of the platelet. Finally, the group of vesicles is released into the circulation where their components are active in blood coagulation events.
In 1971, Peter Wolf's microparticle work was furthered by the contribution of one of his colleagues at the University of Birmingham in England. Using blood from humans and animals, Crawford studied platelet-derived microparticles to enhance the field's knowledge of microparticle appearance and composition. Crawford described electron micrographs of microparticles obtained from fresh pig platelet-free plasma as possessing “a heterogeneous granular appearance.” He also found that microparticles possess adenosine triphosphate (ATP) ATP-ase activity similar to platelet contractile proteins, like thrombosthenin, which he believed supported the theory that plasma microparticles originate from platelets and contain actomyosin-like contractile proteins. In addition, Crawford discovered that his microparticle samples contain lipids and contribute to the “free serotonin fraction” of plasma with their capabilities of harboring the neurotransmitter. In his discussion, Crawford[22] addressed the findings of other scientists of the time, such as Webber and Johnson, regarding the multivesicular membranous sacs of platelet pseudopodia. He stated, “the contents of these sacs or vacuoles closely resemble in structure the microparticles isolated from platelet-free plasma, being of a fine granular nature and of approximately the same size”. James George and others utilized a new method to characterize microparticles from normal human plasma and serum in 1981. They were able to obtain platelet-derived microparticle counts using an immunoelectrophoretic assay with an antibody against the platelet membrane glycoprotein complex IIb/IIIa. Platelet microparticles were identified in all samples, and concentrations were 10 times higher in serum compared to plasma. The authors also noted that a portion of the ultracentrifuged pellet did not associate with the plasma membrane antibody,[23] which was an early indication that platelets were not the only origin of circulating microparticles.
Over the past few decades, microparticles from different cell types have been identified as a source of blood-borne or circulating tissue factor. Tissue factor is a transmembrane protein capable of forming a complex with factor VII/VIIa, ultimately leading to thrombin generation. Tissue factor is not typically present in significant amounts in healthy circulations.[24] After observations that tumor cells release procoagulant vesicles,[25] a 1983 publication reported vesicles obtained from tumor cells by ultracentrifugation exhibit tissue factor activity.[26] Approximately, half the population of microparticles generated in vitro from both breast and pancreatic cancer cell lines express tissue factor, according to a 2008 study. The generated microparticles possess procoagulant activity in vitro and when injected into an animal model. These effects are abolished by the addition of a tissue factor antibody, indicating that the procoagulant activity of microparticles released by breast and pancreatic cell lines is tissue factor-dependent.[9]
In 1994, while investigating the effects of lipopolysaccharide (LPS) stimulation on monocyte procoagulant activity, it was revealed that the microparticles released by stimulated human monocytes possess more tissue factor activity than their parent monocytes.[27] A subsequent 2003 article reported the supernatant of mononuclear cells stimulated with LPS contained increased particle counts and greater tissue factor activity relative to the cell supernatant obtained from unstimulated monocytes. Ultracentrifugation of the supernatants removed the tissue factor activity, suggesting the presence of tissue factor on microparticles released from the monocytes.[28] Scientists then examined the effects of treating monocytes with microparticles produced by platelets in response to shear stress. Results indicated that the monocytes were activated, which led to tissue factor expression by the monocytes, themselves, and by the microparticles they generated.[29] Tissue factor-bearing microparticles produced by monocytes, platelets, smooth muscle, and endothelial cells have been identified in numerous pathologies, such as cancer, atherosclerosis, type II diabetes, sepsis, acute respiratory distress syndrome (ARDS), and pulmonary hypertension.[8,30–34] Circulating microparticles obtained from healthy individuals also express tissue factor.[35] Therefore, microparticles are potential mediators of blood coagulation by serving as circulating sources of tissue factor.
Furthermore, microparticles generated in response to certain stimuli can participate in coagulation by harboring phosphatidylserine on their surfaces. Phosphatidylserine is a negatively charged aminophospholipid that is localized on the cytosolic side of the plasma membrane under normal conditions. In response to stimulation or during apoptosis, membrane symmetry can be lost, resulting in exposure of phosphatidylserine on the extracellular surface.[36] The negatively charged aminophospholipid can then bind and assemble circulating coagulation factors. These events lead to the production of thrombin from prothrombin.[37] Upon stimulation with a calcium ionophore or collagen in combination with thrombin, platelets and red blood cells release vesicles possessing the ability to generate thrombin. The authors speculated this thrombin production was due to the presence of phosphatidylserine on the surface of the shed vesicles.[38] Years after this observation, fluorescent labeled annexin V binding detected by flow cytometry was developed as a technique to confirm the presence of phosphatidylserine on the surface of microparticles.[39] A group of scientists using this technique in the early 1990s concluded that circulating microparticles obtained from patients with meningococcal sepsis exhibited a greater affinity for annexin V and, therefore, an increased surface expression of phosphatidylserine.[33] In addition to characterizing the tissue factor activity of microparticles from LPS-stimulated monocytes, the results of a 1994 study previously mentioned pointed out that monocyte stimulation also leads to an increased phosphatidylserine expression on the surface of microparticles.[27] Microparticles exhibiting phosphatidylserine, like those harboring tissue factor, can contribute to procoagulant events once released into the circulation.
CHARACTERIZING MICROPARTICLES IN THE MODERN ERA: DEFINITIONS AND PROTOCOLS
Over the past 4 decades, since Peter Wolf's original description, studies of microparticles have increased nearly exponentially. As such, many technical and characteristic questions have arisen as to the “definition” of a microparticle and the other classes of circulating intact particles such as exosomes, ectosomes, and microvesicles. The International Society on Thrombosis and Haemostasis vascular biology subcommittee developed a standard definition describing microparticles at their 2005 conference. Microparticles are 0.1–1 micrometer cell-derived vesicles that lack a nucleus or synthetic capacity, may contain cytoskeletal proteins, and expose some quantity of phosphatidylserine on their surfaces (Fig. 1).[40] Frequently in the literature, the term microvesicle is used synonymously with the term microparticle and currently, there is no distinct definition reported for this term. There seems to be no consensus on a membrane or intravesicular marker to separate microparticles as a whole from another type of small particle, exosomes. However, there are some membrane markers utilized to identify parent cell phenotype. Included in these membrane markers is a number of the cluster of differentiation proteins including CD31+/CD42−, CD62e, and CD105 for endothelial microparticles; CD45 for leukocyte microparticles; and CD42+/CD31− for platelet microparticles. Flow cytometry using latex beads to set size-specific gates is most often used, especially in identification of microparticles from blood samples where there are expected to be mixed populations.[41–43] However, many studies merely show bar graphs of accumulated data and not the actual flow cytometry plots, thus making reviewing, interpreting, and comparing some studies difficult.
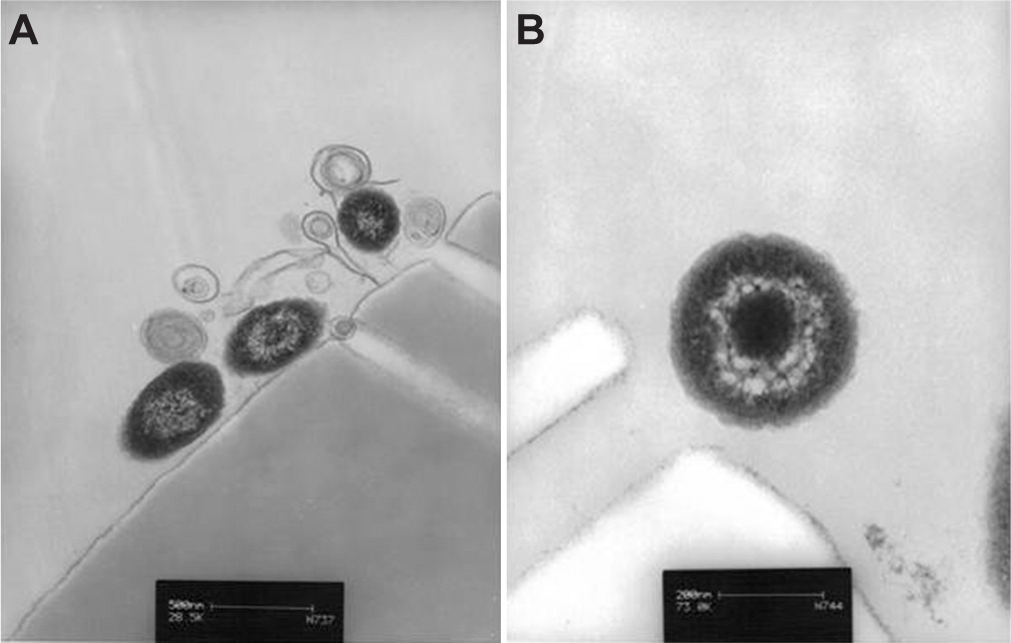
Microparticles isolated by ultracentrifugation and analyzed by electron microscopy.
Protocols used to collect microparticles from either cell culture medium or blood samples are as heterogeneous as the microparticles themselves. One common denominator among laboratories collecting the vesicles from blood is to begin with a centrifugation step to collect platelet-free plasma. This ensures that no further steps in the protocols induce activation and subsequent release of further microparticles from platelets. For analysis strictly in the biomarker sense, many investigators then proceed to a range of centrifugation speeds from 10,000 × g to 20,000 × g. The pellet from these centrifugations is then analyzed by flow cytometry.[6,44–46] However, others have gone on to include subsequent ultracentrifugation at 1,00,000 × g for microparticles used in studies to determine their contents and function.[8,47,48] There is currently no general consensus as to which protocols for microparticle isolation are the best. Some groups suggest that the ultracentrifugation in fact causes microparticles to congregate forming doublets and therefore giving false readings larger than the 1μm in diameter; however, others maintain that not performing the ultracentrifugation step leads to contaminated samples. With the formation of the International Society for Extracellular Vesicles, investigators are now beginning to work together to standardize the protocols that will drive microparticle research forward. Until that point in time, careful flow cytometry and electron microscopy analysis of the collected microparticle pellet is necessary.
Our first definition of an exosome comes from studies investigating maturation of reticulocytes. When describing a phenomenon that occurred during the maturation of reticulocytes into erythrocytes, Johnstone et al.,[49] coined the term exosome. During the maturation process, the transferrin receptor is lost from the plasma membrane and was found to be released in endocytic vesicles that fuse with the plasma membrane and are then excreted, thus called exosomes.[49] Currently, the size characteristic of exosomes varies slightly with some investigators reporting that they are 40–100 nm and others defining them as 0.5 μm and under. They are frequently isolated using sucrose gradients based on their defined buoyant density of 1.16–1.18 g/mL.[50] Exosomes have their own unique makeup that is defined by their extrusion pathway.[51,52] In a seminal study by Thery et al.,[53] proteomic analysis of exosomes isolated from apoptotic dendritic cells revealed distinct protein signatures including specific proteins from the endocytic compartment such as major histocompatibility complex II (MHC II) and Alix, a protein thought to regulate endosomal trafficking along with Rab7. To date, exosomes are frequently characterized by the presence of these marker proteins and collection by sucrose gradient centrifugation. However, in studies to isolate microparticles via high-speed ultracentrifugation it is plausible that exosomes remain in the pellet. Thus, many functional studies reported in today's literature may in fact be the actions of microparticles or a combination of both microparticles and exosomes.
Ectocytosis was originally used to describe a mechanism by which membrane-attack complexes are shed to protect a cell and was defined as such to differentiate from traditional exocytosis. The traditionally defined exocytic process requires fusion with the plasma membrane and emptying of the vesicle components into the extracellular milleu. Ectocytosis involves the shedding of right-side out membrane vesicles called ectosomes. Ectosomes were initially defined by the presence of C9, one of the pore forming members of the complement system.[54] This seminal work by Stein and Luzio[54] illustrated for the first time that there may be specific protein and lipid sorting mechanisms in the formation of a vesicle released from a cell. Since this study, ectosomes have been incorporated into the umbrella terms of microparticles and microvesicles based on isolation by ultracentrifugation and their size of approximately 1 mm in diameter. [55] The mechanisms of microparticle, exosome, and ectosome release are still active areas of investigation described below. Regardless of the preparation, whether made up of microparticles, exosomes, microvesicles, or ectosomes one thing is becoming increasingly evident: All of these circulating tiny vesicles have significant impact on both physiological and molecular signaling.
MICROPARTICLES: FORMATION AND CLEARANCE
Because the earliest identified roles of microparticles related to coagulation, some of the first investigations into their mechanisms of release targeted phosphatidylserine exposure. Many studies focused on platelets, and activation of platelets leads to increased intracellular calcium, which in turn regulates membrane flippase, floppase, and scramblase.[56] These proteins are responsible for maintaining membrane asymmetry. Dysregulation of membrane asymmetry proteins induces exposure of phosphatidylserine on the outer membrane of a cell, as mentioned previously. The increase in intracellular calcium also activates intracellular signaling molecules such as calpain, responsible for cleaving portions of the membrane cytoskeleton, and Rho kinase. Rho kinase activation alters the cytoskeleton resulting in cellular contraction and membrane blebbing. Thus, a phosphatidylserine positive microparticle is generated and released (Fig. 2).[57,58] Interestingly, inhibition of Rho kinase in stimulated endothelium prevents the release of caspase 3-containing microparticles and results in cell detachment and death.[59] So microparticle release may, in fact, be a prosurvival mechanism for some cell types.
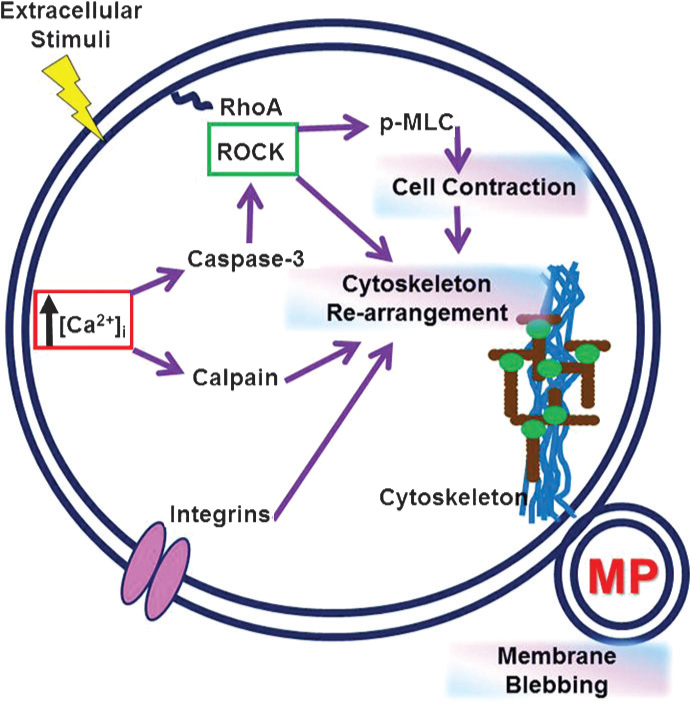
Mechanisms of microparticle generation. The mechanisms of microparticle generation are multifaceted. Many of the current identified mechanisms are calcium-dependent, and all mechanisms to date depend on some part of the complex intracellular or membrane cytoskeleton.
Not all microparticle release is governed by increases in intracellular calcium. The spontaneous release of microparticles from unstimulated platelets is dependent on αIIβ3 integrin and cytoskeletal turnover, but not on an intracellular calcium signal or activation of calpain (Fig. 2).[46] The microparticles released from unstimulated platelets, however, did retain procoagulant properties and stimulated thrombin generation over a time course of platelet storage. This work from Cauwnberghs and others certainly suggests that microparticles from stored platelets could have consequences for transfusion-related diseases such as transfusion related acute lung injury, which is discussed below.
Annexin-V, which binds phosphatidylserine, is used as a marker for apoptotic cells and is frequently used to analyze microparticles. However, not all microparticles are phosphatidylserine positive. Microparticles also carry membrane markers from their parent cell such as the antigenic clusters of differentiation (i.e., CD31, CD105, etc). Jimenez and others asked an important question in their 2003 paper: Do endothelial cells release phenotypically and quantitatively distinct microparticles dependent on stimulus? Using both macrovascular and microvascular endothelial cells exposed to either growth factor depletion to stimulate apoptosis or tumor necrosis factor-α treatment as cellular activator, they examined total microparticle production and differences in CD and annexin-v staining. They found that regardless of stimulus, macrovascular endothelial cells released fewer microparticles overall than did microvascular endothelium and that in response to apoptotic stimuli constitutive endothelial markers, such as CD31 and CD105, were released. However, in response to cell activation, inducible endothelial markers, such as CD62E, CD54, and CD106, were released on microparticles. These data suggest two important concepts. First, that formation of the microparticles is tightly regulated and responsive to the surrounding environment, and second that analyzing the specific phenotype of microparticles from parental cells may yield valuable information about their potential roles in signaling.[41]
It would be remiss to discuss formation of microparticles without some discussion of how they are cleared from the circulation. As with previous work in microparticles the first studies focused on clearance of platelet microparticles and capitalized on exposure of phosphatidylserine. Blood samples taken at 10, 30, and 60 min from rabbits after injection with isolated biotin-labeled platelets or platelet microparticles indicated there was no increase in the circulating number at any time point.[60] This acute study would suggest that circulating platelet microparticles are cleared/utilized within 10 min. Rand et al.,[60]and others also did an in vitro study mixing the labeled microparticles and platelets in anticoagulated blood. Detection of microparticles in the in vitro setting remained the same as the dose given. This is not surprising in a situation where there are most likely no available clearance mechanisms. However, this study did not address any of the potential mechanisms of clearance, nor did they analyze tissue from the rabbit model to determine whether the microparticles had been taken up by the lungs, liver, spleen, and so on.
The presence of phosphatidylserine on the exposed membrane of platelet microparticles has been exploited to understand potential mechanisms of clearance. Lactadherin, also known as milk fat globule epidermal growth factor-8, is a circulating protein that recognizes phosphatidylserine-expressing cells and membrane fragments and promotes their uptake into macrophages. Lactadherin was found to be bound to circulating CD41+ microparticles (platelet) from normal healthy patients, and further that in vitro addition of lactadherin stimulated a dose-dependent increase in platelet microparticle uptake by THP-1 derived macrophages.[61] Last, lactadherin knockout mice have increased microparticle numbers and are hypercoagulable, producing significantly more thrombin than control.[61] An alternative opsonin implicated in the removal of microparticles is developmental endothelial locus-1 (Del-1).[62] Del-1 is secreted by endothelial cells and functions as a bridging molecule between integrins expressed on endothelial cells and phosphatidylserine. Uptake of platelet microparticles by cultured endothelium was inhibited by a monoclonal antibody to Del-1 and an antibody to αVβ3 integrin. Further, Dasgupta
Investigating the circulating half-life of microparticles on a translational level is challenging due to the ethical considerations of injection of labeled microparticles into a patient. To circumvent this issue, Rank et al.,[63] analyzed blood samples from patients with thrombocytopenia both before and after receiving apherisis platelet concentrates containing high numbers of microparticles. They found that platelet microparticles increased immediately following transfusion, but exhibited a steady decay afterward. The calculated half-life for platelet microparticles was 5.8 h, which is significantly longer than that described in rabbits.[63] It should be noted that the studies performed in rabbits utilized microparticles from stimulated platelets and the patient studies did not. Thus, microparticles from activated platelets may present membrane markers that target them for clearance more readily than those from unstimulated platelets. In all, this work begins to tease out potential mechanisms for clearance and helps us to begin to understand the pitfalls of these experiments.
MICROPARTICLE FUNCTIONS BEYOND THE SCOPE OF COAGULATION
Shedding of microparticles is amplified in response to cellular activation, high shear stress, as well as cellular apoptosis.[64–67] In contrast to their original portrayal as platelet debris, microparticles are now recognized as conveyors of intercellular communication. Microparticles from healthy volunteers serve as circulating pools of microRNAs in the bloodstream. The combination of microRNAs that are expressed in microparticles depends on the parental cell type.[68] A 2007 study also showed that endothelial progenitor cells are capable of producing microparticles that harbor cellular adhesion molecules (Fig. 3). These cell adhesion molecules enabled microparticles to fuse with endothelial cells in vitro. Furthermore, the authors found the endothelial progenitor cell-derived microparticles promoted cellular proliferation, which they attributed to a transfer of mRNA from the microparticles to the cells upon incorporation.[69] Microparticles are also able to influence target cells indirectly by exposing membrane-bound molecules and ultimately activating target cell receptors to induce a specific response (Fig. 3). The Fas antigen and Fas ligand were identified on the surface of vesicles generated by human colorectal cancer cell lines in a 1998 publication. The authors hypothesized in their discussion that microparticles expressing these Fas proteins may serve as a mechanism for long distance communication in the extracellular environment.[70] Additionally, platelet-derived microparticles are capable of promoting proliferation of B cells via CD154 surface expression.[71] The current era of microparticle research is focused predominantly on examining potential mechanistic roles of the small vesicles in diseases.[34,72,73]
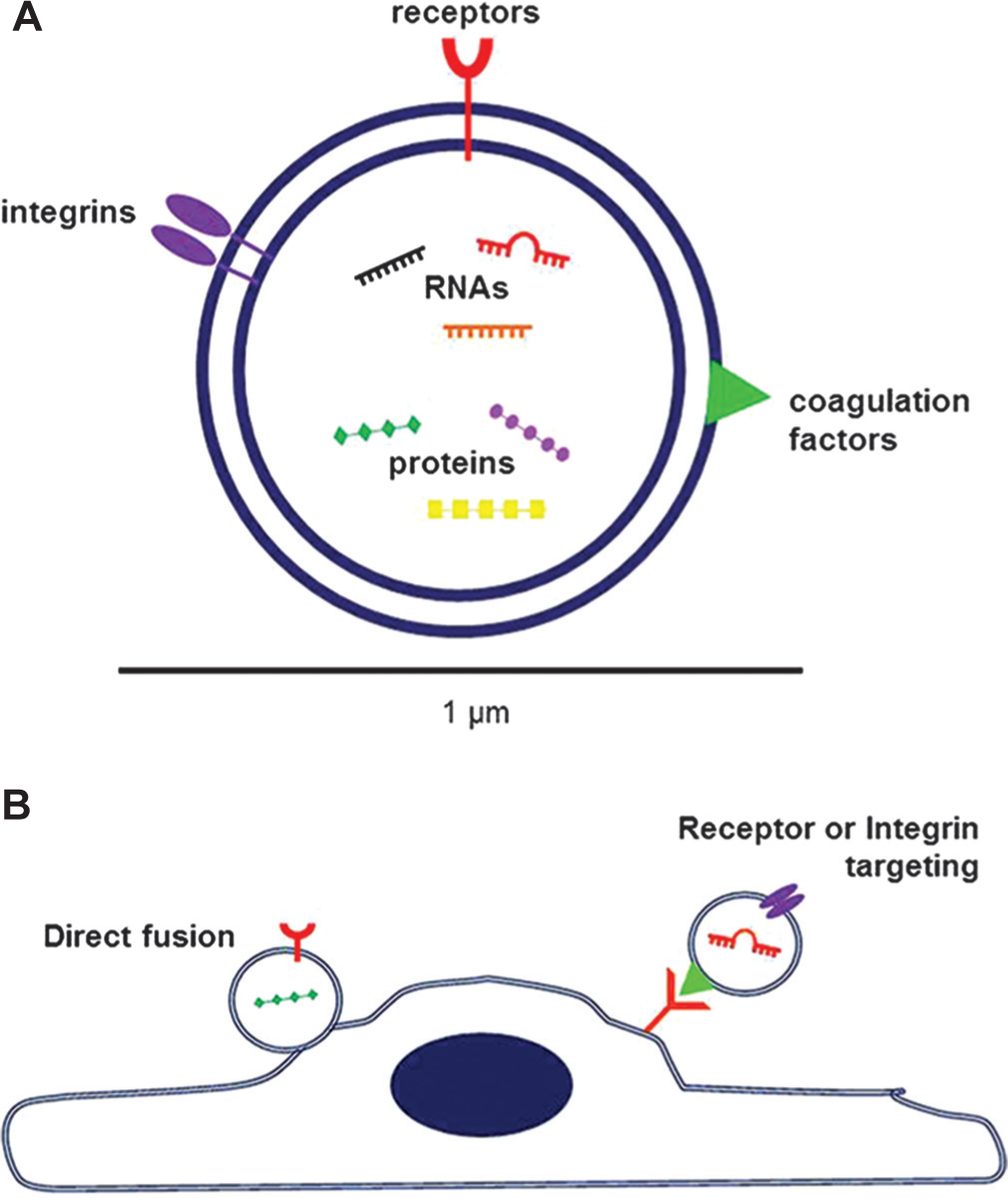
Contents and targeting of microparticles.
Because recent research has focused on microparticles in pathological conditions, the vesicles can easily be mistaken as purely detrimental. However, microparticles can be beneficial and are present in healthy individuals. In fact, a study conducted in 2001 detected a basal level of microparticles in blood samples obtained from healthy male volunteers.[7] Microparticles have the potential to harbor cytoprotective proteins and receptors. Specifically, endothelial microparticles containing activated protein C and the endothelial protein C receptor have been identified. This protein-receptor complex serves as an anticoagulant and can induce expression of anti-inflammatory and cytoprotective genes in target endothelial cells.[74,75] To illustrate the essential roles that microparticles play in health, an excessive bleeding disorder known as Scott syndrome can be considered. Scott syndrome patients suffer from an impaired ability of their platelets to generate microparticles. Without the proper quantity of circulating microparticles, Scott syndrome patients do not experience normal blood coagulation, especially in response to injuries.[76] The key to microparticle benefits is a proper balance of the tiny vesicles. Surpluses or shortages yield detrimental effects.
MICROPARTICLES IN LUNG DISEASE: A DETRIMENTAL IMBALANCE
Microparticles have been highlighted for their positive correlations with various vascular diseases.[72,73,77–79] Specifically in the lung, increased numbers of microparticles have been reported in patients with pulmonary hypertension, ARDS, and lung destruction associated with cigarette smoke exposure. Significantly increased endothelial microparticle numbers are observed in active smokers with reduced diffusion capacities compared to nonsmokers. Additionally, smokers exhibit heightened counts of microparticles originating from the pulmonary capillary endothelium and from apoptotic endothelial cells. Smoking is recognized as a contributor to the development of chronic obstructive pulmonary disease (COPD). The increased microparticle numbers evident in smokers may be indicative of apoptosis of pulmonary capillaries, which could initiate the alveolar damage that is characteristic of COPD.[80] Bronchoalveolar lavage fluid collected from ARDS patients exhibits an increased number of epithelium-derived microparticles. These microparticles from ARDS patients contain high levels of tissue factor and are likely to contribute to the heightened intraalveolar procoagulation activity in ARDS.[8] Circulating microparticles serve as proinflammatory stimuli in acute lung injury (ALI). Interaction between platelet-derived microparticles and neutrophils increases neutrophil secretion of interleukin-8, promotes neutrophil degranulation, and enhances leukocyte rolling in ALI. Endothelial cells are activated by circulating microparticles in ALI leading to increased production of proinflammatory cytokines by the pulmonary endothelium. Furthermore, circulating microparticles are incriminated as contributors to the increased vascular permeability associated with ALI. Platelet-derived microparticles are capable of converting arachidonic acid to thromboxane A2 which promotes pulmonary edema. Increased numbers of circulating platelet-derived microparticles in a murine model of pneumosepsis correlate with higher thromboxane A2 plasma concentrations and vascular permeability in the lungs.[81]
In a recent manuscript by Tantawy et al.,[82], a significant increase in circulating platelet and erythrocyte microparticles was observed to correlate with pulmonary hypertension and acute chest syndrome in patients. Further, patients on current hydroxyurea therapy had lower microparticle numbers indicating microparticles may be a way to measure effectiveness of treatment. It has been reported that endothelial, platelet, and leukocyte microparticles are increased in the circulation of pulmonary hypertension patients.[83] Several studies have additionally correlated the specific populations of endothelial microparticles with mean pulmonary artery pressure, six minute walking distance, and patient survival.[34,44,45] More recently Nadaud et al.,[84] identified a population of circulating small platelet microparticles that were increased in patients with several forms of pulmonary arterial hypertension. Though these small microparticles did not directly correlate with known clinical parameters, their increased levels may be indicative of the pathology of this disease. The role of microparticles in the development or progression of pulmonary arterial hypertension is an active area of investigation. In animal studies using the hypoxia-induced pulmonary hypertension model, microparticles isolated from the circulation inhibited endothelium-dependent vaosreactivity of isolated pulmonary arteries and decreased nitric oxide production in isolated pulmonary endothelial cells.[85] These data would suggest that circulating microparticles may play an active role in PH. Further, microparticles expressing the ectonucleotidase CD39 are increased in patients with idiopathic pulmonary arterial hypertension.[48] Previous work showed that treatment of liver sinusoidal endothelium with CD39-positive microparticles-induced release of cytokines and expression of intercellular adhesion molecule (ICAM) and vascular cell adhesion molecule (VCAM); however, these studies have not been performed with microparticles from idiopathic pulmonary arterial hypertension (IPAH) patients.[86] Further studies will be required to fully understand the contributions of microparticles to pulmonary arterial hypertension (PAH), but with pulmonary hypertension being a devastating disease that is difficult to detect prior to patient decline, these studies suggest that microparticles could potentially serve as biomarkers for early detection of the disease and further our understanding of the pathogenesis of PAH.
Microparticles also contribute to blood transfusion complications, such as transfusion-related acute lung injury (TRALI). TRALI is characterized by a proinflammatory state and the presence of activated neutrophils in the pulmonary vasculature.[87] Red blood cells can release vesicles containing complement receptor 1 in response to ATP-depletion. This phenomenon is thought to occur among red blood cells over storage time as well.[88] Thus, microparticles are capable of mediating pulmonary inflammation via activation of the complement pathway in transfusion recipients, potentially leading to TRALI. In a large animal model of TRALI, injection of LPS followed by infusion of platelet-free supernatant obtained from 5-day-old human blood resulted in the development of lung injury. Infusion of the supernatant also caused a significant increase in circulating neutrophil numbers compared to baseline counts.[89] While this study did not explicitly identify microparticles as the cause of lung injury or neutrophil recruitment, other investigations have identified microparticles as mediators of neutrophil activation. Incubating neutrophils with platelet-derived microparticles caused an interaction between the two, which was captured by fluorescent microscopy. Additionally, this incubation period led to an increase in the phagocytic activity of neutrophils.[90] In summary, microparticles are capable of contributing to the inflammation and neutrophil activation that precedes TRALI.
Microparticles have been investigated in pulmonary pathologies due mainly to their amplified numbers, procoagulant properties, and participation in inflammatory events. A recent study brought attention to another facet of circulating microparticles. Using microparticles from pulmonary hypertensive rats, it was determined that the vesicles are capable of causing endothelial dysfunction. Endothelial dysfunction can be defined as an increased production of vasoconstrictors and a decreased production of vasodilators by the endothelial cells. Additionally, microparticles from pulmonary hypertensive animals impair endothelium-dependent relaxation in pulmonary artery rings.[85] Beyond this particular study, little exploration has focused on how microparticles impact their target cells in the pulmonary circulation. Further, it remains unknown which cells of the pulmonary circulation may be targeted and how. The effects of microparticles on target cells could prove research-worthy in healthy versus pathological conditions. For instance, if microparticlescarried to the vasculature by the blood or generated locallydo contribute to the endothelial dysfunction of pulmonary hypertension they may provide a new therapeutic target leading to a better patient prognosis.
POTENTIAL ROLES FOR MICROPARTICLES IN LUNG HOMEOSTASIS
The pulmonary vascular bed receives 100% of the cardiac output, which includes a multitude of circulating microparticles. One can infer that microparticles take part in conventional activities within the pulmonary vasculature because they are present in healthy circulations in small quantities relative to their numbers in pathological conditions.[4,91] Even in a healthy lung, harmless foreign debris is inhaled and the lung's immune cells must practice tolerance. It has been reported that microparticles are capable of harboring anti-inflammatory constituents.[92,93] Microparticles may also induce the production of anti-inflammatory mediators in their target cells.[75,94] Although microparticles were first identified for their procoagulant properties, recent literature indicates that certain populations of microparticles contain anticoagulant factors.[74,95] Taken together, these anti-inflammatory and anticoagulant characteristics may indicate that healthy microparticles contribute to patency within the pulmonary circulation and, furthermore, play a role in creating an environment where optimal gas exchange can take place.
SUMMARY AND FUTURE DIRECTIONS
Data from microparticle studies in diverse pathologies and model systems is being reported more frequently over time. From their humble beginnings as inert cellular debris and platelet dust, our understanding of microparticles has grown exponentially. Initially thought to be secreted only from platelets and play regulatory roles in coagulation, we now know that microparticles can be released actively from virtually all cells and play important roles in both the physiology and the pathology of systems.
Mechanisms of formation of specific microparticles are yielding interesting results and suggest a multitude of diversity. The membrane and intraparticle constituents suggest highly regulated processes by which microparticles are organized and secreted. Although calcium signaling and cytoskeletal components have thus far been implicated, understanding at a molecular level what comprises a microparticle is of utmost importance. Proteomic and molecular studies are just beginning to unmask the membrane and intravesicle components. Many of these studies are being performed in in vitro settings, but will provide valuable insight into the packaging, message delivery, and possibly targeting mechanisms of microparticles.[47,96,97] Considering their roles in disease and vascular pathology, understanding cell-specific mechanisms of release may provide novel drug targets for prevention of TRALI and possibly pulmonary arterial hypertension.
Also at issue is the downstream targeting of microparticles. A major question that needs to be addressed in future studies is whether microparticles can interact with vascular cells or circulating elements under conditions of active flow to have an effect. Investigations where microparticles were injected into animals and decreased vasoreactivity later examined support the idea that, in fact, microparticles are capable of stimulating their target under physiologic conditions. Studies specifically addressing targeting of microparticles, such as those with the Del-1 knockout, are instrumental in determining how and where they deliver their cargo. As we further develop our comprehension of the heterogeneity of the pulmonary circulation, we may also find vascular segment-specific targets for microparticles. These endothelial targets may be instrumental in our understanding of the pathology of diseases of the pulmonary circulation in which microparticles may contribute.
Several lines of research into whether microparticles can serve as potential biomarkers for severity or contribute to the pathogenesis of pulmonary hypertension, ARDS, and TRALI are promising. The current studies in pulmonary hypertension have focused on correlations with known outcomes such as hemodynamics and 6-minute walk distance. However, we may ultimately find that microparticles are predictive of disease well before clinical symptoms are evident. Understanding the role of microparticles in the propagation of molecular signaling which may perpetuate vascular injury is vital. The work to investigate these concepts is ongoing in our own laboratory and those of others. The ultimate goal is to identify and treat patients at significantly earlier times and potentially identify new therapeutic targets such as the inhibition of a detrimental circulating microparticle. Studies in animal models addressing the role of microparticles in ALI or ARDS have been fruitful and suggest the deleterious microparticles are capable of stimulating pulmonary vascular permeability; however, the mechanisms by which these events occur remain largely unknown. One could also speculate that subsets of microparticles may be beneficial as a biomarker to assist in further clarification of the definition of ARDS. Last, speculation that microparticles could play an important role in the onset of TRALI is providing an excellent window of opportunity to further our understanding of the nonimmune-related arm of this disease. The implication that phosphatidylserine exposing blood products, such as microparticles, could be vital contributors is intriguing.[98]
Overall, advancement in our understanding of the constituents, generation, and targeting of microparticles will be of great benefit. The potential for microparticles to become either drug targets or drug carriers is an exciting concept that opens many lines of investigation and will ultimately benefit seriously ill patients.
Footnotes
ACKNOWLEDGEMNTS
Thank you, Dr. Ivan McMurtry for your kind review, Dr. Abdallah Alzoubi for your contributions to the article figures, and Dr. Judy King for your contribution to the electron microscope images of microparticles featured in this article.