
Abstract
Although islet transplantation may restore insulin independence to individuals with type 1 diabetes mellitus, most have abnormal glucose tolerance. We asked whether the defective glucose tolerance is due to inadequate β-cell mass or to impaired insulin sensitivity. We performed metabolic studies on four cynomolgus primates before inducing diabetes with streptozotocin (STZ), then again 2–3 weeks after restoring insulin independence via intrahepatic islet transplantation utilizing a calcineurin inhibitor-free immunosuppressive regimen (induction with rabbit antithymocyte globulin and maintenance therapy with rapamycin). Engrafted β-cell mass was assessed by acute insulin and C-peptide responses to glucose (AIRglu and ACRglu) and arginine (AIRarg and ACRarg). Insulin sensitivity (SI) was determined in naive and transplanted primates from an intravenous glucose tolerance test using the minimal model. α-Cell function was determined by the acute glucagon response to arginine (AGRarg). Glucose tolerance (K g ) decreased from 4.1 ± 0.5%/min in naive primates to 1.8 ± 0.3%/min in transplanted primates (p < 0.01). Following transplantation, AIRglu was 28.7 ± 13.1 μU/ml compared to 169.9 ± 43.1 μU/ml (p < 0.03) in the naive condition, ACRglu was 14.5 ± 6.0 ng/ ml compared to 96.5 ± 17.0 ng/ml naive (p < 0.01), AIRarg was 29.1 ± 13.1 μU/ml compared to 91.4 ± 28.2 μU/ml naive (p < 0.05), and ACRarg was 1.11 ± 0.51 ng/ml compared to 2.79 ± 0.77 ng/ml naive (p < 0.05). S I did not differ from naive to posttransplant states. AGRarg was reduced in transplanted primates (349 ± 118 pg/ml) when compared to both naive (827 ± 354 pg/ml) and post-STZ diabetic primates (1020 ± 440 pg/ml) (p < 0.01 for both comparisons). These data suggest that impaired glucose tolerance observed in islet transplant recipients is secondary to low functional β-cell mass and not to insulin resistance shortly after transplant. Furthermore, improved glycemic control achieved via islet transplantation over the diabetic state might be attained, in part, via reduced glucagon secretion.
Introduction
Islet transplantation is a promising way of restoring insulin independence to patients with type 1 diabetes mellitus (T1DM) (8). When assessed by oral glucose tolerance testing, most islet transplant recipients do not regain normal metabolic control (6,29). Therefore, we asked whether the suboptimal glycemic control observed following islet transplantation is due to inadequate islet β-cell mass or impaired insulin sensitivity. Recent evidence has shown that calcineurin inhibitors utilized in islet transplantation regimens induce β-cell toxicity in both rodent and human islets (13,30). Metabolic studies of calcineurin inhibitor-free immunosuppressive regimens for islet transplantation, however, are lacking. Our islet transplant model using nonhuman primates (NHPs) afforded a unique opportunity to address the question since we could assess both islet function and insulin sensitivity longitudinally (prior to diabetes onset, after diabetes induction, and again after receiving an islet transplant with restored euglycemia while on rapamycin monotherapy).
Materials and Methods
Animals
The procedures described in this study were conducted according to the principles set forth by the National Research Council, and were approved by the NIH Animal Care and Use Committee (IACUC).
The four islet donors were 8–10-year-old cynomolgus macaques (Macaca fascicularis, NIH nonhuman primate facility, Poolesville, MD) maintained on a regular primate diet with a continuous water supply and fresh fruits twice daily. Donors weighed 3.0–4.4 kg and were ABO-compatible and major histocompatibility complex (MHC) mismatched to recipients. The four recipients were a subgroup of previously studied Cynomolgus macaques (12) with an age range of 4.5–7 years and a weight range of 2.4–3.1 kg. Vascular access was obtained by use of surgically implanted portacaths into each primate's internal jugular veins at least 2 weeks prior to initial metabolic testing.
Diabetes was induced as described previously by selectively infusing 70 mg/kg body weight streptozotocin (STZ, Pharmacia & Upjohn) directly into the pancreatic arterial bed to result in selective β-cell destruction (32). Diabetes was maintained for at least 1 month and absolutely deficient endogenous insulin production was then confirmed by a negative C-peptide response to an arginine stimulation test. Diabetic primates were treated with 2–3 U/kg body weight of glargine insulin (Lantus©) injected each morning. Fasting and postprandial glucose levels were monitored by three daily tail stick measurements.
Pancreases were procured from donor primates and islet isolation was performed using the automated and Liberase-based method for human islet isolation (28). Islets were surgically infused into the portal vein through a branch of the superior mesenteric vein as previously described (11). To prevent graft rejection, primates were treated with rabbit antithymocyte globulin (rATG) (Thymoglobulin Sangstat, CA) and rapamycin (sirolimus) (Rapamune, Wyeth-Ayerst) (12). rATG (20 mg/kg) was administered intravenously daily for 7 days, with the first dose given immediately prior to transplantation. Rapamycin was given once a day orally, to target 24-h trough levels of 10–15 ng/ml and measured weekly as previously described (12).
Metabolic studies were completed in the naive state (2–4 weeks prior to STZ-induced diabetes), 2 weeks post-diabetes induction, and 14–20 days following islet transplantation. Only arginine stimulation testing was performed post-diabetes induction to prevent severe hyperglycemia due to the mixed meal tolerance test or intravenous glucose tolerance test (IVGTT).
Metabolic Studies
All procedures were performed using primates fasted overnight. Primates were sedated with intramuscular Telazol (Fort Dodge Animal Health, Fort Dodge, IA) at a dose of 3–6 mg/kg. The portacaths were accessed for blood sampling while an intravenous catheter was inserted into the contralateral saphenous vein for infusions.
Mixed Meal Stimulation Test
After basal blood sampling, 30 ml of Ensure® was administered via nasogastric tube. The primate was returned to its cage and retrieved at 90 min for another blood sample.
Insulin Modified Frequently Sampled Intravenous Glucose Tolerance Test
After two basal blood samples were taken, an intravenous bolus of 0.27 g/kg glucose as a 50% solution was injected at time zero (t = 0 min), and blood samples were drawn 2, 4, 8, and 19 min later. At 20 min, regular insulin (0.005 U/kg) was injected intravenously. Further blood samples were drawn at 22, 30, 40, 50, 70, 90, and 180 min.
Arginine Stimulation Test
After two basal blood samples, 2 g arginine (as a 10% solution) was infused by intravenous bolus and blood samples were then taken at 2, 3, 4, 5, 7, 10, and 15 min later.
Sample Handling
Blood samples were collected in chilled tubes and centrifuged soon afterwards. Serum samples were decanted and frozen at −70°C for later insulin, C-peptide, and glucagon assays and plasma was frozen at −70°C for glucose, glycerol, and free fatty acid assays.
Biochemical Determinations
Glucose was measured in plasma using the glucose oxidase method. Serum insulin and C-peptide were measured using a commercially available human ELISA (Alpco Diagnostics, Wyndham, NH), and glucagon was measured using a commercially available radioimmunoassay (RIA) kit (Linco diagnostics, St. Charles, MO). Plasma glycerol was assessed by an enzymatic calorimetric assay (Sigma Diagnostics, St. Louis, MO). Plasma free fatty acids were assessed enzymatically (Wako Chemicals, Richmond, VA). All assays were performed in duplicate.
Calculations
Insulin sensitivity (SI) and glucose effectiveness (SG) were calculated from glucose and insulin values during the IVGTT using the minimal model of glucose kinetics (2). The acute insulin response to glucose (AIRglu) was computed from the insulin levels during the IVGTT as the integrated incremental insulin concentrations above basal levels obtained during the first 10 min. Glucose disappearance (K g ) was calculated as the slope of the line plotting the natural logarithm of glucose values between 4 and 19 min of the IVGTT versus time. The acute C-peptide response to glucose (ACRglu) during the IVGTT was calculated as the incremental area under the curve (AUC) using the trapezoidal rule for the first 19 min of the IVGTT. The disposition index (DI) was calculated as the product of AIRglu (in pmol/L) and SI (14). Data were calculated using the MINMOD 3 software (25).
The acute insulin response to arginine (AIRarg) and acute C-peptide response to arginine (ACRarg) were quantified as mean incremental increases above baseline levels within the first 5 min following arginine injection. The acute glucagon response to arginine (AGRarg) was quantified as the incremental AUC for glucagon levels from 2 to 7 min following arginine injection.
Statistical Analysis
Comparisons of experimental determinations between groups were made utilizing ANOVA, followed by Bonferroni post hoc tests, and two-tailed Student's t-tests. Results are expressed as means ± SEM unless otherwise noted.
Results
All primates (n = 4) were rendered C-peptide negative following intra-arterial STZ. C-peptide-negative diabetes was confirmed by repeated fasting glucose levels above 250 mg/dl, and, 1 month later, by nonmeasurable C-peptide secretion in response to an arginine stimulation test. Following islet transplantation with 9,215 ± 2,420 IEQ/kg body weight, all primates were rendered insulin independent with excellent glycemic control, as previously described (12) (Table 1). The animals' average weight prior to diabetes induction was 2.7 kg, after diabetes induction the average weight was unchanged, and after islet transplant the animals' weight ranged from 2.5 kg (min) to 2.6 kg (max). The most extreme weight loss occurred in an animal that weighed 3.1 kg prior to diabetes onset, 2.85 kg after diabetes induction, and a nadir of 2.6 kg following islet transplant with recovery to 2.75 kg at autopsy. All recipients had supratherapeutic trough rapamycin levels (15–30 ng/ml) during the time period of metabolic studies (12).
Metabolic Data From Individual Transplant Recipients

Individual metabolic data obtained from each primate. SI (insulin sensitivity), AIRglu (acute insulin response to glucose), and DI (disposition index) values were obtained by intravenous glucose tolerance test (IVGTT) and analyzed by MINMOD3 software. AIRarg (acute insulin response to arginine) and AGRarg (acute glucagon response to arginine) values obtained by arginine stimulation testing. FPG, fasting plasma glucose; STZ, post-streptozotocin-induced diabetes; Tx, postislet transplantation. NA, data not available, study not performed; ND, not detected.
Fasting whole blood glucose concentrations in STZ-treated primates obtained by tail-stick glucometer measurements.
Mixed Meal Stimulation Test
Insulin and C-peptide values between the naive (n = 8, including donors) and transplanted (n = 4) primates were comparable at both 0 and 90 min. Fasting glucose values between the groups were also similar. However, at 90 min, glucose values were higher in transplanted primates compared to naive primates (138.6 ± 3.7 vs. 66.5 ± 6.8 mg/dl) (p < 0.01) (Fig. 1).

Mixed meal stimulation test-derived (A) glucose, (B) insulin, and (C) C-peptide concentrations ± SEM. Time 0 samples are listed in white with 90-min samples shown in black. *p < 0.01.
IVGTT
The mild posttransplant glucose intolerance observed during mixed meal tolerance testing was also observed during the IVGTT, with significant reductions in glucose tolerance as assessed by the glucose disposal rate (Kg) in the transplanted group (Fig. 2A, Table 2). The first phase insulin and C-peptide responses were diminished in primates following islet transplantation compared to the same primates before diabetes was induced (Fig. 2B, C). This is best demonstrated by the acute insulin response to glucose (AIRglu), which was diminished 83.3% from baseline (Table 2). Similarly, the acute C-peptide response to glucose (ACRglu) was 96.5 ± 17.0 ng/ml in naive primates but only 14.5 ± 6.0 ng/ml in the transplant group (p < 0.01). Minimal model-derived SI and SG values calculated from the IVGTT data were not different among the groups (Table 2). The impaired first-phase insulin secretion (whether measured by AIRglu or ACRglu) best explains the decreased glucose disappearance rate (Kg) seen in the transplanted group as neither insulin sensitivity (SI) nor glucose effectiveness (SG) was impaired. When comparing the native to the posttransplantation state, the calculated disposition index (DI) was significantly reduced (Table 2), further suggesting that the β-cells are not compensating adequately for the prevailing insulin sensitivity.
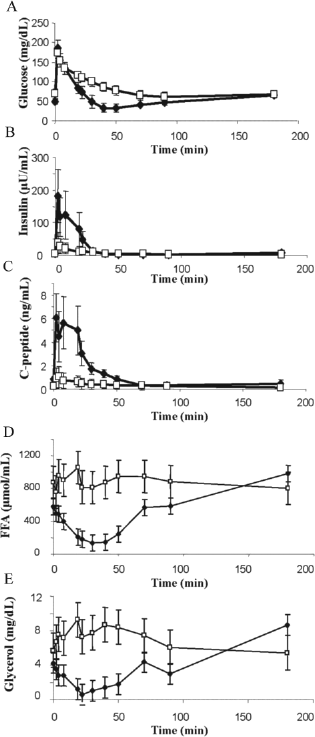
Comparison of (A) glucose, (B) insulin, (C) C-peptide, (D) free fatty acids (FFA), and (E) glycerol concentrations ± SEM during the intravenous glucose tolerance test (IVGTT). The responses of the naive primates are shown in black, while white markers indicate those of the islet transplant recipients (p < 0.001 for both FFA and glycerol by ANOVA).
Metabolic Data From Frequently Sampled Intravenous Glucose Tolerance Test

Data obtained from IVGTT, analyzed by MINMOD3 software, and presented as mean ± SEM for each measurement (n = 4/group). AIRglu, acute insulin response to glucose;, K g , glucose disappearance; SG, glucose effectiveness; SI, insulin sensitivity.
Consistent with the lower circulating insulin levels found in the animals post-islet transplant, we observed that while naive animals show the expected decline in plasma free fatty acid (FFA) and glycerol levels in a fashion parallel to insulin secreted during the IVGTT, transplanted primates show no such decrease in FFA and glycerol (Fig. 2D, E).
Arginine Stimulation Test
The acute insulin response to arginine (AIRarg) was reduced from 91.4 ± 28.2 μU/ml in naive primates (n = 8, including donors) to 29.1 ± 13.1 μU/ml in transplanted primates (n = 4, p < 0.05). Similarly, ACRarg was reduced from 2.79 ± 0.77 ng/ml in naive primates to 1.11 ± 0.51 ng/ml in the transplanted primates (p < 0.05).
Consistent with previous reports (34), we observed a robust glucagon response to arginine in the diabetic state; however, the glucagon response was markedly reduced following islet transplantation (Fig. 3). The AGRarg was 827 ± 354 pg/ml in the naive state and 1,020 ± 440 pg/ml in the post-STZ diabetic state with a significant decline to 349 ± 118 pg/ml in the posttransplantation state (p < 0.01 for both comparisons).
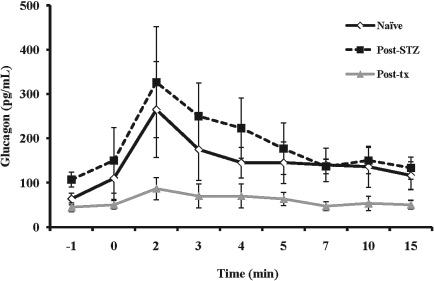
Glucagon concentrations ± SEM following arginine stimulation with suppressed α-cell response in posttransplant animals (solid gray line) compared to naive (solid black line) and diabetic (dotted black line) animals (p < 0.01 for both naive and diabetic animals vs. posttransplant animals by ANOVA).
Discussion
We demonstrate here through reductions in the acute insulin and C-peptide responses to glucose and arginine that a low islet β-cell mass relative to the naive state is the principal cause of the impaired glucose tolerance observed following intrahepatic islet transplantation. Furthermore, rapamycin monotherapy is not associated with insulin resistance in the early posttransplant period. Finally, our results demonstrate that the improved glycemic control following engraftment of a suboptimal islet mass may be achieved, in part, through suppression of glucagon secretion.
The apparent reduction in islet function we observe in transplanted primates may be due to mechanisms other than reduced functional β-cell mass. Reductions in circulating insulin concentration could result from increased insulin extraction by the liver. This is not likely, however, as human recipients of intrahepatic islet transplant have been reported to have both normal insulin-mediated suppression of hepatic glucose production and hepatic insulin extraction (20,22), as well as our observations of reduced C-peptide levels, which is unaffected by liver extraction and not disproportionately different than insulin levels in this study. Our primates did have elevated systemic rapamycin levels (12), possibly inhibiting islet function. Unfortunately, consistent rapamycin levels were difficult to maintain (11) because of reluctance of the animals to take oral rapamycin and due to its gastrointestinal toxicities (24). As orally administered rapamycin results in a 1.65-fold higher drug concentration in portal relative to peripheral vein blood, islets infused into the portal vein are exposed to slightly higher sirolimus concentrations (5). Supratherapeutic sirolimus concentrations have been shown to have a deleterious effect on rat and human islets in vitro (1), and on rat islet function in vivo (19). On the other hand, the improved insulin secretion shown in minipigs treated with sirolimus in vivo, as well as the extremely low incidence of diabetes reported in human nonislet transplant recipients treated with sirolimus in the absence of glucocorticoids, both suggest that sirolimus may not cause significant islet functional impairment (15,21).
Islet functional assays may also be limited by a relative lack of vascular supply of the transplanted islets. Revascularization of islets in primates may take between 7 and 30 days following transplant (10) and within 10–14 days in rodents (3,23). It is possible, therefore, that our studies were performed at a time when revascularization has occurred. If this is not the case, it is still likely, however, that despite improvements in islet function following full vascularization, that suboptimal islet β-cell insulin secretion would be observed due to the relatively low mass of islets transplanted. It is less likely that ongoing rejection contributed to the abridged islet function, as we did not observe lymphocyte infiltration surrounding or penetrating the transplanted islets (12).
We assessed insulin sensitivity by the minimal model of glucose kinetics to calculate the SI, a mathematical index of insulin sensitivity that has been extensively validated in rodents and multiple large animal models, including nonhuman primates (2,4). The SI data obtained from the IVGTT suggest that while islet mass is reduced, insulin sensitivity remains unchanged between the naive and transplanted animals and so is unaffected by rapamycin treatment. Nonetheless, the FFA and glycerol responses observed during the IVGTT provide further insight into islet function postislet transplantation. Insulin's well-described antilipolytic effects inhibit triglyceride breakdown to its products, FFA, and glycerol. In naive primates, we observed a rapid decline in FFA and glycerol levels soon after insulin concentrations rise during the IVGTT. In fact, FFA and glycerol levels remain near their minimum as glucose increases above basal levels. Only after insulin concentrations begin to fall do the FFA and glycerol levels rise. In contrast, neither the FFA nor the glycerol levels decline below basal levels in the transplant group. Therefore, it is possible that the islets of transplanted primates produce just enough insulin to control blood glucose but insufficient insulin to inhibit lipolysis in peripheral tissues. These findings are in contrast to those observed in human islet transplant recipients, who demonstrate the expected insulin-mediated suppression of FFAs (9,27). These distinct differences may potentially suggest that nonhuman primates exhibit a greater impairment in insulin-mediated inhibition of lipolysis over that of glucose disposal in the setting of impaired insulin secretion. On the other hand, these effects may be related to rapamycin therapy, as rapamycin has been shown to increase lipolysis and FFA release in 3T3-L1 adipocytes (31) and lead to increased triglyceride concentrations in human nonislet transplant recipients following conversion from calcineurin inhibitors to rapamycin for immunosuppression (33).
We demonstrate reductions in arginine-stimulated glucagon secretion that suggest the improved glycemic control following islet transplantation may be related in part to lowering of ambient circulating glucagon levels. The factors influencing glucagon secretion and maintenance of α-cell mass are quite complex. Our group recently showed a surprising reduction in α-cell mass following autoimmune mediated β-cell destruction (26). Many groups have reported that hypoinsulinemic diabetic patients manifest a paradoxical rise in glucagon secretion induced by secretagogues such as arginine; and that optimal insulin administration normalizes such abnormalities (17). Further, patients with insulinopenic diabetes but who also lack glucagon (e.g., postpancreatectomy) are less prone to ketoacidosis, despite the insulin deficiency (7). In addition, genetic mouse models lacking the insulin receptor in α-cells exhibit elevated and inappropriate glucagon secretion and subsequent hyperglycemia (16). Therefore, it is possible that transplantation of functional islets with a complement of all endocrine cell types through secretion of insulin, or other factors, restores or potentially enhances intraislet signaling within the endogenous islets of the recipient, leading to a reduction in α-cell glucagon secretion. Recent evidence has also shown that inhibition of the mammalian target of rapamycin (mTOR) pathway with rapamycin can reduce proliferation of α-cells (18). It is possible that in a setting of rapamycin monotherapy, the supratherapeutic rapamycin concentrations used in our studies, may have led to reductions in both α-cell mass and function. It is also possible that the mechanisms of reduced β-cell function following islet transplantation, such as revascularization and denervation of transplanted islets, may lead to impaired glucagon secretion.
In summary, we assessed several metabolic parameters using the nonhuman primate islet transplant model. These data suggest that although insulin independent, the imperfect glycemic control observed in islet transplant recipients is secondary to limited functional islet β-cell mass. We demonstrate no evidence of insulin resistance during rapamycin monotherapy in the early posttransplant period. Further, we suggest that the suppression of an inappropriate glucagon response to stimuli observed in T1DM patients and β-cell-deficient primates may play an important role in achieving improved glycemic control.
Footnotes
Acknowledgments
The authors wish to thank Monnie Choongkittaworn and Noelle Patterson for their technical support. We wish to thank Michael Quon and Alan Cherrington for their advice. We wish to acknowledge the excellent husbandry and care provided by the NIH Veterinary Resources Program, building 14D staff. The authors also wish to acknowledge support of the HHMI-NIH Research Scholars Program, the Benjamin and Mary Siddons Measey Foundation, the Charles H. Humpton, Jr. Endowment, and the Margaret Q. Landenberger Foundation to S.A.S., and support of the Schiffrin Family Foundation to M.R.R. A.E.S. and K.I.R. were supported by the intramural program of NIDDK/NIH.