
Abstract
Islet transplantation has been proven to be an effective treatment for patients with type 1 diabetes, but a lack of islet donors limits the use of transplantation therapies. It has been previously demonstrated that hepatocytes can be converted into insulin-producing β-like cells by introducing pancreatic transcription factors, indicating that direct hepatocyte reprogramming holds potential as a treatment for diabetes. However, the efficiency at which functional β-cells can be derived from hepatocyte reprogramming remains low. Here we demonstrated that the combination of Pdx1 and Ngn3 can trigger reprogramming of mouse and human liver cells to insulin-producing cells that exhibit the characteristics of pancreatic β-cells. Treatment with PDGF-AA was found to facilitate Pdx1 and Ngn3-induced reprogramming of hepatocytes to β-like cells with the ability to secrete insulin in response to glucose stimulus. Importantly, this reprogramming strategy could be applied to adult mouse primary hepatocytes, and the transplantation of β-like cells derived from primary hepatocyte reprogramming could ameliorate hyperglycemia in diabetic mice. These findings support the possibility of developing transplantation therapies for type 1 diabetes through the use of β-like cells derived from autologous hepatocyte reprogramming.
Keywords
Introduction
Type 1 diabetes mellitus is characterized by complete loss of β-cells due to T-cell-mediated autoimmune attack, which leads to insulin deficiency. Transplantation of islets isolated from the cadaveric donor pancreas has been proven to be a promising treatment for patients with type 1 diabetes mellitus1–3. By 2009, more than 700 patients had received islet transplants; by a year after transplantation, over half of the patients did not need to receive additional insulin injections, and thus experienced an improvement in quality of life 4 . However, such transplantation therapy is severely limited by the lack of islet donors. Moreover, patients who received allogeneic islet transplantation suffered from the side effects of immunosuppressive medications 5 . Hence, the ability to produce large amounts of β-cells from autologous resources would be helpful in the development of cell-based therapeutics for type 1 diabetic patients. Pancreatic and duodenal homeobox 1 (Pdx1) is an essential transcription factor involved in the initial development of pancreatic progenitors and the maintenance of β-cell functions6,7. Previously, we and others have reported that liver cells can be converted into insulin-producing β-like cells through the introduction of Pdx18,9. Moreover, other pancreatic transcription factors, such as neurogenin-3 (Ngn3) and neuronal differentiation 1 (NeuroD), which are involved in β-cell development, have also been shown to induce the transdifferentiation of liver cells into insulin-producing β-like cells10–12. However, the efficiency of reprogramming liver cells to insulin-producing cells using these transcription factors was reported to be low10,13.
Expression of Ngn3 during the secondary transition of pancreatic development is associated with the rapid expansion of endocrine tissues 14 . Pancreatic duct ligation of mice revealed that Ngn3 is expressed in proliferating β-cells during pancreas regeneration15,16. These findings indicate the pivotal role of Ngn3 in expanding pancreatic endocrine progenitors. Additionally, platelet-derived growth factor (PDGF) can promote DNA synthesis of cultured pancreatic islet cells17,18. Recent studies have demonstrated that platelet-derived growth factor receptor a (PDGFRα)-mediated signaling is crucial for the proliferation of β-cells in the pancreas of neonatal mice 19 . Moreover, glucagon-like peptide-1 (GLP-1), an incretin hormone secreted from intestinal L cells in response to food intake, is known to enhance insulin biosynthesis and secretion from β-cells20–23. Additionally, GLP-1 has been shown to stimulate β-cell proliferation in diabetic rodents24,25 and to protect β-cells from apoptosis caused by glucotoxicity or high cholesterol26–28.
In the current study, we evaluated the possibility of improving reprogramming efficiency by combining Pdx1 and Ngn3 with activated PDGFRα and GLP-1 receptor (GLP-1R) signaling. We demonstrate that combined use of Pdx1 and Ngn3 in GLP-1-containing medium can reprogram the α mouse liver 12 (AML12) cells, mouse primary hepatocytes, and human C3A liver cells (the HepG2/C3A cell line is a derivative of HepG2 cells) into insulin-producing cells, which displayed the characteristics of pancreatic β-cells. The reprogrammed cells were found to express PDGFRα. Addition of PDGF-AA (homodimers of platelet-derived growth factor-A) not only facilitated the reprogramming of hepatocytes into β-like cells but also enhanced the ability of reprogrammed cells to secrete insulin in response to glucose stimulus. Importantly, we also established that insulin-producing β-like cells derived from adult hepatocyte reprogramming were functional and may therefore be of use in diabetic transplantation therapies.
Materials and Methods
Preparation of Adenoviruses
Three adenoviral constructs (pAd), (1) a control vector, pAd-nGFP, containing the cytomegalovirus (CMV) promoter-driven nuclear-formed green fluorescent protein (nGFP), (2) a mouse Pdx1 expression vector, pAd-Pdx1-IRES-nGFP, containing the inserted sequence of Pdx1-internal ribosome entry site (IRES)-nGFP, and (3) a mouse Ngn3 expression vector, pAd-Ngn3-IRES-nGFP, containing the inserted sequence of Ngn3-IRES-nGFP, which were originally provided by Dr. Douglas Melton 29 , were obtained from Addgene (Cambridge, MA, USA). PacI-digested adenoviral constructs were then transfected into 293A cells (Invitrogen, Waltham, MA, USA) with the use of Genejuice transfection reagents (Novagen, Madison, WI, USA). Purification of recombinant adenoviruses was performed by double cesium chloride (CsCl) gradient centrigufation. CsCl gradient centrifugation was performed by gently adding 1.4 g/ml density of CsCl (Sigma-Aldrich, St. Louis, MO, USA), 1.2 g/ml density of CsCl, and viruses containing supernatant from bottom to top into polyallomer tubes (Beckman Coulter, Brea, CA, USA) to finally form a three-layer solution, and this CsCl solution was further ultracentrifuged for 16 h. Thereafter, the virus band at a density of 1.33 g/ml was harvested. After removal of CsCl by dialysis, the recombinant adenovirus stock was stored at −80°C; the titer of recombinant adenoviruses was determined using the plaque assay with 293A cells. A 10-fold serial dilution of purified adenoviruses was performed, and then the 10 μl of 106 to 109 diluted viruses was separately added into each well of 293A cells. Subsequent to infection for 24 h, low-melting temperature agarose (Lonza, Rockland, ME, USA) was utilized to overlay onto the virus-infected cells, and the plaques, cellular dead zones caused by adenoviral lysis of host cells, were calculated after 1 week.
Cell Culture and Treatment
The AML12 cell line [AML12 cells, established from hepatocytes of human transforming growth factor-α (TGF-α) transgenic CD1 mice] and human HepG2/C3A liver cell line were both obtained from the Food Industry Research and Development Institute (FIRDI, Hsinchu, Taiwan). These two liver cell lines display phenotypes of mature hepatocytes, including expression of albumin or transferrin30,31. AML12 cells were maintained in Dulbecco's modified Eagle's medium (DMEM)/F12 containing penicillin and streptomycin (Invitrogen), 10% fetal bovine serum (FBS; Gibco, Waltham, MA, USA), 1 μmol/L of dexamethasone (Sigma-Aldrich), 5 μg/ml of apo-transferrin (Sigma-Aldrich), 5 ng/ml of selenium (Sigma-Aldrich), and 5 μg/ml of insulin (Sigma-Aldrich). HepG2/C3A cells were grown in minimum essential medium (MEM) (Gibco) with penicillin, streptomycin, 10% FBS, 0.1 mM of nonessential amino acids (Gibco), and 1.0 mM of sodium pyruvate (Gibco). NIT-1 cells (FIRDI), a pancreatic β-cell line of NOD/Lt mice, were seeded in Ham's F12K (Gibco) with penicillin, streptomycin, and 10% 56°C heat-inactivated FBS. βTC6 cells (FIRDI), an insulinoma cell line of SV40 large T transgenic mice, were cultured in DMEM containing penicillin, streptomycin, and 15% 56°C heat-inactivated FBS. All media were replaced every other day.
Mouse Experiments
All animal experiments were approved by the Academia Sinica Institutional Animal Care and Use Committee (IACUC). Both ICR, a strain of albino mice that were initially established at the Institute of Cancer Research and were named by the first letter of the institute, and C57BL/6 mice were obtained from the National Laboratory Animal Center (Taipei, Taiwan). A single intraperitoneal (IP) injection of streptozotocin (STZ) (200 mg/kg body weight) was used to induce diabetes in C57BL/6 mice. On day 4 after STZ injection, hyperglycemia was defined as a nonfasting blood glucose level higher than 300 mg/dl. Next, 1 × 105 of green fluorescent protein (GFP)+-reprogrammed cells were transplanted into the kidney capsule of diabetic mice. Blood glucose levels were measured every 2 days posttransplantation, and a glucose tolerance assay was performed on day 8 posttransplantation.
Isolation and Culture of Primary Hepatocytes
Primary hepatocytes were isolated from 8- to 12-week-old C57BL/6 mice using a two-step collagenase perfusion protocol as described previously 32 . Liver cells obtained from perfusion were initially analyzed by flow cytometry to confirm that over 99% of the cells were hepatocytes that stained positive for albumin and CYP7A. Primary hepatocytes were then cultured on dishes coated with collagen I (BD Biosciences, Franklin Lakes, NJ, USA) in DMEM/F12 (Gibco) supplemented with 10% FBS, 5 μg/ml of apo-transferrin, 5 ng/ml of selenium, 5 μg/ml of insulin, 10 nM of dexamethasone, 2 mM of L-glutamine (Gibco), 50 nM of 2-mercaptoethanol (2-ME), and 1× NEAA (Gibco).
Induction of Hepatocyte Reprogramming
Hepatocyte reprogramming toward the β-cell lineage was triggered by infecting mouse primary hepatocytes or liver cell lines, including AML12 and HepG2/C3A cells, with a dosage of 3 or 50 multiplicity of infection (MOI) of each adenovirus (carrying either nGFP, Pdx1-IRES-nGFP, or Ngn3-IRES-nGFP). The adenovirus-containing medium was replaced with medium that contained 25 mM of glucose (USB Corporation, Cleveland, OH, USA), 1 nM of exendin-4 (Sigma-Aldrich), 30 nM of GLP-1 (American Peptide Company, Sunnyvale, CA, USA), and 20 nM of PDGF-AA (ProSpec Protein Specialists, Rehovot, Israel) on the following day.
Immunofluorescent Staining
Cells were fixed with 4% paraformaldehyde (PFA; Sigma-Aldrich) for 30 min at room temperature (RT) and then permeabilized using 0.1% Triton X-100 (Sigma-Aldrich) for 20 min. After blocking with 5% bovine serum albumin (BSA; USB Corporation) for 1 h, the cells were incubated with primary antibodies overnight and secondary antibodies for 1 h at RT. The dilutions of primary and secondary antibodies are shown in Table 1. Slides were imaged on a Zeiss LSM 780 confocal microscope (Carl Zeiss, Thornwood, NY, USA).
Antibodies for Immunostaining and Flow Cytometry

Western Blotting Analysis
Five days postinfection, cells were lysed with radioimmunoprecipitation assay (RIPA) buffer, and 30 μg of proteins was separated by 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE). Subsequent to transfer to polyvinylidene difluoride (PVDF) membranes (EMD Millipore, Billerica, MA, USA), membranes were blocked in 5% nonfat milk in Tris-buffered saline (TBS) for 1 h. The membrane was then incubated with primary antibodies in blocking buffer at 4°C overnight. On the next day, the membrane was washed and then incubated with appropriate secondary antibodies in blocking buffer for 1 h at RT. The blot was developed using Western Chemiluminescent HRP Substrate (EMD Millipore). The antibodies are summarized in Table 1.
RNA Isolation, RT-PCR, and Real-Time RT-PCR
Total RNA was extracted using a TRIzol reagent according to the manufacturer's instructions (Invitrogen). Purified RNA was then reverse transcribed using modified Moloney murine leukemia virus (M-MLV) reverse transcriptase (Thermo Fisher Scientific, Waltham, MA, USA) to obtain cDNAs for use in PCR. All the PCR products were separated by electrophoresis on a 2% agarose gel and stained with ethidium bromide for analysis. Ten nanograms of cDNA was mixed with the KAPA SYBR master mix (Kapa Biosystems, Wilmington, MA, USA) to perform real-time PCR using the ABI-7900 machine. In order to detect expression of endogenous pdx1 and ngn3 in reprogrammed β-like cells, we designed two pairs of primers that could either anneal to the 5′ untranslated region (5′-UTR) of the mouse pdx1 or ngn3 mRNA sequences. The sequences of primer pairs are listed in Table 2.
Primers for RT-PCR Analysis

EdU Assay
Cell proliferation of reprogrammed β-like cells was examined using Click-iT EdU assays (Invitrogen). For EdU labeling at day 5 postinfection, adenovirus-infected liver cells were incubated with 10 μM of EdU for 1 h at 37°C. The cells were then fixed with 4% PFA for 30 min at RT. The reacting EdU with azide-Alexa 647 dye was performed using the Click-iT Kit protocol, after which cells were treated with 1 μg/ml of DAPI and 1 μg/ml of RNaseA for 30 min. FACS CantoII (BD Biosciences) was used to detect the percentage of EdU-labeled cells (indicating cell entry into S phase).
Insulin Secretion Assay
Liver cells were seeded in 12-well plates (Falcon@ Brand Products, Corning, Corning, NY, USA) at 105 cells per well, and 50 MOI of each adenovirus was used to infect one well of hepatocytes at 24 h after cell plating. The original media were removed at 5 days postinfection, and the cells were washed with Krebs–Ringer bicarbonate (KRB) buffer supplemented with 0.1% BSA (Gibco). The cells were then incubated in fresh KRB with 5 mM or 25 mM of glucose, or 45 mM of KCl at 37°C for 1 h. At the end of the incubation period, the insulin content in the cultured KRB buffer was measured by using ultrasensitive mouse insulin enzyme-linked immunosorbent assay (ELISA) kit (Mercodia, Uppsala, Sweden) according to the manufacturer's protocols. Subsequently, cell protein concentration for each well was determined by Bradford protein assay (Bio-Rad, Hercules, CA, USA), and values of insulin content were normalized to total protein content. The stimulation index of insulin was calculated as a ratio of higher glucose-stimulated insulin secretion levels (25 mM of glucose) to the levels of basal insulin secretion (5 mM of glucose), for the purposes of evaluating the insulin secretion capabilities of reprogrammed β-like cells in response to increased glucose levels.
Statistical Analysis
All values are shown as the mean ± standard error of the mean (SEM). Values of p for comparison between control and experimental groups were calculated using the Kruskal–Wallis test or two-way analysis of variance (ANOVA) by GraphPad Prism 6.0 software (GraphPad Software, Inc., La Jolla, CA, USA). The Bonferroni's multiple comparisons test was used to perform the post hoc analysis after ANOVA. A value of p < 0.05 was regarded as significant.
Results
Pdx1 and Ngn3 Induce Reprogramming of Mouse Liver Cells Into PDGFRα-Expressing β-Like Cells
Initially, we investigated reprogramming efficiencies resulting from overexpressing various combinations of pancreatic transcription factors that were previously shown by adenoviral infection strategies to be involved in differentiation or proliferation ofβ-cells. Mouse AML12 cells infected with Ad-Pdx1, Ad-Ngn3, or simultaneous administration of Ad-Pdx1 and Ad-Ngn3 (Ad-NP) and cultured in medium containing 22.5 mM of glucose were examined 5 days postinfection. Immunofluorescent staining results revealed that infection with Ad-NP significantly induced production of insulin-expressing cells (8.47 ± 2.90% of total population) in which the production rate was 2.06- or 2.48-fold higher than in AML12 cells infected with Ad-Pdx1 or Ad-Ngn3 alone, respectively (Fig. 1A). To further characterize the β-cell characteristics of the insulin-producing cells derived from reprogramming, we performed RT-PCR analysis on 1.5 × 106 Ad-Pdx1-, Ad-Ngn3- and Ad-NP-infected GFP+ cells and on 1,000 islets isolated from 4-week-old ICR male mice. We found that Ad-NP could efficiently induce expression of both insulin1 and insulin233,34, compared with the effects of Ad-Pdx1 and Ad-Ngn3 infection alone. In addition to insulin expression, infection with Ad-NP also induced expression of other β-cell markers, including nkx2.2, pax4, rfx6, islet amyloid polypeptide (iapp), prohormone convertase 1/3, and kir6.2 (ATP-sensitive K+ channels involved in glucose-sensing function in mature β-cells). Conversely, infection with Ad-Pdx1 alone slightly induced expression of someβ-cell markers. Importantly, infection with Ad-Pdx1 alone was unable to induce expression of the mature β-cell marker, rfx6. In contrast, the α-cell hormone glucagon exhibited remarkable induction upon infection with Ad-Pdx1 alone. Most importantly, both endogenous pdx1 and ngn3 were successfully induced in Ad-NP-infected cells, suggesting that these cells may be able to produce insulin in the long term (Fig. 1B).
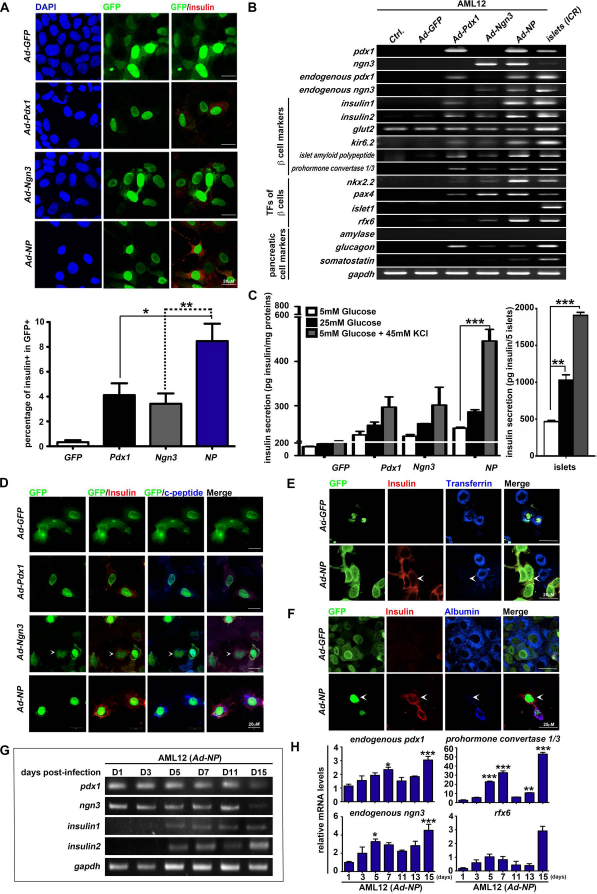
Ectopic expression of Pdx1 and Ngn3 enhances reprogramming of mouse liver cells to insulin-secretingβ-like cells. Mouse AML12 liver cells were infected with Ad-nGFP, Ad-Pdx1, or Ad-Ngn3 or were coinfected with Ad-Pdx1 and Ad-Ngn3 (shown as Ad-NP) for 5 days. (A) The reprogrammed cells were stained with anti-insulin antibodies (red). GFP marked adenovirus-infected cells. Nuclei were counterstained with DAPI (blue). The graph below shows the percentage of insulin and GFP+ cells. Statistical significance was calculated using Kruskal–Wallis test, and the data are shown as the mean ± SEM; *p < 0.05 and **p < 0.01. (B) RT-PCR was performed to analyze selected marker genes, including β-cell markers (insulin1, insulin2, glut2, kir6.2, islet amyloid polypeptide, and prohormone convertase 1/3), β-cell expressing transcription factors (pdx1, ngn3, endogenous pdx1, endogenous ngn3, nkx2.2, pax4, islet1, rfx6), and other pancreatic cell markers (amylase, glucagon, somatostatin), as well as an internal control gene (gapdh), in control and reprogrammed cell groups. Islets isolated from 4-week-old ICR mice were used as a positive control for pancreatic gene expression. Ctrl. refers to untreated AML12 cells. (C) Measurement of insulin secretion from reprogrammed cells cultured in medium containing 5 mM glucose (white bars), 25 mM glucose (black bars), and 5 mM glucose with 45 mM KCl (gray bars). Five islets were used as a positive control. Insulin secretion values were normalized to total proteins. Data were analyzed from at least three independent experiments. Two-way ANOVA was utilized to confirm statistical significance, and the data are shown as the mean ± SEM; ***p < 0.001 indicates that the difference between the 5 mM glucose group and the 45 mM KCl group was statistically significant. (D–F) On day 5 postinfection, the reprogrammed cells were stained with anti-insulin (red) and (D) anti-C-peptide (blue), (E) anti-transferrin (blue), or (F) anti-albumin (blue) antibodies. White arrowheads indicate insulin-positive cells that have lost (E) transferrin or (F) albumin expression. (G, H) Time course experiments to determine insulin expression in reprogrammed cells. Total RNA of AML12 cells that were infected with Ad-NP was extracted at days 1, 3, 5, 7, 11, 13, and 15. (G) RT-PCR was performed to determine expression of total pdx1, total ngn3, insulin1, insulin2, and gapdh. (H) Real-time quantitative RT-PCR was performed to detect expression of endogenous pdx1, endogenous ngn3, rfx6, and prohormone convertase 1/3 transcripts in Ad-NP-infected AML12 cells. Statistical significance was calculated using Kruskal–Wallis test, and data are shown as the mean ± SEM; *p < 0.05, **p < 0.01, and ***p < 0.001 indicate that the difference between mRNA levels at day 1 postinfection and the indicated time point is statistically significant.
In mammals, functional β-cells synthesize, store, and secrete insulin in response to physiological stimuli. To further evaluate the functional status of reprogrammed cells, we examined insulin secretion capabilities in response to glucose stimulation (5 mM or 25 mM of glucose) or potassium concentration (5 mM of glucose with 45 mM of KCl). On day 5 postinfection, β-like cells induced by Ad-NP treatment secreted 1.06-, 1.12-, and 1.5-fold higher amounts of insulin compared to reprogrammed cells generated by Ad-Pdx1, under conditions of 5 mM of glucose, 25 mM of glucose, and 5 mM of glucose + 4 mM of KCl, respectively (Fig. 1C). The stimulation index of insulin was further calculated to analyze the insulin-releasing capability upon changes in glucose levels (from 5 mM to 25 mM). We found that Ad-NP-infected cells had a stimulation index of 1.38, less than that of the control islet cells (2.20) (Fig. 1C). Expression of C-peptide, a by-product during insulin biosynthesis, could also be detected in Ad-NP-induced β-like cells (Fig. 1D). To further determine if reprogramming triggered by Pdx1 and Ngn3 can indeed switch liver cell fate toward β-cell fate, we assessed whether the reprogrammed β-like cells lost their hepatic phenotypes through immunofluorescent staining. We observed that some insulin-positive cells induced by Ad-NP lost transferrin and albumin expression (Fig. 1E and F, cells indicated by arrows). Additional time course experiments further revealed that Ad-NP-infected cells started to express both insulin1 and insulin2 transcripts as early as day 5 postinfection, and continuously expressed insulin even after exogenous expression of pdx1 or ngn3 was lost at day 15 postinfection (Fig. 1G). Real-time quantitative RT-PCR analysis further revealed that expression of endogenous pdx1, endogenous ngn3, prohormone convertase 1/3, and rfx6 was induced in Ad-NP-infected cells from day 3 to day 5 postinfection and were maintained after exogenous expression of pdx1 or ngn3 was lost at day 15 postinfection (Fig. 1H). These findings indicate that lineage reprogramming of liver cells to functional β-like cells can be triggered by Pdx1 and Ngn3.
A recent study showed that a reduction of PDGFRα is correlated with an age-dependent loss of β-cell proliferation in mice 19 , indicating that PDGFRα-mediated signaling plays a role in regulating β-cell expansion in young mice. Moreover, GLP-1R is expressed in β-cells, and GLP-1 is known to regulate insulin biosynthesis and secretion from β-cells20–23. Therefore, we investigated whether insulin-producing cells generated by Ad-NP express PDGFRα and GLP-1R. AML12 liver cells were harvested on day 5 postinfection for real-time RT-PCR, revealing that reprogrammed cells generated by Ad-NP or Ad-Pdx1 infection expressed significantly higher levels of glp-1r compared with the Ad-GFP- or Ad-Ngn3-infected cells (Fig. 2A). Additionally, RT-PCR analysis showed that AML12 cells expressed pdgf-a, and infection of liver cells with Ad-NP further induced expression of pdgfrα (NIH3T3 cells and islets derived from 4-week-old mice were used as positive controls for examining pdgfrα expression) (Fig. 2B). Immunofluorescent staining confirmed that some insulin-producing cells induced by Ad-NP expressed PDGFRα at higher levels than those observed in the reprogrammed cells induced by Ad-Pdx1 or Ad-Ngn3 (Fig. 2C). FACS analysis further confirmed that liver cells did not express PDGFRα; after transduction of Ad-NP, about 2.23% of the Ad-NP-infected cells (GFP+ cells) were positive for PDGFRα (Fig. 2D and E). Based on these findings, it appears that the combination of Pdx1 and Ngn3 is sufficient to reprogram liver cells into insulin-secreting β-like cells; however, the reprogramming efficiency remained low. Though data obtained from our characterization experiments suggest that insulin secretion by Ad-NP-induced β-like cells in response to changes in glucose is not comparable to that of islet cells, insulin-producing cells induced by Ad-NP possess PDGFRα-expressing characteristics reminiscent of those of β-cells in young mice, suggesting that these insulin-producing β-like cells may be modulated by PDGFRα or GLP-1R signaling.

Combined expression of Pdx1 and Ngn3 induces PDGFRα and GLP-1R in reprogrammed cells. Mouse AML12 liver cells were infected with Ad-nGFP, Ad-Pdx1, or Ad-Ngn3 or were coinfected with Ad-Pdx1 and Ad-Ngn3 (shown as Ad-NP) for 5 days. (A) Real-time quantitative RT-PCR was performed to detect expression of glp1 receptors in control (shown as Ctrl.) and reprogrammed cells. The Kruskal–Wallis test was utilized to confirm statistical significance, and the data are shown as the mean ± SEM; *p < 0.05 and **p < 0.01 indicate that the difference between the Ad-NP-infected cells and the Ad-GFP or Ad-Ngn3-infected cells was statistically significant. n.s., not significant. (B) Expression of pdgfrα and pdgf-a in control and reprogrammed cell groups was analyzed by RT-PCR. NIH3T3 and islets of 4-week-old ICR mice were used as a positive control for pdgfrα and pdgf-a. (C) On day 5 postinfection, reprogrammed cells were stained with anti-insulin (red) and anti-PDGFRα (pink) antibodies. Arrows indicate Ad-NP-induced insulin-positive cells displaying expression of PDGFRα. GFP marked adenovirus-infected cells. Nuclei were counterstained with DAPI (blue). (D and E) FACS analysis was performed to analyze surface expression of PDGFRα on reprogrammed cells derived from infection with Ad-GFP, Ad-Pdx1, Ad-Ngn3, or Ad-NP. (E) Averaged percentage of surface expression of PDGFRα on reprogrammed cells was quantified from three independent experiments. Ctrl. refers to noninfected AML12 cells. The Kruskal–Wallis test was utilized to confirm statistical significance, and the data shown represent the mean ± SEM; ***p < 0.001 indicates that the difference between the Ad-NP-infected group and the control group was statistically significant.
PDGF Facilitates Reprogramming of Liver Cell to β-Like Cells
Since the combination of Pdx1 and Ngn3 was sufficient to reprogram liver cells into insulin-producing β-like cells expressing PDGFRα or GLP-1R, we next evaluated if the addition of PDGF-AA, GLP-1, or exendin-4 (GLP-1R agonists) would either enhance the efficiency of generating insulin-producing β-like cells or increase the insulin secretion capability of β-like cells in response to glucose stimulation. AML12 cells infected with Ad-NP were cultured in basal medium (DMEM/F12 medium) with or without 20 nM of PDGF-AA, 30 nM of GLP-1, 1 nM of exendin-4, and/or 25 mM of glucose. We report that treatment with PDGF-AA resulted in a 3.5-fold increase in the percentage of insulin-positive cells in GFP+ cells (Ad-NP-infected) in comparison to that in Ad-NP-infected cells cultured in the basal medium. Treatment of Ad-NP-infected cells with PDGF-AA in GLP-1/exendin-4-containing high glucose medium resulted in a further 4.77-fold increase in the percentage of insulin-positive cells compared with that in Ad-NP-infected cells cultured in basal medium (Fig. 3A and B). We then investigated if the addition of PDGF-AA, GLP-1, and exendin-4 could further enhance insulin secretion by Ad-NP-reprogrammed cells in response to glucose stimulation. Exposure of reprogrammed cells generated in basal medium to 25 mM of glucose resulted in a secretion of 351 ± 13 pg insulin/mg total proteins. Treatment of GLP-1 or PDGF-AA significantly increased the capability of reprogrammed cells to secrete insulin in response to 25 mM of glucose (GLP-1: 600 ± 72; exendin-4: 549 ± 39; PDGF-AA: 725 ± 43) and enhanced the stimulation index of insulin to 4.61 or 3.58, respectively (Fig. 3C and D). Treatment of Ad-NP-infected cells with PDGF-AA in GLP-1/exendin-4-containing medium resulted in a 1.66fold increase in insulin secretion in response to 25 mM of glucose compared to that of Ad-NP-infected cells cultured in basal medium; the stimulation index of insulin was actually increased to 5.84 (Fig. 3C and D). The findings demonstrate that activation of PDGFRα together with GLP-1 treatment can further promote reprogramming of liver cells to insulin-secreting β-like cells that can respond to increased glucose levels.

Treatment with PDGF-AA enhanced Pdx1 and Ngn3-induced reprogramming of mouse liver cells to insulin-secretingβ-cells. Mouse AML12 liver cells were coinfected with Ad-Pdx1 and Ad-Ngn3 (shown as Ad-NP). At 1 day postinfection, Ad-NP-infected cells were cultured in basal medium supplemented with the indicated combination of 25 mM glucose, 30 nM GLP-1, 1 nM exendin-4, and 20 nM PDGF-AA for 4 more days. AML12 control refers to noninfected cells cultured in original liver cell culture medium, and basal medium refers to DMEM/F12 supplemented with 10% FBS and antibiotics. (A and B) The reprogrammed cells were stained with anti-insulin antibodies (red) on day 5 postinfection. GFP marks adenovirus-infected cells. Nuclei were counterstained with DAPI (blue). Quantitative analysis results are shown in (B). Kruskal–Wallis tests were used to confirm statistical significance, and data are shown as the mean ± SEM; ***p < 0.001 indicates that the difference between basal medium and the PDGF-AA group was statistically significant. (C) Insulin secretion assay was performed on control AML12 cells and Ad-NP-reprogrammed cells in basal medium supplemented with the indicated combination of 25 mM glucose, 30 nM GLP-1, 1 nM exendin-4, and 20 nM PDGF-AA. Measurement of insulin secretion in response to 25 mM glucose was performed in KRB buffer. Insulin secretion values were normalized to total proteins (***p < 0.001). (D) The stimulation index of insulin was calculated as a ratio of glucose-stimulated insulin-secreting levels (25 mM glucose) to the basal insulin-secreting levels (5 mM glucose). Data was averaged from at least three independent experiments. Kruskal–Wallis tests were used to confirm statistical significance, and data are shown as the mean ± SEM; *p < 0.05 and ***p < 0.001 indicate that the difference between the basal medium group and GLP-1- of PDGF-AA-treated groups was statistically significant. (E) On day 5 postinfection, Ad-NP-infected cells treated with or without PDGF-AA (shown as w/o PDGF-AA) were treated with 10 μM EdU. EdU and DAPI double staining were performed to determine the cell cycle profile by FACS analysis (top) and immunofluorescence staining [bottom, EdU (cyan) and insulin (red)]. EdU+ cells indicate cells entering S phase. The reprogrammed cells were gated and analyzed based on the expression of GFP [GFP– (white bars) and GFP+ (green bars)]. NIT1 cells treated with or without PDGF-AA represent actualβ-cells (***p < 0.001). (Bottom) Data was analyzed from at least three independent experiments. Two-way ANOVA tests were applied to confirm statistical significance, and the data are shown as the mean ± SEM; ***p < 0.001 indicates that the difference between the control and PDGF-AA groups was statistically significant. (F) Western blot analysis was performed to determine the expression of PDGFRα, phosphorylated PDGFRα, and CDK4 in AML12 cells infected with Ad-GFP, Ad-Pdx1, Ad-Ngn3, or Ad-NP with or without PDGF-AA treatment (shown as w/o PDGF-AA). GAPDH was used as an internal control. Quantification of the levels of PDGFRα or phosphorylated PDGFRα by densitometry is shown on the right of (F). Kruskal–Wallis tests were used to confirm statistical significance, and the data are shown as the mean ± SEM; *p < 0.05, **p < 0.01, and ***p < 0.001 indicate that the difference in the level of PDGFRα or phosphorylated PDGFRα was statistically significant.
PDGF has been demonstrated to possess the capacity to promote DNA synthesis in cultured islets17,18. We analyzed whether PDGF-AA treatment enhances liver cell reprogramming through increased proliferation of β-like cells by administering ethynyl deoxyuridine (EdU) on day 5 postinfection and then measuring the percentage of reprogrammed cells entering the S phase under PDGF-AA treatment. In the control groups, addition of PDGF-AA did not increase cell proliferation in GFP– cells or GFP+ (Ad-GFP infected) cells. Infection with Ad-NP slightly increased the percentage (12.96%) of GFP+ cells entering the S phase (EdU+) (Fig. 3E), which we hypothesize as maybe because AML12 cells expressed low levels of PDGF-AA (Fig. 2B). Treatment of Ad-NP-infected cells with PDGF-AA resulted in a 1.81-fold increase in the proportion of GFP+ cells entering the S phase (EdU+) compared to untreated Ad-NP-infected cells, and immunofluorescence staining confirmed there was an increase in EdU-labeled insulin-producing cells after treatment with PDGF-AA. We also confirmed that PDGF-AA has the capacity to promote the entry of mouse NIT1 β-cells into the S phase (Fig. 3E). Additionally, Western blot analysis further confirmed that PDGF-AA treatment also significantly increased PDGFRα phosphorylation in Ad-NP-reprogrammed cells by 2.54-fold compared with Ad-NP infection alone, and upregulation of CDK4 was also observed. The above results confirm that PDGF signaling is activated in the reprogrammed cells (Fig. 3F) and imply that activated PDGFRα signaling may play a role in triggering the expansion of insulin-secretingβ-like cells during liver cell reprogramming.
Reprogramming of Human Liver Cells Into Insulin-Secreting β-Like Cells
We examined whether infection of human HepG2/C3A cells with Ad-Pdx1, Ad-Ngn3, or Ad-NP and incubation in GLP-1/exendin-4-containing high-glucose medium could enhance reprogramming into insulin-secreting β-like cells. Immunofluorescent staining revealed that 38.9 ± 2.5% of GFP+ (Ad-NP infected) cells coexpressed insulin, which was 4.96- and 3.91-fold higher than that of cells in the Ad-Pdx1 (7.9 ± 4.4%) and Ad-Ngn3 (9.9 ± 2.5%)-infected groups, respectively (Fig. 4A–C). Notably, these insulin-producing cells derived from Ad-NP infection formed clusters resembling islet-like clusters (Fig. 4A and B). Importantly, PDGF-AA treatment resulted in a 6.06-fold increase in the capability of Ad-NP-induced β-like cells to secrete insulin in response to 25 mM of glucose compared with Ad-NP-infected cells cultured in basal medium (Fig. 4D). Additionally, either PDGF-AA or exendin-4 treatment further induced a 6.14- or 4.14-fold enhancement, respectively, in the stimulation index of insulin in Ad-NP-infected cells compared with cells cultured in basal medium (Fig. 4E). Moreover, treatment with PDGF-AA in GLP-1/exendin-4-containing high-glucose medium actually resulted in a 7.52-fold increase in the capability ofβ-like cells to secrete insulin in 25 mM of glucose compared with Ad-NP-infected cells cultured in basal medium (Fig. 4D) and resulted in the highest stimulation index (10.72 ± 0.65) (Fig. 4E). Immunofluorescent staining confirmed downregulation of transferrin in Ad-NP-induced insulin-expressing clusters in GLP-1/exendin-4-containing high-glucose medium (Fig. 4F). At the same time, we performed Western blot analysis to confirm that exendin-4 or PDGF-AA further enhanced reprogramming, as judged by the reduced levels of the hepatic marker, transferrin, in reprogrammed cells generated by Ad-NP (Fig. 4G). These results confirm that activation of PDGFRα by PDGF-AA in GLP-1/exendin-4-containing high-glucose medium could greatly enhance the efficiency of insulin-secreting β-like cell generation during Pdx1 and Ngn3-induced human liver cell reprogramming.

PDGF-AA significantly enhances Pdx1 and Ngn3-induced reprogramming of human liver cells to insulin-secreting β-like cells. Human HepG2/C3A liver cells were infected with Ad-nGFP, Ad-Pdx1, or Ad-Ngn3 or were coinfected with Ad-Pdx1 and Ad-Ngn3 (shown as Ad-NP) for 5 days. (A–C) On day 5 postinfection, the reprogrammed cells were stained with anti-insulin (blue), anti-Pdx1 [red in (A)], or anti-Ngn3 [red in (B)] antibodies. GFP marked adenovirus-infected cells. The results of quantitative analysis are shown in (C). The average percentage of insulin-positive cells in GFP+ reprogrammed cells was determined from five different images. Kruskal–Wallis tests were used for confirming statistical significance, and the data are shown as the mean ± SEM; ***p < 0.001 indicates that the difference between Ad-Pdx1, Ad-Ngn3, and Ad-NP groups was statistically significant. (D) Insulin secretion assay was performed on control or Ad-NP-reprogrammed cells in basal medium containing 30 nM GLP-1, 1 nM exendin-4, 20 nM PDGF-AA, and/or 25 mM glucose. Measurement of insulin secretion release in response to 25 mM glucose was performed in KRB buffer. Insulin secretion values were normalized to total proteins (**p < 0.05, ***p < 0.001). (E) The stimulation index of insulin was calculated as a ratio of glucose-stimulated insulin-secreting levels (25 mM glucose) to the basal insulin-secreting levels (5 mM glucose). Kruskal–Wallis tests were utilized for confirming statistical significance, and the data are shown as the mean ± SEM; ***p < 0.001 indicates that the difference between basal medium and other experimental groups was statistically significant. (F) Immunofluorescence staining for transferrin (red) and insulin (blue) in Ad-GFP, Ad-Pdx1, Ad-Ngn3, or Ad-NP-infected C3A cells. White arrows indicate GFP+ cells with high levels of insulin that have lost transferrin expression. (G) Western blot analysis was performed to determine the expression of transferrin (shown as TFN) in Ad-NP-infected C3A cells cultured in medium containing 30 nM GLP-1, 1 nM exendin-4, 20 nM PDGF-AA, or 25 mM glucose. GAPDH was used as an internal control.
Direct Reprogramming of Primary Hepatocytes to Expandable β-Like Cells
To determine whether Pdx1 and Ngn3 can also trigger direct reprogramming of normal hepatocytes (but not just immortalized liver cell lines) to expandable β-like cells, we isolated primary hepatocytes from adult C57BL/6 mice, infected them with Ad-GFP, Ad-Pdx1, Ad-Ngn3, or Ad-NP, and cultured them in GLP-1/exendin-4-containing high-glucose medium with or without PDGF-AA. At 5 days postinfection, our FACS analysis demonstrated that around 65% of infected hepatocytes were positive for GFP (data not shown). Immunofluorescent staining revealed that 6.75% ± 2.48 of GFP+ (Ad-NP infected) cells expressed insulin, which was 3.86- or 6.31-fold higher than that in the Ad-Pdx1- or Ad-Ngn3-infected groups, respectively (Fig. 5A and B). PDGF-AA treatment was found to significantly increase the amounts of Ad-NP-infected cells that became insulin-producing β-like cells (2.31-fold higher than Ad-NP infection alone) (Fig. 5A and B). Real-time quantitative RT-PCR analysis further showed that, in addition to both insulin1 and insulin2, endogenous pdx1 and ngn3 and other β-cell markers, including mafa and rfx6, were actually induced in Ad-NP-infected hepatocytes. The observation of expression of endogenous pdx1 and ngn3 in PDGF-AA-treated groups suggested that reprogrammed cells are capable of being maintained in a β-like cell state (Fig. 5C).

PDGF-AA facilitates Pdx1 and Ngn3-induced reprogramming of mouse primary hepatocytes to expandable β-like cells. (A–C) Primary hepatocytes isolated from 6-week-old C57BL/6 mice were infected with 3 MOI of Ad-nGFP, Ad-Pdx1-nGFP, Ad-Ngn3-nGFP, or Ad-NP for 3 days. At 24 h postinfection, media were replaced with DMEM/F12 supplemented with 10% FBS, 2 mM L-glutamine, 50 nM 2-ME, 1× NEAA, 25 mM glucose, 1 nM exendin-4, and 30 nM GLP-1, with or without 20 nM PDGF-AA. (A) The reprogrammed cells were stained with anti-insulin antibodies (red) on day 3 postinfection. GFP expression marks adenovirus-infected cells. Nuclei were counterstained with DAPI (blue). Quantitative analysis of the results is shown in (B). White or pink bars indicate the percentage of insulin-positive cells in the culture conditions without or with PDGF-AA treatment, respectively. Kruskal–Wallis tests were used to confirm statistical significance, and the data are shown as the mean ± SEM; ***p < 0.001 indicates statistically significant differences. (C) Real-time quantitative RT-PCR analysis was performed to determine the expression of insulin1, insulin2, endogenous pdx1, endogenous ngn3, mafa, and rfx6 in reprogrammed cells. The level of β-glucuronidase (gusb) was used as a normalized control. Kruskal–Wallis test was used to determine statistical significance. Values are shown as the mean ± SEM. *p < 0.05, **p < 0.01, and ***p < 0.001 indicate statistically significant differences. (D and E) On day 3 postinfection, the reprogrammed cells were labeled with 10 μM EdU for 1 h and then fixed. EdU staining was detected using azide-conjugated Alexa 649 (pink). Insulin expression was detected using anti-insulin antibodies (red). GFP marks adenovirus-infected cells. Nuclei were counterstained with DAPI (blue). The results of quantitative analysis are shown in (E). Kruskal–Wallis tests were performed to confirm statistical significance, and the data shown represent the mean ± SEM; *p < 0.05 or **p < 0.01 indicate statistically significant differences. (F and G) The reprogrammed cells were stained with anti-insulin (red) and anti-PDGFRα (pink) antibodies on day 3 postinfection. The results of quantitative analysis are shown in (G). Kruskal–Wallis tests were used to confirm statistical significance, and the data shown represent the mean ± SEM; ***p < 0.001 indicates statistically significant differences.
To further determine whether the facilitation of reprogramming by PDGF-AA arises from PDGFRα-mediated proliferative induction, we administered EdU at day 3 postinfection to mark reprogrammed cells that enter the S phase under PDGF-AA treatment. As shown in Figure 5D and E, 20.62% of EdU-labeled cells derived from Ad-NP-infected hepatocytes treated with PDGF-AA were found to be positive for insulin (cells indicated by arrows). Indeed, PDGF-AA treatment resulted in a 7.88fold increase in the percentage of insulin-producing cells labeled with EdU compared to Ad-Pdx1 infection alone. Immunofluorescent staining further revealed that PDGF-AA treatment resulted in coexpression of insulin and PDGFRα in 17.76% of Ad-NP-infected cells (Fig. 5F and G), implying that PDGF treatment can induce proliferative expansion of PDGFRα-expressing β-like cells (cells indicated by arrows in Fig. 5F).
Taken together, these above results suggest that combined use of Pdx1 and Ngn3 can trigger reprogramming of adult hepatocytes into PDGFRα-expressing β-like cells. The PDGFRα-expressing β-like cells directly derived from adult hepatocyte reprogramming were shown to be expandable, as activation of PDGFRα by PDGF-AA could promote the entry of β-like cells into the cell cycle.
Transplantation of Hepatocyte-Derived β-Like Cells Can Ameliorate Hyperglycemia in Diabetic Mice
To further confirm that β-like cells derived from adult hepatocyte reprogramming are functional and can be used to treat diabetes, we treated adult C57BL/6 mice with STZ to induce diabetes and then transplanted different amounts of GFP+ reprogrammed cells (PDGF-AA-treated Ad-NP-infected hepatocytes) into the kidney capsule of diabetic mice. We subsequently measured the fasting blood glucose level of four mice in each group at 2, 4, 6, and 8 days after transplantation. Transplantation of 1 × 105 of GFP+ reprogrammed cells was sufficient to ameliorate hyperglycemia (Fig. 6A). Glucose tolerance tests were performed to evaluate if β-like cells derived from adult hepatocyte reprogramming are functional. Mice with GFP+ reprogrammed cell transplants were intraperitoneally injected with glucose (2 mg/g body weight), and then left to fast for 6 h. In contrast to diabetic control mice that were transplanted with Ad-GFP-infected hepatocytes, diabetic mice that were transplanted with 1 × 105 GFP+ reprogrammed cells (PDGF-AA-treated Ad-NP-infected hepatocytes) could efficiently dispose of glucose within 2 h (Fig. 6B). To determine if enhanced glucose tolerance was due to the secretion of insulin byβ-like cells derived from hepatocyte reprogramming in response to glucose injection, we examined the level of plasma insulin of the mice at 120 min after initial injection of glucose. As shown in Figure 6C, higher levels of insulin (113.59 ± 39.67 pmol) were observed in the plasma of mice transplanted with 1 × 105 GFP+ PDGF-AA-treated Ad-NP-infected hepatocytes, compared to that in mice transplanted with GFP+ Ad-NP-infected hepatocytes or Ad-GFP-infected hepatocytes; there was no statistically significant difference between the plasma insulin level of the B6 control mice (170.70 ± 39.64 pmol) and that of mice transplanted with PDGF-AA-treated Ad-NP-infected hepatocytes (Fig. 6C). Immunohistochemical staining revealed that those insulin-positive cell clusters remaining in the graft were positive for Pdx1 and Ngn3 (Fig. 6D). Importantly, higher levels of insulin staining were identified in the plasma of mice transplanted with PDGF-AA-treated reprogrammed cells, which explained why improvement of the blood glucose control of diabetic mice could be achieved (Fig. 6D). These findings indicate thatβ-like cells derived from adult hepatocyte reprogramming can secrete insulin in response to changes in blood glucose levels in vivo, and this successfully relieves hyperglycemia in diabetic mice.

Transplantation of β-like cells derived from hepatocyte reprogramming ameliorates hyperglycemia in diabetic mice. Intraperitoneal administration of streptozotocin (STZ) (200 mg/kg body weight) induced hyperglycemia in C57BL/6 mice (hyperglycemia was defined as a nonfasting blood glucose content higher than 300 mg/dl). On day 4 after STZ injection, GFP+ reprogrammed cells were sorted and transplanted to the kidney capsule of hyperglycemic mice. (A, B) Dose-dependent experiments were performed to determine the minimal amount of reprogrammed β-like cells required to ameliorate hyperglycemia. (A) Blood glucose of transplanted mice was measured every 2 days posttransplantation. (B) Glucose tolerance was examined at day 8 posttransplantation. [Green line: diabetic mice transplanted with 2 × 105 Ad-GFP-infected cells; blue dashed line: diabetic mice transplanted with 2 × 104 PDGF-AA-treated Ad-NP-infected cells; blue line: diabetic mice transplanted with 7 × 104 PDGF-AA-treated Ad-NP-infected cells; red line: diabetic mice transplanted with 1 × 105 PDGF-AA-treated Ad-NP-infected cells; dark blue line: diabetic mice transplanted with 1 × 105 Ad-NP-infected cells; black line in (A): diabetic mice that did not receive transplantation; black line in (B): C57BL/6 control mice that were not injected with STZ (gray bar), shown as B6 ctrl. W/O STZ.] (C) The insulin levels in the plasma of C57BL/6 control mice uninjected with STZ and diabetic C57BL/6 mice that received transplants of either 1 × 105 GFP+ Ad-GFP-infected, Ad-NP-infected, or PDGF-AA-treated Ad-NP-infected cells were determined at day 8 posttransplantation. Kruskal–Wallis tests were used to confirm statistical significance, and the data are shown as the mean ± SEM; *p < 0.05, *** p < 0.001 indicate statistically significant differences. n.s., nonsignificant. (D) At 8 days posttransplantation, kidney grafts were dissected and fixed for histological analysis. Higher magnification images of immunohistochemical staining corresponding to the yellow (Ad-GFP transplanted groups) or green (transplantation with PDGF-AA-treated Ad-NP-induced β-like cell groups) boxed regions are shown below the original images.
Discussion
The current findings demonstrate that combined treatment with Pdx1 and Ngn3 can reprogram hepatocytes intoβ-like cells expressing PDGFRα. Treatment with PDGF-AA in GLP-1/exendin-4-containing medium can facilitate the generation of expandable β-like cells possessing the capacity to secrete insulin in response to stimulatory levels of glucose, enabling their application for treating diabetic mice in vivo. Compared with previous work using Pdx1 alone to reprogram liver cells into insulin-producing cells, the modified strategy developed from the current work not only improved the efficiency of reprogramming hepatocytes into β-like cells but also enhanced the capacities of reprogrammed cells to secrete insulin.
Ngn3 is a key transcription factor involved in regulating β-cell differentiation and the expression of β-cell-specific genes, including pax4, neurod, and insulin 35 . Moreover, reactivation of Ngn3 at secondary transitions during pancreas development has been shown to be accompanied by rapid expansion of endocrine cell lineages 14 , suggesting that Ngn3 not only functions as a regulator triggering expression of β-cell-specific genes but also is involved in regulating the expansion of β-like cells during hepatocyte reprogramming. In addition, Ngn3 has been demonstrated to transactivate downstream genes by indirectly binding to the E-box region on promoters through collaborating with the E47 transcription factors36,37. By analyzing the transcription factor binding sites of the promoter region of pdgfrα genes, at least two E47 and three Pax4-binding regions were identified (data not shown). The present results confirm that pax4 is expressed in reprogrammed cells induced either by Ngn3 alone or by both Ngn3 and Pdx1, implying that Ngn3 is capable of transcriptionally regulating pdgfrα expression, possibly via coordinating with Pax4 and E47 in reprogrammed cells.
Although exendin-4 has been recognized as the agonist of GLP-1 receptors (GLP-1R), our present results show that exendin-4 and GLP-1 treatment induced different effects on reprogrammed cells generated by Ad-NP. We found that while GLP-1 and exendin-4 can both significantly enhance the capacity of reprogrammed cells to secrete insulin in response to stimulatory levels of glucose, only exendin-4 treatment suppressed hepatocyte growth, which would be advantageous for the selective enrichment of reprogrammed β-like cells. Indeed, the amino residues in the central region of either GLP-1 or exendin-4 form a helical structure that interacts with GLP-1R. However, there are sequence differences between GLP-1 and exendin-4, resulting in different binding affinities to the N-terminal domain of GLP-1R 38 . A previous study has shown that exendin-4, but not GLP-1, can promote glucose uptake in adipocytes in response to insulin 39 . Furthermore, exendin-4 has partial cardioprotective effects in GLP-1R knockout mice 40 . These findings suggest that, in addition to triggering a GLP-1R-mediated response, exendin-4 treatment may have beneficial effects on hepatocyte reprogramming that are independent of GLP-1R.
PDGFRα-mediated signaling is known to play an important role in stimulating β-cell replication 17 , and the age-dependent decline of PDGFRα was found to be associated with a reduction in β-cell proliferative capacities 19 . Additionally, activation of PDGFRα has also been shown to elevate cyclin D1/CDK4 complex formation, which results in a further reversion of adult β-cells into a proliferative status. CDK4 has been identified as the critical regulator for inducing proliferation of (i) Pdx1+ progenitors at E11.5 and (ii) Ngn3+ endocrine precursors during the secondary transition of pancreas development 41 , suggesting that CDK4 may play an important role in regulating the cell division of reprogrammed cells. In the present study, we demonstrated that treatment of PDGF-AA not only induced the tyrosine phosphorylation of PDGFRα in reprogrammed cells but also elevated levels of CDK4; however, CDK4-pRb-E2F1 signaling is also known to be involved in regulating insulin secretion and glucose tolerance via regulation of kir6.2 expression 42 . We therefore speculate that PDGFRα-CDK4 signaling plays critical roles not only in regulating expansion of reprogrammed β-like cells but also in facilitating complete hepatocyte reprogramming toward mature β-cells.
Recipients of allogeneic islet transplantation require immunosuppressive medications in order to slow down rejection of transplanted islets 26 . However, under current regimens, immunosuppressive medications are known to be toxic to β-cells, ultimately leading to poor long-term function of the transplanted islets. Our present findings raise the possibility of producing large amounts of β-cells from autologous resources. We confirmed that β-like cells derived from adult hepatocyte reprogramming resemble pancreatic β-cells that express PDGFRα in young mice and can therefore respond to PDGF-mediated expansion signals. Importantly, transplantation of the expandable β-like cells derived from adult hepatocyte reprogramming ameliorated diabetes in recipient mice. These findings support the possibility of utilizing β-like cells derived from autologous hepatocyte reprogramming for the development of cell-based therapeutics for patients with type 1 diabetes mellitus in the future.
Footnotes
Acknowledgments
Experiments and data analysis were performed in part through the use of the confocal microscope at the Scientific Instrument Center of Academia Sinica and with the assistance of Shu-Chen Shen. The authors thank the flow cytometry facility at the Genomics Research Center of Academia Sinica for technical assistance. The authors are grateful to Dr. Douglas Melton (Harvard University Department of Stem Cell and Regenerative Biology) for kindly providing various constructs, including pAd-nGFP, pAd-Pdx1-I-nGFP, and pAd-2B-Ngn3-I-nGFP. This work was supported in part by an Intramural Grant of Academia Sinica and the Ministry of Science and Technology (Grants 99-2314-B-001-002-MY3, 102-2314-B-001-006-MY3, and 104-0210-01-09-02 to C.N.S.). The authors declare no conflicts of interest.