
Abstract
Epidermal organization and homeostasis are regulated by mesenchymal influences through paracrine actions. Until today, dermal fibroblasts (DFs) are used in the “dermal” layer to support keratinocyte growth in vitro in dermal and skin substitutes. In the present work, we used human adipose tissue-derived mesenchymal cells (ADMCs) as a support of keratinocyte growth in vitro (in monolayer culture and in 3D skin cell culture models) and in vivo (mouse wound healing models) and compared our findings with those obtained using dermal fibroblasts. ADMCs induce reepithelialization during wound healing more efficiently than DFs, by enhancing keratinocyte proliferation through cell cycle progression, and migration. This effect is mediated (at least partially) by a paracrine action of KGF-1 and PDGF-BB, which are more prominently expressed in ADMCs than in DFs. Furthermore, replacement of DFs by ADMCs in the dermal compartment of organotypic skin cultures leads to an artificial epidermis resembling to that of normal skin, concerning the general histology, although with a higher expression of cytokeratins 5 and 19. In Rag1 knockout mice, ADMCs induced a more rapid reepithelialization and a more effective wound healing, compared to dermal fibroblasts. In conclusion, we provide evidence that ADMCs can serve as supportive cells for primary keratinocyte cultures. In addition, because of their abundance and the great cell yield achieved during ADMC isolation, they represent an interesting cell source, with potential aspects for clinical use.
Keywords
Introduction
The development of dermal and skin substitutes for the treatment of large or nonhealing wounds or burn coverage has been accelerated during the last two decades. Presently, full thickness skin substitutes consist of fibroblasts embedded in a biodegradable matrix and three to eight layers of primary keratinocytes. Introduction of autologous fibroblasts into grafted matrices improved wound healing (32,35). Indeed, the formation and maintenance of epidermal organization and homeostasis, as well as processes involved in wound healing, are regulated by paracrine mesenchymal–epithelial interactions. These effects are difficult to study in vivo, and therefore, several organ culture and coculture models have been developed (13,32).
Fibroblast isolation, for use in skin substitutes, requires retrieval of large full thickness skin biopsies, while cell yield is low, requiring ex vivo cell expansion. However, until today dermal fibroblasts are the main source of mesenchymal cells used for skin wound healing or generation of skin reconstructs, such as Dermagraft®, Apligraf®, and others. Bone marrow-derived mesenchymal cells (BMMCs), an evident source of mesenchymal cells, were recently introduced in wound healing trials as an alternative source of cells in dermal structures (5,17). However, bone marrow retrieval is a rather painful procedure, while cell yield is limited due to small volumes of acquired material, rendering also in vitro cell expansion obligatory. Besides, BMMC abundance decreases with age (28,30). Therefore efforts have been made to find alternative sources of mesenchymal cells. Adipose tissue has been recognized as such: adipose-derived mesenchymal cells (ADMCs) exhibit self-renewal capacity and multilineage differentiation to mesodermal cell types (9). The ability to isolate and culture stem cells, deriving from its stromal–vascular fraction, a highly abundant and easily accessible source [100 g of adipose tissue yields ADMCs equivalent to cells generated from a 6-cm2 skin biopsy, after two passages in vitro (24)], has triggered research and clinical efforts to maximize their applicability in regenerative medicine (3,10). ADMCs have been recently proposed for local delivery into dermal matrices in order to accelerate wound healing (28,29). However, few data exist and no systematic comparison has been made for their potential advantage over autologous fibroblasts in dermal substitutes.
The aim of the present work is to examine whether ADMCs can efficiently substitute dermal fibroblasts (DFs) in artificial skin reconstructs and in the support of reepithelialization during wound healing. For the first time, ADMCs are carefully compared to DFs, in respect to their ability to support keratinocyte cell growth and reepithelialization during wound healing. We show that ADMCs induce human primary keratinocyte cell proliferation and migration without requiring cell–cell contact; this effect is mediated primarily through keratinocyte growth factor-1 (KGF-1) or fibroblast growth factor-7 (FGF-7) and platelet-derived growth factor-BB (PDGF-BB). Because we used adult primary keratinocytes, which might present experimental variations due to different cell passage and differentiation state, age or sex of the donor, deriving body part, etc., we executed the experiments in parallel to a well-established keratinocyte cell line, the HaCaT cells, in which we observed similar results. Furthermore, 3D keratinocyte organotypic cultures with ADMCs embedded in the artificial dermis were developed and compared to normal skin, concerning differentiation and proliferation markers. Finally, the effect of ADMCs on reepithelialization occurring during wound healing was compared to that of DFs, in vivo in recombination activation gene knockout (Rag1-/-) mice. Our in vivo data provide evidence that ADMCs enhance reepithelialization more efficiently than DFs due to enhancement of keratinocytes proliferation and migration.
Materials and Methods
Cell Culture
Lipoaspirates were obtained by liposuction of subcutaneous adipose tissue of seven healthy donors (42 ± 12 years) under local anesthesia after informed consent of each individual. Human ADMCs were isolated as previously described (1). Isolated ADMCs were cultured in low glucose (1 g/L) Dulbecco's modified Eagle's medium (DMEM), with 10% fetal bovine serum (FBS) and penicillin/streptomycin (all from GIBCO Brl, Invitrogen, Antisel, Harilaou, Greece) and used at passages 0–5.
Total populations of human keratinocytes and fibroblasts were isolated from full thickness skin, obtained during lipectomies for cosmetic reasons after informed consent of five individuals, and cultured as previously described (32,38). In brief, primary keratinocytes and fibroblasts were isolated using the Keratinocyte Primary Isolation Kit (Cascade Biologics, Invitrogen). Keratinocytes were cultured in Cnt-57 medium (CellNTec, Switzerland) with gentamycin/amphotericin (Cascade) and were used at passages 0–4. Dermal fibroblasts (passages 0–5) were cultured in low glucose (1 g/L) DMEM, supplemented with 10% FBS and penicillin/streptomycin. HaCaT cells (ATCC, Invitrogen) were cultured in high glucose (4.5 g/L) DMEM, supplemented with 10% FBS. All cells were cultured at 37°C and 5% CO2 in a humidified atmosphere.
Of note, in the experiments executed in this study, different cell types did not match the same donors, as lipoaspirates and skin biopsies were in all cases obtained from different donors.
This study was approved by the Evaggelismos Hospital Research and Ethics Committee.
Organotypic Skin Cultures
Organotypic skin cultures were prepared as previously described (38). Eight volumes of ice-cold rat tail collagen I solution (Sigma) were mixed with one volume of 10x Hank's buffered saline and neutralized with NaOH. ADMCs or DFs were diluted in FBS and embedded in the collagen gel at a final concentration of 105 cells/ml. The mix was transferred into filter inserts (8-μm pore diameter) and equilibrated overnight in DMEM/F12 (3:1), supplemented with 10% FBS, 0.5 μg/ml insulin, 0.4 μg/ml hydrocortisone, and 0.1 μM isoproterenol. The next day, keratinocytes or HaCaT cells were plated onto the collagen gel at a density of 106 cells/cm2. After submerse incubation overnight, the cultures were raised to the air–medium interface by lowering the medium level, and ascorbic acid (50 μg/ml) plus epidermal growth factor (EGF; 0.1 ng/ml, Sigma) were added. Organotypic cultures were maintained under these conditions for 1 or 2 weeks for primary keratinocytes or HaCaT cells, respectively.
Cell Growth Assay
ADMC- or DF-conditioned medium was generated by culturing cells in basal culture medium (low glucose DMEM/10% FBS/antibiotics) for 48 h at high confluence (80–90%). Medium was cleared by centrifugation and applied to keratinocytes after mixing with normal/ untreated keratinocyte medium 1/1 (v/v) for 1–7 days. Anti-KGF-1 and anti-PDGF-BB (both from Abcam) neutralizing antibodies were used at concentrations of 10 and 1 μg/ml, respectively. Cell growth and viability were measured by tetrazolium bromide (MTT) assay (Sigma) or crystal violet staining.
Cell Cycle Analysis
Cell cycle analysis was performed by propidium iodide staining (BD Biosciences, San Diego, CA) after fixation of cells with 70% ethanol and incubation with RNase A for 30 min at 37°C. Flow cytometry (10,000 cells sampled per condition) was performed with a FACScan operator (Beckton Dickinson, Heidelberg, Germany). Results were analyzed with the FACScan (Beckton Dickinson) and the Cyflogic (Turku, Finland) software.
Cell Migration (Scratch) Assay
Primary keratinocytes or HaCaT cells were cultured until confluence and incubated for 2 h with mitomycin-C (10 μg/ml), and then a scratch was made to the culture with a tip. Cultures were treated with conditioned media, as described above. Anti-KGF-1 or anti-PDGF-BB neutralizing antibodies were used at concentrations 10 and 1 μg/ml, respectively. Photos were taken with a digital camera and analyzed by the Image J (NIH, Bethesda, MD) software.
Immunohistochemistry
Organotypic cultures or normal skin biopsies were fixed in 2% paraformaldehyde, embedded in paraffin and 3-μm-thick slices were cut. After deparaffinization and hydration, slides were incubated for 30 min with primary antibodies for cytokeratin 1 (CK1; SantaCruz Biotechnologies), CK5, CK10, CK14, CK19 (ThermoScientific), and Ki67 (Neomarkers). The UltraVision LP Detection System (TL-060-AL, Lab Vision, Freemont, US) with Fast Red, was used for immunodetection. Counterstaining was performed by Mayer's hematoxylin.
Mouse skin biopsies including the healing wound were fixed in 2% paraformaldehyde embedded in paraffin and 3-μm-thick slices were cut. After deparaffinization and hydration, slides were incubated for 30 min with 3% bovine serum albumin (BSA) and then for 1 h with primary antibody against Ki67 (1/200, Thermo Fisher Scientific, CA) in PBS, containing 1% BSA, 0.1% gelatin, and 0.05% NaN3. After a washing step, slides were incubated for 30 min with an endogenous peroxidase blocking solution (PBS containing 0.3% H2O2), washed, incubated for 30 min with a secondary anti-rabbit horse radish peroxidase (HRP) antibody (1/1,000), incubated with DAB peroxidase substrate (DAKO) for 5 min, and finally counterstained with Mayer's hematoxylin.
Immunofluorescence
Cells were detached by scrapping and washed in PBS/ 1% BSA. For direct immunofluorescence, cells were incubated for 1 h at 4°C with the following antibodies: anti-cluster of differentiation 13 tagged with phycoerythrin (CD13PE), anti-CD14 fluorescein isothiocyanate (FITC), anti-CD34PE, anti-CD45FITC, anti-CD54PE, anti-CD73PE, anti-CD90FITC (Beckton Dickinson–Pharmingen), anti-CD44FITC (J173; Immunotech/Coulter, Marseille, France), anti-CD133PE (MiltenyiBiotec, Germany), anti-CD105PE, and anti-CD144FITC (Santa Cruz Biotechnologies). Staining was quantified by flow cytometry.
For indirect immunofluorescence, cells were permeabilized or not with 0.05% TritonX-100 for 1 min on ice. Triton-treated cells were washed with PBS/10% FBS/1% NaN3. Cells were incubated for 1 h at 4°C with primary antibodies against CD31 (Thermo) and then with goat anti-mouse FITC (Santa Cruz Biotechnologies) for 1 h at 4°C, and staining was quantified by flow cytometry.
Western Blot Assay
Cells were lysed in 1% SDS, 10 mM Tris buffer, pH 7.4; 60 μg proteins were run in 8% or 10% acrylamide gels and transferred onto nitrocellulose membrane. Rabbit polyclonal anti-KGF, anti-PDGF (Abcam), anti-Ki67 (Neomarkers), or anti-actin (SantaCruz Biotechnologies) were used at concentrations 1:400, 1:400, 1:200, and 1:1,000, respectively, diluted in 5% nonfat milk (anti-KGF, anti-actin) or 5% BSA (anti-PDGF). Appropriate secondary antibodies were used in each case.
PCR
Total RNA was isolated with the Miniprep RNA isolation kit (Macherey-Nagel EURL, France), followed by DNase treatment (Invitrogen). One microgram of RNA was subjected to Thermoscript RT-PCR (Invitrogen) using poly(dT) as primers. Semiquantitative PCR was performed with Taq polymerase kit (Invitrogen) with following conditions: 94°C 3 min, 38 cycles of 94°C 30 s 55°C 30 s, 74°C 1 min, with a final step of 74°C 5 min. Real-time PCR was performed with KAPA SYBR Green Fast Universal qPCR Kit (KAPA Biosystems) in a Mx3000P apparatus (Stratagene) at following conditions: 40 cycles of 95°C for 30 s. Each set of primers was tested with at least three different RNA samples treated independently. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) and 18S expression were used as internal controls. Primers used were KGF-1: 5′-AGAAAACTAGAAAGAACTGCCC-3′ (Fw), 5′-GAAGCCTGACCTCAAAACC-3′ (Rv); PDGF-BB: 5′-GTGGCTGGAGTGGCCAAATAG-3′ (Fw), 5′-ATG GAGGTCATGTGGACAGCA-3′ (Rv); GAPDH: 5′-CGG ATTTGGTCGTATTGGGC-3′ (Fw), 5′-GCTCCTGGAA GATGGTGATG-3′ (Rv); ribosomal 18S: 5′-GTAACCC GTTGAACCCCATT-3′ (Fw), 5′-CCATCCAATCGGTAG TAGCG-3′ (Rv).
Animals
All procedures described below were approved by the Animal Care Committee of the University of Crete, School of Medicine. Rag1-/- mice (5 weeks old; female; body weight, 20–25 g; Harlan, Italy) were a gift from Dr. K. Mamalaki (FORTH). Immunodeficient animals were housed under aseptic conditions and kept for 1 week of experiment with a HEPA cage-covered filter. Animals were housed in cages, maintained under a constant 12 h light–dark cycle at 21–23°C, with free access to food and tap water. The national regulations on the use of vertebrate animals for experimental use are primarily based on the ratification of the EU regulation 86/609/ CEE; authorization number of the animal facility: 3044 (School of Medicine).
Wound Healing Model and Fibroblasts or ADMC Transplantation
For the wound healing experiments, 3-mm full thickness excisional skin wounds were created on each side of the midline with a punch. In each wound bed, a rat-tail collagen type I matrix with 500,000 human DFs, 500,000 human ADMCs, or without cells was applied. One group of mice received collagen without cells in one wound and DFs in the other, one group received collagen without cells and ADMCs, and one group received DFs and ADMCs. Animals were all housed individually. Adhesion of the matrix was inspected every hour for the first 6 h postwounding.
Digital photographs of wounds were taken at 0, 1, 2, 3, 5, and 7 days. Wound area was measured by tracing the wound margin, and surface was calculated using the Image J program. Mice were sacrificed at day 7. Skin samples including (i) the wound and 10 mm of the surrounding skin and (ii) normal adjacent skin. Samples were fixed in 2% paraformaldehyde and embedded in paraffin for histochemical examination.
Statistical Analysis
Statistical analysis of data was made by the use of the appropriate parametric test by the use of SPSS V18 microcomputer-based program (SPSS, Chicago, IL). For two-sample comparisons, we used the t test to calculate 95% confidence intervals (CIs) on differences between means and the Mann–Whitney test to obtain the p value. For multiple comparisons, we used ANOVA with the residual variance as a common estimator of error, with a Bonferroni correction. All statistical tests were two sided. All results were expressed as means ± SEM, and a value of p < 0.05 was considered to be statistically significant.
Results
Immunophenotypic Characteristics of ADMC
ADMCs, screened at passage 1, are positive for mesenchymal cell precursor markers, such as alanine amino peptidase (CD13), hyaluronan acid receptor (CD44), ecto-5-nucleotidase (CD73), and Thy-1 (CD90) at a cell percentage of 88%, 80%, 73%, and 99%, respectively; increasingly with cell passage they express endoglin (CD105) (Table 1). Furthermore, they are positive for intracellular adhesion molecule-1 (CD54; ICAM-1; 81% of whole cell population), as well as CD34 (36% of whole cell population), the latter being an important adhesion molecule that is highly expressed in hematopoietic and vascular-associated tissue and related to angiogenesis (1,40). On the contrary, ADMCs are negative (<1%) for CD31 [platelet endothelial cell adhesion molecule (PECAM)], CD14 [lipopolysaccharide (LPS) interacting protein], CD45 (protein tyrosine phosphatase, receptor type, C-associated protein), CD133 (prominin 1), CD144 [vascular endothelial (VE)-cadherin], and other markers of hematopoietic lineages. As reported previously, ADMCs possess a differentiation capacity towards adipocytes and osteocytes (1).
Surface Markers Expressed in Adipose Tissue-Derived Mesenchymal Cells (ADMCs) and Dermal Fibroblasts (DFs) Indicated Percentage = Positive Cells/Total Cell Population x 100%

Comparing the presence of the aforementioned cell surface markers in ADMCs and DFs, we observed that a smaller cell percentage of DFs are positive for mesenchymal stem cell markers (CD73, CD90, CD105). Furthermore, a greater cell percentage of ADMCs is positive for CD34 and CD54 compared to DFs, supporting a greater differentiation capacity of subpopulations of ADMCs towards endothelial cells, as it has been previously reported (1,23). A remarkable similarity of DFs and ADMCs concerning the expression of cell surface antigens and chemokines has also been described by others (18).
Humoral Effects of ADMCs on Keratinocyte Proliferation
Previous data suggest that, in order to achieve a normal keratinocyte proliferation, migration, and differentiation, a paracrine humoral effect initiated from dermal fibroblasts is necessary (37). We examined whether ADMCs, an alternative mesenchymal cell source, could support keratinocyte growth: addition of ADMC conditioned medium (1/1, v/v) to primary keratinocyte cultures increased their proliferation, attaining 48% ± 5.8% over control cells (Fig. 1A) after 48 h of treatment, an effect that was significantly higher than the one observed by DF-conditioned medium (38% ± 4.3%). This effect was sustained in a colony formation assay after 7 days of treatment (Fig. 1B). Primary keratinocytes are very sensitive to low seeding concentration; however, this was surpassed by addition of ADMC-conditioned medium, which strongly enhanced colony formation capacity and cell growth, contrarily to fibroblast-conditioned medium that had no effect. These data strongly suggest that humoral factors, contained in the ADMC medium, exert a highly supportive effect on keratinocyte cell growth. Similar effects were observed when HaCaT cells were utilized instead of primary keratinocytes (Fig. 1C). However, in contrast to highly promoting effects of ADMC over fibroblast medium in primary cells, an equivalent effect of both media was observed in HaCaT cells (compare Fig. 1A with C).
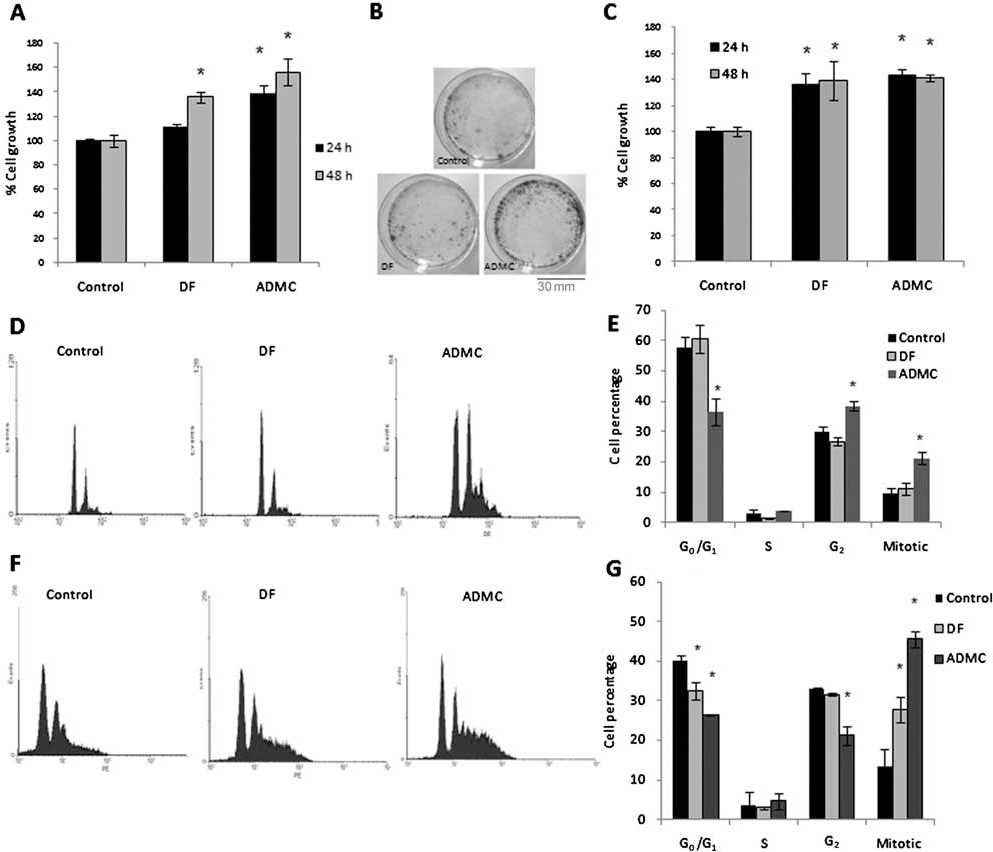
Greater induction of keratinocyte proliferation by adipose tissue derived mesenchymal cells (ADMCs) compared to dermal fibroblasts (DFs). (A) Primary keratinocytes were treated for 24 or 48 h with DF- or ADMC-conditioned medium 1/1 diluted in keratinocyte culture medium Cnt57. Cell growth (normalized over control-nontreated cells) is presented (mean ± SEM of five different assays in triplicate) and measured with MTT assay. ∗p < 0.05. (B) Primary keratinocytes, plated at a very low initial density (2,000/cm2) were treated with DF- or ADMC-conditioned medium 1/1 to Cnt57 medium for 1 week; medium was changed every second day. Cells were stained with crystal violet. The experiment was done twice. Scale bar: 30 mm. (C) HaCaT cells were treated for 24 and 48 h with DF- or ADMC-conditioned medium, and cell growth was estimated as in (A). (D–G) Primary keratinocytes (D, E) or HaCaT cells (F, G) were treated with DF- or ADMC-conditioned medium applied 1/1 to Cnt57 medium for 48 h, and cell cycle analysis using flow cytometry was performed. (E) and (G) are the quantification histograms of (D) and (F), respectively. Experiments of cell cycle analysis were carried out three times for each cell type; representative results are shown in (D) and (F).
This proliferative effect of ADMC-conditioned medium induced marked changes in cell cycle progression of keratinocytes (Fig. 1D, E) and HaCaT cells (Fig. 1F, G). In both cell types, ADMC medium significantly increased cell passage to G2 phase and mitosis, with a concomitant decrease of cells in G0/G1, a change not observed in this extent with the DF-conditioned medium.
Effect of ADMCs on Keratinocyte Migration
Reepithelialization during wound healing relies on two procedures: keratinocyte proliferation and migration. ADMC-conditioned medium induced a significant enhancement of keratinocyte and HaCaT cell migration (Fig. 2A and B, respectively). In contrast, DF-conditioned medium had a significantly lower effect (Fig. 2A, B). Additionally, ADMCs induced matrix metalloproteinase 9 (MMP9) and MMP2 collagenase but not gelatinase activities, supporting the enhanced migratory activity of ADMC medium; however, no changes have been observed in MMP1 activity in the presence of conditioned media (data not shown).

Greater induction of keratinocyte migration by ADMC compared to DF. (A, B) Primary keratinocytes (A) or HaCaT cells (B) were cultured until confluence, and they were treated for 2 h with mitomycin-C (10 μg/ml). A scratch was made with a tip (time 0), and cells were treated with DF- or ADMC-conditioned medium applied 1/1 in Cnt57 medium. Healing was measured by examining photos taken at different time points. Four experiments have been performed with similar results; representative results are shown in the photos. A quantification of all experiments after 24 h is shown in the right graphs. ∗p < 0.05. Scale bar: 150 μm.
ADMCs Express KGF-1 and PDGF-BB, Which Mediate Their Effect on Keratinocyte Proliferation and Migration
The above data suggest that ADMCs might secrete a number of humoral factors, enhancing keratinocyte proliferation and migration. Here, we concentrated on two factors previously described to induce keratinocyte proliferation and migration, KGF-1 (FGF-7) and PDGF-BB. Keratinocyte growth factor-1 (KGF-1) is expressed in wounds and stimulates proliferation and migration of keratinocytes (33,44). Platelet-derived growth factor-BB (PDGF-BB) affects reepithelialization and tissue remodeling (6,20). ADMCs synthesize significantly higher amounts of KGF-1 mRNA (Fig. 3A) and protein (Fig. 3B), as compared to DFs (by 14-fold at the mRNA and 4.3-fold at the protein level) and PDGF-BB (4.3- and 2-fold increase at the mRNA and protein level, respectively) (Fig. 3A, B).

ADMCs exhibit higher keratinocyte growth factor 1 (KGF-1) and platelet-derived growth factor-BB (PDGF-BB) expression than DFs. Total RNA and protein from DF and ADMC cultures were isolated as reported in Material and Methods. (A) Detection of KGF-1 and PDGF-BB expression by real-time PCR (histogram) and semiquantitative PCR (photo). 18S or GAPDH signal was used as the internal control for real-time or semiquantitative PCR, respectively. The graph represents the results of three independent experiments in triplicates. (B) Detection of KGF-1 and PDGF-BB by Western blot; the signal of each protein is normalized to the signal of actin. The photo shows the result of one representative experiment; the graph is the quantification of two independent experiments.
When primary keratinocytes were exposed to ADMC-conditioned medium, in the presence of neutralizing antibodies against KGF-1 or PDGF-BB, a significant reduction of growth was observed (Fig. 4A). These results are in agreement with the expression of KGF-1 and PDGF-BB in ADMCs. Dermal fibroblast-conditioned medium induced similar (although less pronounced) effects (Fig. 4A). The growth of keratinocytes in control medium was slightly inhibited by anti-KGF-1 antibody, suggesting KGF-1 secretion by keratinocytes themselves, supporting an additional autocrine cell growth mechanism. On the contrary, neutralization of PDGF-BB did not affect keratinocyte proliferation in control samples (not shown). Similar results were observed in HaCaT cells (Fig. 4B).

The effect of ADMCs on keratinocyte proliferation is inhibited by neutralizing antibodies against KGF-1 and PDGF-BB. Primary keratinocytes (A) and HaCaT cells (B) were cultured in the presence of DF- or ADMC-conditioned media diluted 1/1 in Cnt57 medium, in the absence (black bars) or presence of neutralizing antibodies against KGF-1 (aKGF-1; 10 μg/ml) and PDGF-BB (1 μg/ml) for 48 h. Neutralizing antibodies were applied to cells simultaneously with conditioned media. Cell growth was measured by MTT assay. Mean ± SEM of three different experiments assayed in triplicate. ∗p < 0.05 compared to control-nontreated cells. (C) Primary keratinocytes were cultured in the above described conditions, and total protein was collected and examined with Western blot analysis on a 8% polyacrylamide gel for Ki67, using the signal of actin as an internal control. A quantification of one representative experiment is shown here. (D) Organotypic skin cultures were developed by culturing primary keratinocytes on top of a collagenous matrix containing ADMCs for 1 week, in the absence or presence of neutralizing antibodies against KGF-1 (10 μg/ml) and PDGF-BB (1 μg/ml). Scale bar: 100 μm.
An additional proof of the implication of KGF-1 and PDGF-BB contained in ADMC-conditioned medium on keratinocyte proliferation is presented in Figure 4C: ADMC supernatant increased Ki67 expression, suggesting induction of keratinocyte proliferation, an effect partially reversed by KGF-1 and PDGF-BB neutralizing antibodies. A similar, but less pronounced, effect was obtained with DF-conditioned medium, tallying with the higher expression of these factors in ADMCs. Moreover, Ki67 expression in primary keratinocytes of organotypic skin cultures containing ADMCs in the collagen matrix was significantly reduced by the presence of neutralizing antibodies against KGF-1 and PDGF-BB (Fig. 4D), further supporting the inductive effect of ADMCs on keratinocytes proliferation though secretion of KGF-1 and PDGF-BB.
A similar implication of KGF-1 and PDGF-BB was evidenced on primary keratinocyte cell migration (Fig. 5). Addition of anti-KGF-1 (Fig. 5B) and anti-PDGF-BB (Fig. 5C) significantly inhibited the effect of the ADMC-conditioned medium on keratinocytes migration. However, the less prominent effect of DF medium was not reversed by KGF-1 or PDGF-BB depletion (Fig. 5D). Similar results were also observed with HaCaT cells (not shown). These observations suggest that the inductive effect of ADMCs on keratinocytes migration is mediated, at least in part, by KGF-1 and PDGF-BB and subsequent mechanisms triggered by these factors. However, the effect is partially reversed, suggesting the implication of additional regulatory mechanisms.

The effect of ADMCs on primary keratinocytes migration is inhibited by neutralizing antibodies against KGF-1 and PDGF-BB. Primary keratinocytes cultured until confluence were treated with DF- or ADMC-conditioned media diluted 1/1 in Cnt57 without neutralizing antibodies (A), with anti-KGF-1 (10 μg/ml) (B), or anti-PDGF-BB (1 μg/ml) (C), and photos of the scratch were taken after 24 h. (D) A quantification of three experiments is shown. Representative results are shown in (A–C). ∗p < 0.05. Scale bar: 150 μm.
ADMCs Used as Feeder Cells in the Dermal Compartment of Organotypic Skin Cultures
In order to examine whether ADMCs can replace DFs as feeder cells for keratinocytes in skin substitutes, we developed 3D organotypic skin cultures, with either DFs or ADMCs in the collagen type I matrix and primary keratinocytes or HaCaT cells [used in multiple studies for the development of organotypic culture models (7,35,39)] cultured on top. We examined both cell morphology and expression of cytokeratins [CK5 and 14, normally expressed only in the keratinocytes of the basal layer, CK1 and 10, expressed by keratinocytes in the suprabasal layers and CK19, known to be a biochemical marker for proliferating epidermal stem cells and keratinocytes (26)].
Primary keratinocytes in organotypic cultures with DFs in the collagen compartment formed five to eight epidermal layers, however presenting a limited organization, compared to normal skin (Fig. 6, left). In the presence of ADMCs in the dermal matrix, the artificial epidermis presented a similar thickness. However, in cultures with DFs, CK5 was mainly present in the lower “epidermal” layer, while cultures with ADMCs showed a rather abnormal expression of CK5, which was present in all keratinocyte layers. CK14 expression was similar in 3D cultures with DFs or ADMCs, although being greater in the lower “epidermal” layers in the presence of ADMCs. CK1 and CK10 had similar expression in 3D cultures with either DFs or ADMCs. Finally, the artificial epidermis of cultures with ADMCs exhibited higher expression of CK19 compared to the one of cultures with DFs, standing in accordance with the stronger induction of keratinocyte proliferation by ADMCs.
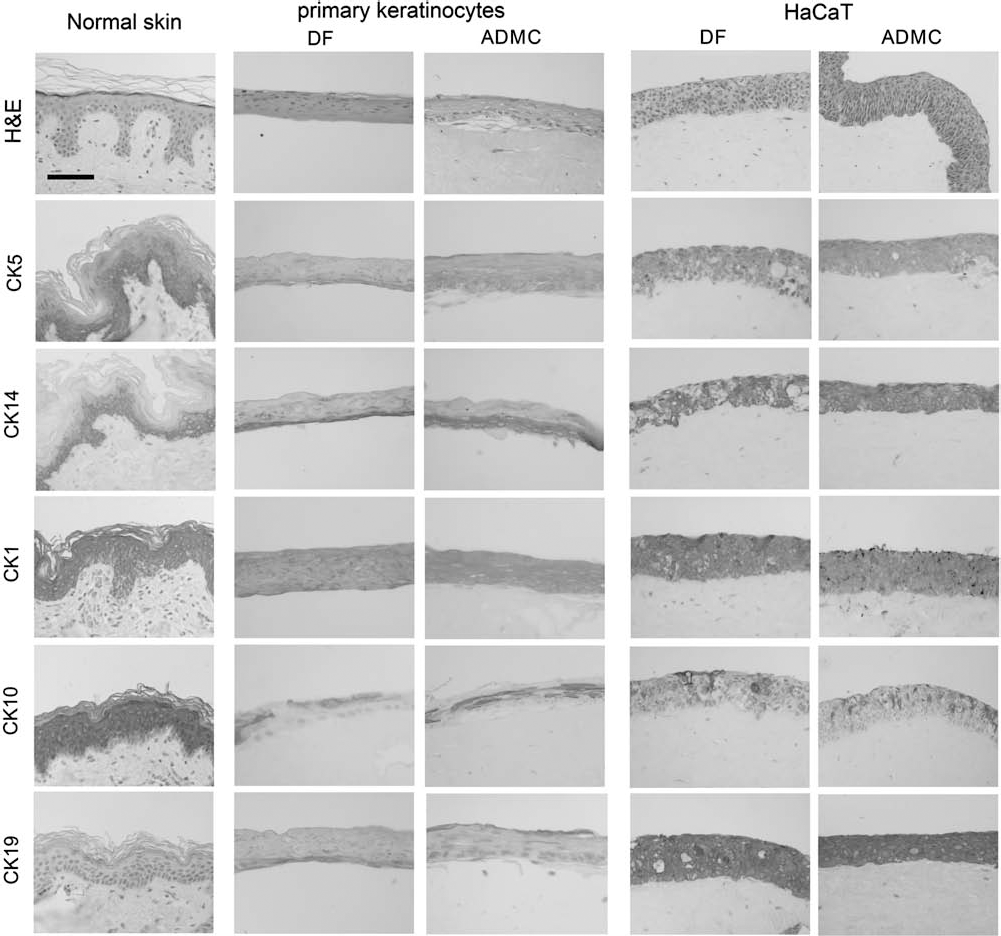
Organotypic skin cultures of primary keratinocytes (left) or HaCaT cells (right) with DFs or ADMCs. Three-dimensional organotypic skin cultures (OTC) have been developed using primary keratinocytes or HaCaT cells for the epidermal compartment and DFs or ADMCs for the dermal compartment. Cultures were fixated in 2% paraformaldehyde and embedded in paraffin, and 3-μm slides were processed for hematoxylin–eosin (H&E) or cytokeratins staining, as described in Materials and Methods. Scale bar: 100 μm.
The expression of cytokeratins in the epidermal layers of HaCaT organotypic cultures was very different from the pattern observed in normal skin: expression of CK5 and CK14 was evident in all layers and was higher in the presence of ADMCs compared to DFs. CK19, which was highly expressed throughout the “epidermis,” matching with the strong proliferative capacity of HaCaT cells, was also somewhat induced by the presence of ADMCs compared to DFs. Finally CK1 and CK10 exhibit in HaCaT 3D cultures an abnormal expression, which was similar in the presence of either DFs or ADMCs (Fig. 6, right).
ADMC Induce Reepithelialization in a Mouse Wound Healing Model
Our data show that ADMCs promote keratinocytes proliferation and migration in vitro. In order to verify whether a similar result could also occur in vivo, we used immunodeficient Rag1-/- mice, in which cutaneous wounds were created and human ADMCs or DFs embedded in a collagen type I matrix, were applied (Fig. 7A). A cellular collagen matrix was used as a negative control. Both cell types (ADMCs and DFs) significantly promoted reepithelialization and induced more efficient wound closure compared to negative control wounds (Fig. 7A, B). However, in the presence of ADMCs the formed epithelium was much thicker than in control wounds (Fig. 7B). Wounds that received ADMCs had developed at 3 days a thin epidermal layer (not seen in DF-grafted animals) and presented at 7 days a thicker epidermis (Fig. 7C), with keratinocytes exhibiting higher expression of Ki67 (Fig. 8). Furthermore, the dermis in the presence of DFs was richer in connective tissue (Fig. 7C), and inflammatory elements were less prominent in ADMC-treated compared to DF-treated wounds at 7 days (not shown), suggesting an earlier and more efficient healing procedure in the presence of ADMCs.

ADMCs induce more efficiently than DF reepithelialization and wound healing in vivo in immunodeficient mice. (A–C) Three-millimeter full thickness excisional skin wounds were created on each side of the midline of Rag1-/- mice with a punch, and DFs or ADMCs, cultured for 24 h in a collagen type I matrix or an acellular matrix (negative control), were applied in each wound. (A) Three mice received an acellular collagen gel on the left and a collagen gel with DFs on the right wound. (B) Three mice received an acellular collagen gel on the left and a collagen gel with ADMCs on the right wound. (C) Three mice received a collagen gel with DFs on the left and a collagen gel with ADMCs on the right wound. Photos were taken at days 0, 1, 2, 3, 5, and 7. Representative photos and biopsies of one mouse of each group are shown here. Scale bar: 300 μm. Graphs represent wound closure (wound bed size vs. time) in all mice of each group. CR, crust; IF, inflammatory features; NE, new epithelium.

Higher Ki67 expression in wounds with ADMCs. Representative photos of Ki67 staining of two wounds of the same mouse, having received DFs or ADMCs. Scale bar: 150 μm. Some Ki67-positive cells are indicated by arrows.
Discussion
Skin engineering refers to the development of dermal or skin substitutes for temporary or permanent replacement in large acute or chronic wounds, ulcers, or burns. Production of epidermal sheets is still under constant improvement, with a variety of culture techniques and dermal matrices being developed (25). Irradiated or mitomycin C-treated fibroblasts boost primary keratinocyte proliferation and have been used in cocultures with keratinocytes (32,45). Furthermore, introduction of autologous dermal fibroblasts into the grafted matrices of artificial skin accelerated wound healing (35,37). Our data confirm these findings, as human adult dermal fibroblasts induced keratinocyte proliferation and migration in vitro and accelerated skin wound healing and reepithelialization in vivo. Additionally, we examined whether ADMCs could represent an alternative population of cells to facilitate keratinocyte proliferation and wound healing.
Initially, BMMCs have been studied in wound healing. Treatment with BMMCs accelerated wound healing in diabetic rats (19), while they have been used in clinical studies for the treatment of chronic wounds, with encouraging results (5,17). Recently, full thickness engineered skin has been developed by the differentiation of BMMCs to epidermal cells and fibroblasts (15). An attractive alternative source of mesenchymal cells are ADMCs due to the abundance of adipose tissue and its great yield of mesenchymal cells, permitting their use without any ex vivo culture steps (24,34). These cells are known to share similarities with BMMCs (1) and fibroblasts, as reported here and by others (18,46). ADMCs consist of several cell subpopulations, such as preadipocytes, capillary endothelial cells, macrophages, and mesenchymal stem cells (36,41), the latter being characterized by positivity for CD44/CD73/CD90/ CD105 (1,18). ADMCs exhibit higher expression of these markers, compared to dermal fibroblasts, indicative of their greater plasticity. In addition, ADMCs exhibit important advantages compared to fibroblasts, as they release angiogenic factors and chemokines (16,21,31), while they may differentiate to endothelial cells (12,23,42), thus enhancing neovascularization (27,29,47), a crucial element of successful wound healing. The use of ADMCs in skin substitutes has been firstly described by El-Ghalbzouri (11); our results stand in accordance with those of El-Ghalbzouri, showing the development of a differentiated epidermis in the presence of ADMCs in the dermis. However, these authors also reported a strong contraction of the dermal matrix, which we did not observe, as well as low lateral keratinocytes migration and delayed collagen IV deposition at the epidermal/matrix junction induced by the presence of ADMCs in the dermis. Ever since, very few studies have examined ADMCs in skin reconstruction (2,3). Recently, Trottier and his coworkers suggested that ADMCs could usefully substitute dermal fibroblasts for skin reconstruction using a self-assembly method (43). Our data support these findings and provide a possible mechanism. Indeed, we show that ADMCs induce proliferation and migration of human adult primary keratinocytes, with a net advantage over dermal fibroblasts, promoting cell cycle progression and a shift of keratinocytes from G0/G1 to G2/M phase. We also observed a similar induction of cell growth in immortalized HaCaT keratinocytes, which do not exhibit variations related to donor or sex age, or passage number as usually do the adult primary keratinocytes, a fact that further supports our hypothesis.
Previous results suggest that the induction of keratinocyte growth by fibroblasts is mediated by diffusible fibroblast-produced factors (37). Furthermore, this effect is induced by cocultured keratinocytes, which actively induce the secretion of growth factors, such as KGF (22). Previous studies, suggested that direct cell–cell contact is required for mesenchymal cells to promote keratinocyte growth (45). In contrast, here we show that ADMC- or fibroblast-conditioned media exert proliferative and migratory stimuli on keratinocytes, through a paracrine release of KGF-1 and PDGF-BB, not necessitating cell-to-cell contact.
KGF-1 and PDGF-BB are implicated in paracrine effects of fibroblasts on keratinocytes (22). KGF is a major regulator of keratinocyte growth, differentiation, and morphogenesis (33). In vivo, KGF is strongly up-regulated in dermal cells of wounded skin (8,44), and targeted overexpression of KGF in keratinocytes leads to epidermal hyperplasia (14). PDGF is also upregulated during wound healing, promoting keratinocyte proliferation (6). As shown here, ADMCs exhibit a high expression of KGF-1 and PDGF-BB, much higher than fibroblasts, a fact that explains, at least in part, their stronger effects on keratinocyte proliferation and migration. Neutralization of KGF-1 and PDGF-BB partially inhibits these inductive effects, suggesting the implication of additional growth factors, such as hepatocyte growth factor (HGF), or cytokines such as interleukin (IL)-6 or IL-8, known to promote keratinocyte proliferation and being also produced by ADMCs at high levels (13, 16). Furthermore, KGF-1 and PDGF-BB might be implicated in complex networks in order to regulate the processes of wound healing (keratinocyte proliferation, migration, reepithelialization). For instance, exogenous KGF has been shown to induce granulocyte macrophage-colony stimulating factor (GM-CSF) and other cytokines (such as IL-1α) release (4). ADMCs embedded in the collagen compartment of organotypic skin cultures modifies the morphology of the artificial epidermis, which exhibits higher and broader expression of cytokeratins 5 and 14, which are normally expressed only in basal keratinocytes.
Furthermore, strong induction of keratinocyte proliferation by human ADMCs compared to DFs has been also observed in an in vivo mouse wound healing model. Application of ADMCs in a collagenous matrix induced, compared to DFs, more efficient reepithelialization due to higher keratinocyte proliferation, evidenced by higher Ki67 expression. Furthermore, wounds having received ADMCs exhibit less inflammation, and less connective tissue formation features resembling more to those of normal skin than of scar tissue (46).
In conclusion, in the present work, we provide evidence that adipose-derived mesenchymal cells induce reepithelialization stimulating wound healing through induction of keratinocytes proliferation and migration, mediated by a paracrine action of KGF-1 and PDGF-BB. ADMCs are shown to serve as a new keratinocyte supporting dermal layer, exhibiting two major advantages, compared to dermal fibroblasts: (i) their great abundance and ease of their isolation and (ii) their greater proliferative effect on keratinocytes. Achieving promotion of reepithelialization is important in the treatment of nonhealing wounds. However, strong induction of keratinocytes proliferation and migration, as well as induction of abnormal CK5, CK14, and CK19 expression in 3D skin cultures, could be considered as hallmarks of a potential hyperproliferation-inducing behavior of ADMCs. Therefore, although being a promising cell source in the treatment of nonhealing wounds, ADMCs and their effects on keratinocytes have to be thoroughly examined in vitro and in vivo prior to considering their possible clinical use.
Footnotes
Acknowledgments
The excellent technical assistance of Mrs. Maria Klinaki is acknowledged. The authors declare no conflicts of interest.