
Abstract
Intra-arterial (IA) delivery of bone marrow-derived mesenchymal stem cells (BM-MSCs) has shown potential as a minimally invasive therapeutic approach for stroke. The aim of the present study was to determine the whole-body biodistribution and clearance of technetium-99m (99mTc)-labeled rat and human BM-MSCs after IA delivery in a rat model of transient middle cerebral artery occlusion (MCAO) using single-photon emission computed tomography (SPECT). Our hypothesis was that xenotransplantation has a major impact on the behavior of cells. Male RccHan: Wistar rats were subjected to sham operation or MCAO. Twenty-four hours after surgery, BM-MSCs (2×106 cells/animal) labeled with 99mTc were infused into the external carotid artery. Whole-body SPECT images were acquired 20 min, 3 h, and 6 h postinjection, after which rats were sacrificed, and organs were collected and weighed for measurement of radioactivity. The results showed that the majority of the cells were located in the brain and especially in the ipsilateral hemisphere immediately after cell infusion both in sham-operated and MCAO rats. This was followed by fast disappearance, particularly in the case of human cells. At the same time, the radioactivity signal increased in the spleen, kidney, and liver, the organs responsible for destroying cells. Further studies are needed to demonstrate whether differential cell behavior has any functional impact.
Keywords
Introduction
Stroke is the most common reason for adult disability (6). At present, the only pharmacological agent approved for treating acute ischemic stroke is tissue plasminogen activator, but due to its narrow therapeutic window of 4.5 h it can be used for only 2–4% of ischemic stroke patients (24). Thus, there is an urgent need for an effective treatment with a wider therapeutic window. Cell-based therapies have emerged as a promising approach and may present noteworthy health benefits for stroke patients (4,27). Cell-based therapies may act at least partly through regenerative mechanisms, such as neurogenesis, angiogenesis, and synaptogenesis (13,22).
A particularly attractive method is intravascular cell delivery because it is a noninvasive and an effective administration route that is suitable for large or multiple infarcts (33). To understand how therapeutic cells work after systemic infusion, it is crucial to follow their whole-body distribution, kinetics, and engraftment patterns within the target tissue. We have previously shown that human umbilical cord blood cells and human embryonic stem cell-derived neural progenitors accumulated primarily in the lungs after intravenous (IV) infusion in rats subjected to middle cerebral artery occlusion (MCAO). Lung accumulation was followed by cellular relocation to the liver and spleen with no major sign of engraftment in the brain (16,24). The lung entrapment pattern seems to depend on cell size and active cell surface adhesion mechanisms (11,16). Fortunately, it is possible to circumvent entrapment by filtration organs through targeting the cells directly to the ischemic brain by using intra-arterial (IA) delivery. IA infusion of 111In-oxine-labeled human bone marrow-derived mesenchymal stem cells (BM-MSCs) has resulted in immediate and efficient cell entrapment in the brain (23). However, again most of the signals disappeared from the brain and relocated to the liver and spleen during the next 24 h.
The aim of the present study was to compare rat and human BM-MSCs in sham-operated and MCAO rats during the first hours after IA delivery. Our hypothesis was that xenografting might have an impact on the engraftment and clearance of cells.
Materials and Methods
Animals
Male RccHan: Wistar rats (weight 246–343 g, age 3–4 months) were purchased from Harlan Laboratories (Horst, The Netherlands) and quarantined for 2 weeks prior to the experiment. The animals were housed individually after surgery in Scantainer (Scanbur, Sollentuna, Sweden) cages (temperature 21 ± 1°C, humidity 50–60%, light period 07:00–19:00 h) with access to food and fresh water available ad libitum. Animal care procedures were carried out according to European Community Council Directives 86/609/EEC guidelines, and all procedures were approved by the National Animal Experiment Board (Hämeenlinna, Finland).
Transient Middle Cerebral Artery Occlusion
Animals were initially anesthetized in a gas chamber (70% N2O and 30% O2) with 3% halothane (Nicholas Piramal Ltd., London, UK) for 2–3 min and then maintained with 0.5–1.5% using a nose mask (Viking Medical Products, Medford Lakes, NJ, USA). Body temperature was maintained at 37°C throughout surgery by means of a heating pad connected to a rectal probe (Harvard Homeothermic Blanket Control Unit, Cambridge, MA, USA). Cerebral ischemia was induced by a filament technique (20). The right common carotid artery was exposed, and a heparinized nylon filament of Ø 0.28 mm was inserted via the stump of the external carotid artery (ECA) into the internal carotid artery (1.8–2.1 cm) to occlude the middle cerebral artery (MCA). After MCA occlusion (60 min), the filament was gently pulled out, and the ECA was closed by electrocoagulation, leaving a long ECA stump for cell transplantation. Sham-operated control animals underwent the same procedure as described above but without filament insertion. Lidocaine (2%; AstraZeneca, Espoo, Finland) was applied after closing the wound. Buprenorfine (0.03 mg/kg; RB Pharmaceuticals Ltd., Slough, Berkshire, UK) was administered for postoperative pain. Completeness of MCAO was evaluated by a modified limb-placing test before cell infusion, which shows a correlation with infarct size (25). Animals with minor behavioral impairment were excluded (n = 6). Exclusion criteria were determined before the experiment. Based on limb-placing scores, MCAO rats were also assigned to equal groups.
Preparation and Characterization of Human and Rat BM-MSCs
Human BM-MSCs were obtained from bone marrow aspirates of healthy volunteer donors (four males) after signed informed consent was gained according to the Declaration of Helsinki. The protocol was approved by the Ethics committee of the Helsinki University Central Hospital (Finland). BM-MSC establishment and characterization has been previously described (23). The cells were cultured in heparinized low-glucose Dulbecco's modified Eagle's medium (Gibco, Life Technologies Ltd., Paisley, UK), supplemented with 10% platelet-rich plasma (Finnish Red Cross Blood Service, Helsinki, Finland), 100 U/ml penicillin, and 100 μg/ml streptomycin (Life Technologies, Thermo Fisher Scientific, Waltham, MA, USA) according to Schallmoser et al. (30). The medium was changed twice weekly, and the cultures were passaged when subconfluent (70–80% confluency) and subcultured at 1 × 103 cells/cm2. Cells were detached with trypsin (TryPLe Express, Life Technologies Ltd.). Cell viability was determined for all the samples by Trypan blue (Stem Cell Technologies, Grenoble, France) exclusion or with a Nucleocounter NC-100 (Chemometec, Allerød, Denmark). Cells [passage 2 (P2)] were cryopreserved in 180 mg/ml human serum albumin (HSA, Albunorm 200 g/L; Octapharma AG, Lachen, Switzerland) and 10% dimethyl sulfoxide (Sigma-Aldrich, Steinheim, Germany) at a cell density of 4 × 106/ml.
Oricell™ male Wistar rat BM-MSCs (Cat. No. RAWMX-01001; Cyagen Biosciences Inc., Sunnyvale, CA, USA) were cultured according to the manufacturer's instructions in OriCell MSC growth medium supplemented with 10% FBS, 1% glutamine, and 1% penicillin-streptomycin (Cat. No. GUXMX-90011; all reagents from Cyagen Biosciences Inc.). The medium was changed twice weekly, and the cultures were passaged when subconfluent (80–90% confluency) and subcultured at 1 × 104 cells/cm2. Rat MSCs (P5) were cryopreserved in the OriCell™ NCR Cryopreservation Medium (Cat. No. NCPF-10001; Cyagen Biosciences Inc.) at a cell density of 4 × 106/ml.
Human MSCs were analyzed for cell surface epitope expression with fluorescent-conjugated antibodies against CD73, CD90 (Stem Cell Technologies), CD105, HLA-ABC, HLA-DR, CD14, CD19, CD34, and CD45 as suggested by the International Society for Cellular Therapy (ISCT) (10). All antibodies were purchased from BD Biosciences (San Jose, CA, USA). Rat MSCs were labeled with antibodies against the markers suggested by the manufacturer, namely CD29, CD90, CD44 (all from AbD Serotec, Raleigh, NC, USA), CD11b, and CD45 (both from BD Biosciences, San Diego, CA, USA). Additionally, CD73 (BD Biosciences) expression was analyzed. Secondary antibody staining with goat anti-mouse IgG (H+L) Alexa Fluor 488 (1:500) (Molecular Probes, Life Technologies Corp., Eugene, OR, USA) was done for anti-CD73. The cells were labeled with 2–10 μ1 of the antibodies/1 × 105 cells for 30 min at 4°C and run with a FACSAria flow cytometer (BD, Franklin Lakes, NJ, USA). The results were analyzed with FlowJo software (version 7.6.1; TreeStar Inc., Ashland, OR, USA). Adequate isotype control antibodies (BD Biosciences) were also used.
The cell size was measured in P2 for human MSCs and P5 for rat MSCs, that is, the same passage as used for transplantation. The analysis was done with Beckman Coulter Counter Z1 (Beckman Coulter Inc., Fullerton, CA, USA) according to the manufacturer's instructions. Briefly, the counter was calibrated with nominal latex beads (Coulter CC Size standard L10; Beckman Coulter Inc.) and for measurement, 100 μl of cell suspension, harvested at subconfluency, was diluted with 9.9 ml of phosphate-buffered saline (PBS) (all reagents from Sigma-Aldrich) containing 0.02% sodium azide (Sigma-Aldrich). The aperture range used was from 7 to 22 μm.
Labeling of Cells with 99mTc-HMPAO
For cell labeling, the cryopreserved cells were thawed in a water bath at 37°C (16). The cells were then decanted into the thawing medium containing α-MEM (Life Technologies Ltd.) and 10% HSA. Cells were centrifuged, and medium was removed. 99mTc-pertechnetate (2.5 GBq in 3 ml of 0.9% saline) was purchased from MAP Medical Technologies (Tikkakoski, Finland), and a HMPAO kit (Ceretec™) was obtained from GE Healthcare Ltd. (Amersham Place, Little Chalfont, Buckinghamshire, UK). Freshly eluted 99mTc-pertechnetate (2.5 ml, around 1.7 GBq) in saline solution was administered to a Ceretec™ vial and mixed thoroughly, after which 1 ml of freshly prepared 99mTc-HMPAO complex was added to 1 ml of cell suspension (approximately 10 × 106 cells). Cells were incubated for 15 min at room temperature and mixed gently throughout the labeling reaction. Cells were centrifuged at 500 × g for 5 min, and the supernatant containing unbound free 99mTc-HMPAO was removed. Cells were washed with 10 ml of PBS, centrifuged at 500 × g for 5 min, and the final radioactivity of 99mTc-labeled cells was measured with a dose calibrator (CRC-25R; Capintec Inc., Ramsey, NJ, USA). Cells were resuspended in cell culture media and divided into 500-μl aliquots.
Cell Infusion
For cell infusion, rats were anesthetized with isoflurane (Orion Pharma Animal Health, Turku, Finland) (4% for induction, 1.2% for maintenance), and the ECA stump was exposed. 99mTc-HMPAO-labeled rat or human BM-MSCs (2 × 106 cells/animal) in 500 μl saline were slowly infused over a period of 2 min through the stump of the ECA, while the blood flow was maintained in the internal carotid artery. The ECA was closed by electrocoagulation, the wound was closed, and the animal was prepared for single-photon emission computed tomography-computed tomography (SPECT-CT) imaging.
SPECT-CT Imaging
SPECT-CT imaging was performed with a four-headed small animal scanner, NanoSPECT/CT (Bioscan Inc., Washington, DC, USA), outfitted with multipinhole collimators. The body temperature was maintained throughout the study by using a heated animal bed (Equipement Vétérinaire, Minerve, France). Whole-body SPECT images were acquired 20 min, 3 h, and 6 h postinjection in 20 projections using a time per projection of 35 s resulting in a total acquisition time of 23 min. CT imaging was carried out with a 45-kVp tube voltage with 180 projections. SPECT images were reconstructed with HiSPECT NG software (Scivis GmbH, Germany) and fused with CT datasets by using InVivoScope software (Bioscan Inc., Washington DC, USA).
Six hours after cell delivery, the rats were sacrificed, and organs were collected and stored in RIA tubes. Tissue samples were weighed, and their radioactivity was measured with a gamma counter (RiaCalc WIZ, Wallac 1480 WIZARD® 3; Wallac Oy, Turku, Finland).
Quantification
Reconstructed SPECT images were reoriented and analyzed with the InVivoScope software by using CT data as a reference. Briefly, a three-dimensional volume of interest (VOI) was defined around the brain, lung, liver, kidney, and bladder regions. The same VOI was used for all subsequent analyses of corresponding organs. The radioactivity in each VOI was calculated and corrected to the time of injection. Results are presented as a percentage of the injected dose.
Histology
Additional sham-operated and MCAO rats were perfused for histological analysis 20 min after infusion of human BM-MSCs (baseline). Vibratome sections were stained with an antibody specific for human nuclei (MAB1281, 1:1,000; Millipore) and for endothelial cells (RECA-1, 1:1,000; AbD Serotec) to investigate possible perivascular localization of cells (23).
Statistics
The statistical analyses were done with SPSS software (IBM, Armonk, NY, USA), repeated measures ANOVA (for SPECT data), and two-way ANOVA (for radioactivity data in different organs) with cell type and sham/ MCAO operation as covariates. There were no statistically significant differences between sham-operated and MCAO rats in any measures, which allows us to combine these groups and compare rat versus human cells. Six rats received rat cells, and six rats received human cells. Differences in SPECT data and radioactivity counts between groups at various time points were analyzed by Mann– Whitney U-tests.
Results
Characterization of Human and Rat BM-MSCs
Both human and rat BM-MSCs showed the typical spindle-shaped morphology of MSCs before detachment. In the flow cytometric analysis, a set of markers defined by the ISCT was used. The human BM-MSCs were positive for MSC markers CD73, CD90, and CD105 (all >99% positive) and negative for hematopoietic lineage markers CD14, CD19, CD34, and CD45 (Fig. 1). In addition, human BM-MSCs expressed HLA-ABC (>99%) and HLA-DR (36.3%). Rat BM-MSCs were positive for CD29 (100%), CD44 (100%), CD73 (62.1%), and CD90 (99.8%), and negative for CD11b (1.3% positive) and CD45 (Fig. 1). Based on Coulter Counter measurements, the average cell size was 15–17 μm for rat and 17–18 μm for human cells.

Immunophenotypic profile of human (P2) and rat (P5) BM-MSCs used in the study. Flow cytometric analysis showed the expression of CD73, CD90, CD105 HLA-ABC, and HLA-DR (dark gray histograms) on the human MSC surface compared to respective isotype controls (light gray histograms). Human BM-MSCs were negative for the expression of CD14, CD19, CD34, and CD45. Rat MSCs expressed CD29, CD44, CD73, and CD90, but not CD11b or CD45 (dark gray histograms) on the cell surface. Respective isotype controls are also shown (light gray histograms).
Efficient Accumulation of Both Human and Rat BM-MSCs in the Brain
99mTc-HMPAO-labeled BM-MSCs were infused into the ECA 24 h after sham operation or MCAO. Representative whole-body biodistributions of rat and human BM-MSCs are shown in Figure 2 (see also supplemental figure: https://docs.google.com/drawings/d/1krdKHP_Gfu5oHqOpZpxT6SfUvdtCLveS0dQBa7wf01U/edit?usp=sharing). The majority of both the human and the rat cells were located in the brain and on the hemisphere ipsilateral to infusion shortly after cell delivery (baseline).

Representative SPECT-CT images show homing of rat (A, B) and human (C, D) BM-MSCs in the brain after IA infusion. Images on the left (A, C) are for MCAO rats, and on the right (B, D), images are for sham-operated rats. The images are decay corrected and scaled to baseline.
RECA-1 and MAB1281 double immunohistochemistry at baseline showed that human BM-MSCs were located in the vascular space, and there were no signs of cell aggregation (Fig. 3). There was no statistically significant difference in the initial localization within the brain between the human and rat cells. High signal, although variable, was observed in the bladder and intestines due to the accumulation of free tracer.

Histological staining of human nuclei (MAB1281) and blood vessels (RECA-1) in sham-operated (A) and MCAO (B) rats. Perfusion of animals was made 20 min after cell infusion (baseline). Human BM-MSCs were located within the brain capillaries. Scale bar: 20 μm.
Differential Clearance of BM-MSCs From the Brain
Semiquantitative analysis of different organs was used to assess cell behavior after initial accumulation in the brain. Repeated measures ANOVA showed no difference between sham-operated and MCAO rats (p = 0.817), and thus these groups were pooled. When rat and human cells were compared, ANOVA for repeated measures showed a significant overall group effect (p < 0.01) for a signal in the brain during a 6-h follow-up. A significant group × time interaction (p < 0.01) indicated differential clearance of rat and human cells from the brain. Group comparisons between MCAO rats during the follow-up showed that the signal for human BM-MSCs was lower than for rat cells (p < 0.01) at 6 h after cell delivery (Fig. 4A).
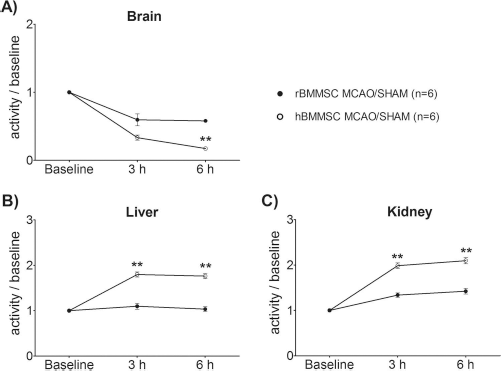
Semiquantitative analysis of activity in the brain (A), liver (B), and kidney (C) after infusion of rat or human BM-MSCs. Data are decay corrected, scaled to baseline, and presented as mean ± SEM. Number of rats was six in both groups. Statistical significance: **p < 0.01 (rat vs. human cells).
There was no difference in liver (p = 0.376) or spleen (p = 0.124) signal between sham-operated and MCAO rats. ANOVA for repeated measures showed a significant group effect (p < 0.001) for a signal in the liver. In addition, a significant group × time interaction (p < 0.001) indicated differential relocation for rat and human BM-MSCs to the liver. Group comparisons during the follow-up showed that the signal for human BM-MSCs was higher (p < 0.01) at 3 and 6 h after cell delivery (Fig. 4B). Similar signal relocation was found in the kidneys. ANOVA for repeated measures showed a significant group effect (p < 0.001) and group × time interaction (p < 0.001) (Fig. 4C). Differences between human cells and rat cells were observed at 3 h and 6 h after cell delivery (p < 0.01) (Fig. 4C).
Radioactivity in Different Organs 6 h After Cell Delivery
Organs were collected for radioactivity measurements at 6 h after cell delivery (Fig. 4). There were no statistically significant differences between sham-operated and MCAO rats, and thus data from these groups were combined. Consistent with SPECT images (Figs. 2, 4), radioactivity for human BM-MSCs was significantly higher in the ipsilateral hemisphere (p < 0.05), spleen (p < 0.001), kidneys (p < 0.001), and liver (p < 0.001) (Fig. 5).

Radioactivity counts as a percentage of the injected dose per organ are shown 6 h after cell infusion of rat or human BM-MSCs. The number of rats was six in both groups. Data are presented as mean ± SEM. Statistical significance: *p < 0.05; **p < 0.01; ***p < 0.001 (rat vs. human cells).
Discussion
Determining the whole-body biodistribution of infused cells is critical for the development of intravascular cell therapies. In this study, we showed that SPECT imaging provides fast and effective information about cell biodistribution and that xenotransplantation has a major effect on cell clearance from the brain in a rat model of stroke, which should be taken into careful consideration when planning safety and efficacy studies for cell therapy products.
There have been significant improvements in small animal SPECT systems over recent years; for example, the imaging resolution is now less than 1 mm, and tracer concentrations in the picomolar or nanomolar range can be detected (26). SPECT is also a translational imaging modality, and the tracers used in animal studies are used in clinical practice. Still, only a few studies have taken advantage of SPECT for cell tracking in stroke models, possibly due to safety issues (2,17,21,23). Cells are affected by low-energy high-linear energy transfer Auger electron emitting 99mTc, although not to the extent of that in the case of radioactive 111In (26). According to Haefliger et al. (12), the radioactivity of 99mTc below the level of 466 Bq/cell had almost no effect on cell proliferation. In our study, the radioactivity of 99mTc was 0.3–14 Bq/cell, which was not considered to be radiotoxic. However, cell viability testing is essential particularly if long-term functional outcome will be studied.
Here we showed efficient entrapment of both rat and human BM-MSCs to the brain shortly after IA infusion by using SPECT imaging. No difference between sham-operated and MCAO rats was observed, indicating that inflammatory signals present in the brain at 24 h after MCAO do not explain the results. The described cell behavior is intriguingly similar to the cell entrapment seen in the lungs following IV cell administration (11). Both cell size and adhesion molecule expression seem to contribute to pulmonary entrapment, which is considered to be the main obstacle impairing efficient cell homing to target tissue. We cannot exclude the possibility that IA infused cells cause blockage of small vessels and brain tissue damage as suggested by Janowski et al. (15). However, this is not supported by histological staining of human BM-MSCs shortly after IA infusion. Only a few scattered cells were observed within vasculature or the brain parenchyma at 24 h after infusion in our previous study (23).
To our surprise, the clearance of cells from the brain tissue was extremely fast. After 6 h, only 58% of the signal related to rat cells was left. In addition, human BM-MSCs disappeared much faster than rat cells. The size of the rat and human BM-MSCs is the same, and thus the cell size does not seem to determine the differential clearance. Interestingly, Brenneman et al. (7) showed a similar disappearance of autologous BM-mononuclear cells in a permanent MCAO rat model. The remaining cells were TUNEL positive 3 h after infusion, indicating a rapid necrotic/apoptotic process. Rosenblum et al. (28) showed that, in a mouse stroke model, therapeutic cells were more likely to be phagocytosed when infused within the first 24 h after hypoxia–ischemia, which may also contribute toward graft cell disappearance. Cells (dead and alive) are eliminated from the body in the liver and spleen, and thus high signal in these organs could reflect the elimination process (23). Consistent with this, detachment of the 99mTc due to cell death results in high signal in the bladder and intestine.
One explanation for the different clearance of human cells compared to rat BM-MSCs from the brain could be that human cells induce an immunological response much faster than rat allogeneic cells, which could lead to rapid rejection of the cells (5). Current studies lack information on the immunological responses to cell administration in the rat stroke model, and the existing data are inconsistent. Human MSC engraftment and survival have been shown in the brains of rats with partially impaired immune systems (3). Comparisons of human and rat allogeneic BM-MSCs showed that both cells elicited only a weak immune response after transplantation, and cells survived for at least 3 months in the host brain (29). Similar findings have been shown in xenotransplantation of mouse MSCs to the intact and ischemic rat brain, although immunosuppression could improve graft survival (14). Contradictory findings suggest that even allogeneic BM-MSCs could be rejected in the healthy rat brain (9).
Whether BM-MSCs evoke immunological reactions or not, functional recovery using human BM-MSCs in rat brain ischemia models has been shown in several studies (1,8,19,34,35). According to Yang et al. (34), systemic administration of either rat or human MSCs was equally beneficial in enhancing motor recovery in MCAO rats, although very few MSCs were found in the brain. In addition, a recent review gathered data from 94 reports to show that in 88 cases MSCs could function in several different cross-species models, indicating that these cells might overcome xenobarriers (18). One explanation could be that MSCs exert their therapeutic effects through a “hit and die” mechanism, that is, rapid migration to the damaged organ and subsequent clearance accompanied with the release of therapeutic molecules. In that case even xenogeneic cells could have enough time to be therapeutic, although they are rather rapidly rejected. The fact that even allogeneic MSCs will be eventually rejected can, in fact, limit the long-term risks of MSC therapy (31,32).
In conclusion, our study suggests that xenografting has an impact on cell clearance from the brain and faster relocation of human MSCs compared to rat MSCs to the liver and spleen in animals. This decreases the bioavailability of cells in the target tissue as well as their therapeutic effect. Overall, the complexity of immunological processes, differences of immune systems between species, and uncertainty behind the exact mechanisms of how MSCs influence these processes makes it difficult to make broad conclusions. Thus, controlled comparative studies of the therapeutic effect and immunological responses between human versus allogeneic MSCs in experimental stroke models are urgently needed. Homologous animal models may provide the most relevant system for proof-of-concept, while xenogeneic models are essential for safety studies. The existing limitations require careful interpretation of data.
Footnotes
Acknowledgments
We would like to thank Sirkka Hirschovits-Gerz, Susanna Räsänen, Lotta Sankkila, and Sofia Oja for their technical assistance with animal care and cell culture. Eero Hippeläinen is acknowledged for professional help with SPECT images. This research work was supported by the Health Research Council of the Academy of Finland and by a grant of the Academy of Finland (263171) in the framework of the ERA-NET NEURON. The authors declare no conflicts of interest.