
Abstract
Human amnion epithelial cells (hAECs) have been shown to modulate inflammation and restore normal lung structure and respiratory function following bleomycin challenge in immune-competent mice. These effects are exerted despite a lack of significant engraftment of hAECs, suggesting that immunomodulatory effect mechanisms are at play. In this study, using the bleomycin model of injury, we explored the interactions between hAECs and macrophages. We administered 4 million hAECs intraperitoneally to C57Bl6 mice 24 h following a bleomycin challenge. Using FACS analysis and qPCR, we showed that hAEC administration significantly reduced macrophage infiltration into the lungs and that the majority of the pulmonary macrophages were of the M2 phenotype. Using bone marrow-derived macrophages, we then showed that hAEC-conditioned media could alter macrophage polarization, migration, and phagocytosis, without affecting macrophage survival or proliferation in vitro. This study provides the first evidence that hAECs directly influence macrophage behavior in a proreparative manner and suggests that hAECs are able to mediate these effects independently of other immune cell types.
Introduction
The human placenta, umbilical cord, amniotic fluid, and fetal membranes have been shown to be rich sources of multipotent stem cells and stem cell-like cells (3,4,9,14,15,19–21,23,31) suitable for regenerative medicine application. Indeed, a variety of placenta-derived cells have been used in diverse tissues for repair and regeneration, including replacement of insulin-producing islets cells (10,13), reduction of liver fibrosis (19), improvement of ventricular function following myocardial infarct (6), functional recovery following stroke (17), and general neuroprotection (11). We, and others, have shown that amnion cells, whether derived from amniotic fluid or amnion membrane, can differentiate into alveolar (lung) cells and can mitigate injury and facilitate repair in varied models of neonatal and adult lung injury (3,4,23,33). While the initial studies of amnion cells in lung injury, undertaken in immune-competent mice, suggested that the cells effected repair via integration and differentiation into lung cells (4,21), more recent work has cast doubt on in vivo differentiation as the primary mechanism of repair (8,23). Instead, it appears that mitigation of lung injury and scarring is more related to modulation of the host inflammatory response to injury. Specifically, amnion epithelial cells or mixed amniotic fluid cells reduce the pulmonary inflammatory infiltrate following injury (3,21,22). In particular, the ability of hAECs to prevent acute lung injury or effect repair appears dependent on modulation of the host macrophage response (23).
Specifically, we have shown that hAECs were unable to abrogate lung injury in surfactant protein C knockout (Sftpc-/-) mice (22). Sftpc-/- mice have macrophages with impaired phagocytic ability (7), suggesting that normal macrophage function and responsiveness may be central to the reparative effects of hAECs. In particular, we wondered whether the polarization of macrophages from an M1 to an M2 phenotype was a crucial step by which hAECs could reduce pulmonary inflammation and fibrosis following bleomycin insult. The concept of M1 and M2 macrophage heterogeneity, where macrophages polarize toward different phenotypes in response to changes in stimuli, arose from the observations of Stein and colleagues (30). Classically activated (M1) macrophages exhibit a Th1-like phenotype. They promote inflammation, degradation of the extracellular matrix, and apoptosis. Alternatively activated (M2) macrophages display a Th2-like phenotype. They promote extracellular matrix deposition and angiogenesis, and are proreparative. In this current study, to gain further insights into the mechanisms by which hAECs may effect tissue repair, we set out to ask whether hAECs could directly alter macrophage phenotype and function.
Materials and Methods
Animals
All experimental procedures were approved by Monash University Animal Ethics Committee and were conducted in accordance with the guidelines stipulated by the Australian Code of Practice for the Care and Use of Animals for Scientific Purposes (1). Six- to eight-week-old female C57Bl6 mice sourced from Monash Animal Services (Monash University, Clayton, Victoria, Australia) were administered intranasal 0.3 U bleomycin (Blenoxane, Bristol Myer Squibb, New York City, NY, USA). Each mouse was administered either 4 million hAECs in 0.2 ml saline, or saline alone, via intraperitoneal injection 24 h following bleomycin insult (n = 6 per group). Isolation of hAECs suitable for clinical use was performed as described previously in accordance with guidelines and approval from the Southern Health Human Research Ethics Committee (24). Of a total of 15 amnions collected, 60% were from female fetuses and the remainder 40% from male fetuses.
Tissue Collection
Mice were culled at days 1, 3, 5, and 7 (after bleomycin administration) where the bronchoalveolar lavage fluid and lungs were collected from each animal for analyses. Lungs were minced using a McIlwain tissue chopper (Campden Instruments, Lafayette, IN, USA). Lung tissues were then digested in Waymouths medium (Invitrogen, Rowville, VIC, Australia) containing 25 mg/ml collagenase IA (101 03578001, Roche Applied Science, Mannheim, Germany), 2.5 mg/ml DNase I (AMPD1, Sigma-Aldrich, St. Louis, MO, USA), 25 mM HEPES (H3375, Sigma-Aldrich), and 10% heat-inactivated chicken serum (16110-082, Invitrogen) for 15 min at 37°C. Cells were stained with the following antibodies prior to fluorescence-activated cell sorting (FACS) analysis; cluster of differentiation 45–violet 450 (CD45–V450; 1:150, 560541, BD Bioscience, North Ryde, NSW, Australia), mouse homolog of epidermal growth factor (EGF)-like module-containing mucin-like hormone receptor-like 1-fluroescein isothiocyanate (F4/80-FITC; 1:100, 60343, AbCam, Cambridge, MA, USA), CD86-phycoerythrin cyanine 7 (PE Cy-7; 1:200, 560501, BD Bioscience), and CD206-Alexa Fluor 647 (1:200, 12310, Australian Biosearch, Karrinyup, WA, Australia). Representative FACS plots for isotype controls are shown in Figure 1. A separate lobe was collected for histological analyses. Briefly, the right lung lobes were ligated at the right mainstream bronchus, and the left lung lobe was perfused with 4% paraformaldehyde (PFA; Sigma-Aldrich). The left lung was then excised, immersed in 4% PFA for 24 h, and processed for histology by paraffin embedding.
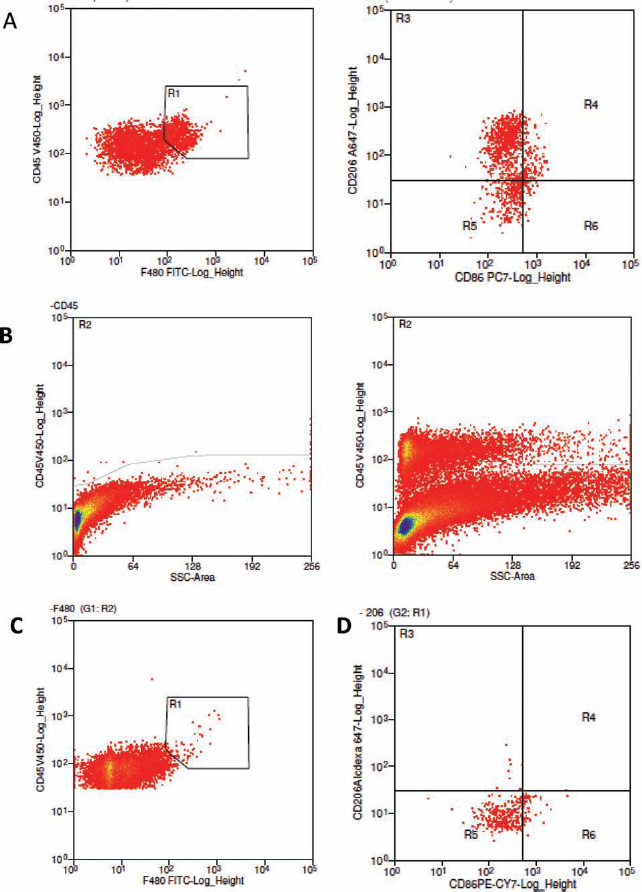
FACS analysis used to detect if hAECs mitigate recruitment and polarize macrophages from M1 toward M2 phenotype in adult C57BL/6 mice given hAECs following bleomycin exposure. (A) Representative fluorescence-activated cell sorting (FACS) plots for cell surface markers cluster of differentiation 45 positive (CD45+), F4/80+, CD206+, and CD86+ from the lungs of adult mice. Macrophages were gated using the criteria of CD45+ and F4/80+ dual staining. Macrophages were then gated for surface markers CD206 and CD86. Isotype controls were used for determining macrophage population and phenotype from total pulmonary leukocyte population. Representative FACS plot for CD45 and its isotype control for IgG2aκ in fluorochrome V450 (B). Isotype control for pan macrophage marker F4/80 for IgG2aκ in fluorescein isothiocyanate (FITC) (C). Isotype controls for surface markers of macrophage phenotypes M1 (CD86) and M2 (CD206) using IgG2aκ isotype controls in phycoerythrin-cyanine 7 (PE-Cy7), and Alexa-Fluor 647, respectively (D). hAECs, human amnion epithelial cells; SSC, side scatter.
Isolation of Alveolar Macrophages
Alveolar macrophages were collected from broncho-alveolar lavage fluid obtained from freshly culled mice by repeated washes with sterile saline and cultured in Dulbecco's modified Eagle's medium (DMEM) high glucose (11965-084, Invitrogen) with 10% (v/v) fetal bovine serum (FBS; 16000-036, Invitrogen) at 37°C for 2 h. Adherent cells were confirmed to be macrophages by FACS analysis where >95% were CD45+ F4/80+.
Collection of hAEC-Conditioned Media
hAECs (0.5 × 106) were isolated as previously described in detail (24) and seeded into a T175 flask (353028, BD Falcon®, Bedford MA, USA) with 20 ml DMEM/F12 (11320033, Invitrogen) supplemented with 10% heat-inactivated FBS (16000-036, Invitrogen) and penicillin– streptomycin (15070-063, Invitrogen). Cells were cultured at 37°C in 5% CO2 for 4 days. Conditioned media was then collected and filtered through a 0.2-μm pore filter (16532, Sartorius Stedim Biotech, Goettingen, Germany) prior to use.
Macrophage Culture and In Vitro Treatment Groups
Bone marrow-derived macrophages were isolated from C57Bl/6 mice and cultured in media as described above. Macrophages at passage four were plated at a density of 0.5 × 106 per well in a six-well plate (353502, 9.6 cm2; BD Falcon, North Ryde, NSW, Australia). Experimental groups consisted of macrophages cultured in control media alone (DMEM/F12 with FBS and antibiotics), hAEC-conditioned media, lipopolysaccharide (LPS), and LPS with subsequent culture in hAEC-conditioned media. Macrophages were exposed to an inflammatory stimulus by culturing macrophages in media containing 10 μg/ml LPS (L2630, Sigma-Aldrich) for 48 h prior to experimental treatments.
Gene Expression Studies
Isolation of RNA and subsequent cDNA synthesis were performed on the alveolar macrophages using the TaqMan® Gene Expression Cells-to-CT kit, according to manufacturer's instructions (4399002, Applied Biosystems, Scoresby, VIC, Australia). Subsequent qPCRs were performed using the following mouse primers: interleukin-1β (IL-1β) forward: CCC AAG CAA TAC CCA AAG AA, reverse: CAT CAG AGG CAA GGA GGA AA; nitric oxide synthase 2 (NOS-2) forward: CAG CTG GGC TGT ACA AAC CTT, reverse: CAT TGG AAG TGA AGC GTT TCG; chemokine (C-C motif) ligand 4 (CCL4) forward: GAA ACA GCA GGA AGT GGG, reverse: CAT GAA GCT CTG CGT GTC TG; arginase-1 (arg-1) forward: CAA GAC AGG GCT CCT TTC AG, reverse: GTA GTC AGT CCC TGG CTT ATG G; found in inflammatory zone 1/resistin-like α (FIZZ-1) forward: CCC AGG ATG CCA ACT TTG A, reverse: AGG AGG CCC ATC TGT TCA TAG; chitinase 3-like 3/T-lymphocyte-derived eosinophil chemotactic factor (ECF-L) (Ym-1) forward: GGG CAT ACC TTT ATC CTG AG, reverse: CCA CTG AAG TCA TCC ATG TC; 18S forward: GTA ACC CGT TGA ACC CCA TT, reverse: CCA TCC AAT CGG TAG TAG CG. Detailed cycling parameters are available from the authors upon request.
Macrophage Migration Assay
Macrophage migration was assessed as briefly described below. Chemotactic assays were performed using 6.5-mm diameter, 3.0-μm pore size, 24-well cell culture inserts obtained from BD Bioscience. The lower chamber was filled with Roswell Park Memorial Institute (RPMI)-1640 (11875-093, Gibco) containing 600 ng/ml of recombinant mouse macrophage inflammatory protein (rMIP)-2 (R&D Systems, USA). The upper chamber contained macrophages (1 × 105), with or without hAEC-conditioned media, with or without prior LPS treatment, and the setup was incubated at 37°C for 4 h. Following incubation, the cell culture insert membranes were stained with Diff-Quick (Lab Aids, Narrabeen, NSW, Australia). The membranes were then removed and placed on microscope slides so that the number of migrated cells could be determined using light microscopy.
Phagocytosis Assay
Macrophages were plated in six-well flat-bottom culture plates (353502, 9.6 cm2; BD Falcon, Australia) at a density of 5 × 105 per well for 48 h prior to stimulating for 12, 24, 48, or 72 h with or without LPS ± hAEC-conditioned media. Staphylococcus aureus particles labeled with pHRodo (Dr Aaron Irving, Monash Micro Imaging, Monash University) were added to each well (10 μg/ml). Phagocytic activity was analyzed by FACS after 1-h incubation at 37°C. Negative control cells were prechilled in an ice/water (50:50) bath in the dark for 10 min to inhibit membrane movement prior to FACS. Only cells that have phagocytosed pHRodo-labeled S. aureus were fluorescent and stained positive on FACS.
MTS Assay
Macrophage viability and proliferation was assessed using the Promega MTS assay (G5421; Promega, Madison, WI, USA), according to the manufacturer's instructions. Briefly, 5,000 macrophages were plated into each well of a 96-well plate (BD Falcon). The cells were allowed to attach overnight before treatment. For each treatment and timepoint, 20 ml of MTS solution was added, and the cells were incubated for 4 h. Optical density was measured at 490 nm with the Benchmark Plus Microplate Spectrophotometer (Bio-Rad, Hercules, CA, USA).
Immunofluorescence for Macrophage Markers CD86 (M1) and CD206 (M2)
To assess macrophage phenotypic markers CD86 and CD206, paraffin-embedded sections were dewaxed and rehydrated. Sections were incubated in sudan black (S0395, Sigma Aldrich) to remove auto fluorescence. Citrate (10 mM sodium citrate, 0.05% Tween 20; S-4641, P1379, Sigma Aldrich) antigen-retrieval step was performed followed by blocking with serum-free protein blocker (X0909, Dako, Glostrup, Denmark). Sections were then incubated with primary F4/80 (1:100, MCA497R, AbD Serotec, Düsseldorf, Germany), CD86 (1:250, NB110-55488, Novus Biologicals, USA), and CD206 (1:25, MCA2235, AbD Serotec) at room temperature for 1 h. CD206 was labeled on a serial section. Tissue sections were incubated in fluorescently labeled secondary antibodies (goat anti-rat IgG Alexa-Fluor® 488; A11006, Invitrogen, Carlsbad, CA, USA and donkey anti-rabbit IgG Alexa Fluor® 568; A10042, Life Technologies, Grand Island, NY, USA) for 1 h followed by a nuclei stain with 4′,6-diamidino-2-phenylindole (DAPI; 1:1000 D9542; Sigma-Aldrich). Images for the serial sections were then acquired using a Nikon C1 confocal laser scanning microscope (Nikon, Tokyo, Japan), aligned with the Autoaligner software (v. 6; Bitplane, Zurich Switzerland), and analyzed using the image analysis software Image J (NIH, Bethesda, MD, USA).
Data Analysis and Statistics
Data were expressed in each experimental groups as mean ± SEM and statistical significance determined by statistical analysis software (GraphPad Prism, GraphPad Software Inc., San Diego, CA, USA). In vivo, FACS and qPCR were analyzed using a one-way ANOVA accompanied by the Bonferroni post hoc test. In vitro FACS, qPCR, and MTS assays were analyzed using the repeated measures analysis of variance (ANOVA) accompanied by the Bonferroni post hoc test. Finally, the in vitro phagocytic and migration assays were analyzed using the Students' t test for paired samples. Statistical significance was accorded when p < 0.05.
Results
hAECs Reduced Macrophage Infiltration and Altered Predominant Phenotype
The total number of macrophages in the lung was determined by FACS analysis for a population of cells that stained double positive for F4/80 and CD45. This population was then gated and further identified as being either CD86+ (M1) or CD206+ (M2) (Fig. 1A). Bleomycin administration significantly increased total macrophage number in the lung at day 7, an effect prevented by the administration of hAECs (4.1% ± 0.4% vs. 0.5% ± 0.1%, p < 0.005) (Fig. 2A). When further analyzed for CD86 and CD206 expression and markers of M1 (NOS-2, IL-1β, and CCL4) or M2 (arg-1, Ym-1, and FIZZ-1), we observed that in bleomycin + saline-treated animals, the infiltrating macrophages were predominantly M1, while in bleomycin + hAEC-treated animals, the predominant macrophage phenotype was M2 as determined by FACS analysis (Fig. 2B, C), immunohistochemistry (Fig. 2D, E), and qPCR for genes specific to each macrophage phenotype (Table 1). Representative confocal images of CD86 and CD206 localization in the lungs of bleomycin + saline and bleomycin + hAEC-treated animals are shown in Figure 2D, E.

hAECs mitigate recruitment and polarize macrophages from M1 toward M2 phenotype in adult C57BL/6 mice given hAECs following bleomycin exposure. (A) Administration of hAECs following bleomycin exposure significantly reduced the percentage of infiltrating macrophages (CD45+/F4/80+) in the lungs of adult mice at day 7. (B) Percentage of CD86+ expressing macrophages was significantly decreased at day 7 in the lungs of mice given hAECs. (C) CD206+ expressing macrophages significantly increased from day 5 to 7 in the lungs of mice given hAECs. Data are represented as mean ± SEM. *p ≤ 0.05, **p ≤ 0.01 is significantly different from control. Representative images showing immunohistochemistry with fluorescent-labeled antibodies for F4/80 (green), CD86 (red), CD206 (white), and nuclei (blue) from bleomycin + saline (D) and bleomycin + hAECs (E) mouse lung sections, respectively. Asterisks indicate colocalization of F480+CD86+ in yellow, and arrows indicate F480+CD206+ in green/white.
Proinflammatory M1 and Proreparative M2 Gene Expression Relative to Housekeeping Gene, 18S of Alveolar Macrophages Isolated From Adult C57BL/6 Mice Given Human Amnion Epithelial Cells (hAECs) or Saline Following Bleomycin Exposure

CCL-4, chemokine ligand 4; IL-1β, interleukin-1β; NOS-2, nitric oxide synthase-2; Arg-1, arginase 1; FIZZ-1, resistin-like protein α; Ym-1, chitinase 3-like protein 3.
hAEC Altered Macrophage Phenotype and Behavior In Vitro
To assess the effects of hAECs on macrophage polarization and recruitment in the absence of other cell types, we performed a series of in vitro experiments, culturing macrophages in hAEC-conditioned media. Either primary macrophages from C57Bl6 mice or macrophages exposed to LPS were treated with hAEC-conditioned media and assessed for CD86 and CD206 by FACS and qPCR. Regardless of whether the macrophages were stimulated with LPS, exposure to hAEC-conditioned media significantly reduced the proportion of CD86+ cells and significantly increased the proportion of CD206+ cells (Fig. 3A, B). This outcome was confirmed by qPCR for genes specific to the M1 and M2 macrophage phenotype (Table 2), where we found a modest increase in M2-specific genes in macrophages exposed to LPS and a further increase of these M2 markers when macrophages were cultured in hAEC-conditioned media compared to LPS-exposed macrophages.

Soluble factors from hAEC-polarized macrophages from proinflammatory M1 toward proreparative M2 phenotype in vitro. Macrophages ± lipopolysaccharide (LPS) exposure were cultured in hAEC-conditioned media for 72 h before FACS analysis. (A) hAEC-conditioned media significantly reduced proportion of CD86+ macrophages. (B) Conversely, hAEC-conditioned media significantly increased the proportion of CD206+ macrophages. Data are represented as mean ± SEM. *p ≤ 0.05, ***p ≤ 0.001, ****p ≤ 0.0001. (C) Soluble factors from hAECs do not increase the proliferative capacity of macrophages. Macrophages were cultured with hAEC-conditioned media or control media ± LPS exposure. Cell viability was determined at 24, 48, 72, 96, and 120 h. Open squares, control media only; closed squares, hAEC-conditioned media; open circles, LPS only; closed circles, LPS + hAEC-conditioned media. Treatment groups with and without LPS exposure remained significantly different from 72 h onward (p < 0.001). hAEC-conditioned media did not affect proliferation and survival of macrophages. Data are represented as mean ± SEM.
Proinflammatory M1 and Proreparative M2 Gene Expression Relative to Housekeeping Gene, 18S of Macrophages Primed With/Without Lipolysaccharide (LPS) and Cultured in hAEC-Conditioned Media or Complete Dulbecco's Modified Eagle's Medium (DMEM)/F12
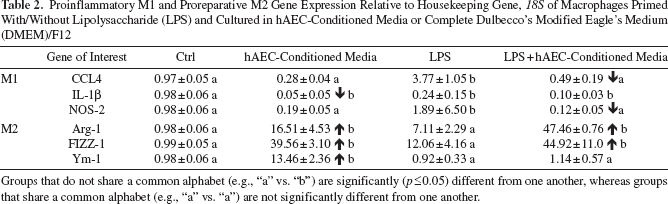
Groups that do not share a common alphabet (e.g., “a” vs. “b”) are significantly (p ≤ 0.05) different from one another, whereas groups that share a common alphabet (e.g., “a” vs. “a”) are not significantly different from one another.
To explore whether the hAEC-associated reduction in lung macrophage number that we observed in vivo was a result of recruitment, proliferation, and/or survival, we performed proliferation and migration assays. hAEC-conditioned media did not affect macrophage proliferation, regardless of whether the macrophages were exposed to LPS or not (Fig. 3C) but significantly reduced chemotaxis of macrophages toward rMIP-2 and increased the phagocytic ability of macrophages (Fig. 4A, B).

hAEC-conditioned media altered macrophage behavior. Chemotaxis toward recombinant mouse macrophage inflammatory protein-2 (rMIP-2) was significantly reduced (A), and phagocytic ability (pHRodo+ cells) was significantly increased (B) when macrophages were cultured in hAEC-conditioned media following LPS exposure. *p < 0.05, **p < 0.01. There was no significant difference in chemotaxis in macrophages cultured in hAEC-conditioned media alone (A) with no effect on the phagocytic ability compared to the control (B).
Discussion
In this study, we show that hAEC administration to bleomycin-challenged mice reduced macrophage infiltration in the lungs and promoted polarization of alveolar macrophages from a predominantly proinflammatory M1 phenotype to a proreparative M2 phenotype. Through a series of in vitro studies, we confirmed that these effects were at least in part due to soluble factors released by hAECs, rather than requiring cell-to-cell contact, and that the polarization of macrophages from M1 to M2 was associated with reduced macrophage migration and increased phagocytic activity.
Activation and accumulation of macrophages is crucial to the development of bleomycin-induced lung injury (16,28). Depletion of macrophages using liposomal chlodronate results in the attenuation of bleomycin-induced inflammation and collagen deposition (25). Here we showed that hAEC treatment following bleomycininduced lung injury reduced macrophage infiltration. This is in line with previous reports by us and others, where administration of cells from the fetal membranes reduced bleomycin-induced lung inflammation and fibrosis, coincident with reduction in macrophage infiltrate (3,22,23). In the current study, we extended that observation by showing that exposure of macrophages to hAEC-conditioned media reduced chemotactic ability of macrophages without altering their proliferation rate but increased phagocytic ability. Together, these in vitro data suggest that the reduced macrophage numbers in hAEC-mediated lung repair are a result of attenuated macrophage migration, rather than reduced proliferation or survival, consequent to soluble factors released by hAECs. hAEC-mediated increase in macrophage phagocytic ability may also have contributed to lung repair by clearing apoptotic neutrophils and hyaluronan fragments, which have been shown to perpetuate lung inflammation (27,29,32).
Polarization of macrophages from a classically activated (M1) to an alternatively activated (M2) state involves complex pathways of resistance or promotion of microbial pathogens and tumors, immune modulation, and wound healing. We acknowledge the limitations of using single surface markers to determine the phenotypes of macrophages. To overcome this, we measured gene expression of M1- and M2-specific markers. Moreover, the use of LPS alone to activate macrophages to an M1 state could be a limiting factor to this study. Despite these limitations, hAEC-conditioned media significantly altered the phenotype of macrophages toward M2.
Amnion cell-induced changes to macrophage polarity are also likely to be important in effecting tissue repair. Switching polarity from M1 to M2 contributes to wound healing (35), and mesenchymal stem cells have been previously reported to induce such a change to the macrophage population (12,18,26). Classically activated (M1) macrophages are usually activated by interferon-γ (IFN-γ) alone or in combination with microbial (e.g., LPS) or cytokine stimuli [e.g., tumor necrosis factor-α (TNF-α)]. In contrast, M2 macrophages are activated by interleukins (ILs) such as IL-4 or IL-13, which are produced in appreciable amounts by the human placenta and fetal membranes (2,5,34). In our current study, we showed that hAEC treatment reduced total macrophage numbers within the lung, of which the majority were predominantly M2 polarized. They expressed cell surface marker CD206, as well as high levels of M2-associated genes arginase-1 and Ym-1 and concurrently low levels of the M1-associated genes NOS-2, IL-1β, and FIZZ-1. This was in comparison to macrophages isolated from lungs of the bleomycin control animals, which were predominantly M1 polarized. These findings are in line with data obtained from a recent study performed by Zhang and colleagues, who reported on the ability of transplanted human gingival-derived mesenchymal stem cells to induce polarization of macrophages to the M2 phenotype and promote cutaneous wound healing in Wistar rats (35). Our in vivo findings were further supported by data from our in vitro experiments. We showed that soluble factors released by hAECs mediated the polarization of macrophages observed in vitro, and the shift of the macrophage population from M1 to M2 was due to a change in polarity of recruited macrophages rather than a change in the proliferation or survival of one phenotype over the other.
In summary, the studies reported here support the notion that hAECs do not require engraftment to repair lung injury and restore function following bleomycin injury (23,33). Instead, we have provided the first line of evidence to show that hAECs can directly modulate macrophage polarization and that this is important in altering macrophage infiltration during inflammation. Future studies should focus on the exact mechanisms by which hAECs mediate macrophage polarization as well as the potential for hAECs to influence other key immune cell types and reparative pathways.
Footnotes
Acknowledgments
We would like to thank our research midwife, Ms. Joanne Mockler, for acquiring consent from women undergoing cesarean section and, thereafter, collection of human placentae used for the isolation of hAECs used in this study. We would also like to thank Dr. Aaron Irving from Monash Micro Imaging for his donation of the pHrodo + S aureus. R.L. is supported by the Luke Proposch Perinatal Award from the Royal Australian and New Zealand College of Obstetricians and Gynaecologists. This work was partly supported by the Victorian Government's Operational Infrastructure Support Program. The authors declare no conflicts of interest.