
Abstract
Polycystic kidney disease (PKD) and nephronophthisis are common manifestation of ciliopathies. PKD is the most common genetic renal condition; it affects 12.5 million people worldwide. PKD is a great example of decades of translational research leading to the discovery of novel treatments and significant number of clinical trials. This review will concentrate on the basic molecular and cellular pathophysiology that led to the development of therapeutic targets for PKD.
Introduction
The most widely encountered renal ciliopathies include polycystic kidney disease (PKD), nephronophthisis (NPHP) and Bardet-Biedl, Joubert, and Meckel-Gruber syndromes. PKD refers to two specific genetic diseases, autosomal recessive polycystic kidney disease/congenital hepatic fibrosis (ARPKD/CHF, MIM *606702) and autosomal dominant polycystic kidney disease (ADPKD-OMIM *601313 and OMIM *173910). ARPKD/CHF and ADPKD have undergone decades of translational research resulting in the discovery of numerous therapeutic targets leading to a significant number of clinical trials. This review will provide a discussion of the basic molecular, and cellular pathophysiology that have led to the development of these therapies.
ARPKD/CHF is a dual organ disease characterized by cystic dilations of renal collecting ducts and hepatobiliary dysgenesis leading to varying degrees of congenital hepatic fibrosis [1, 2]. The incidence of ARPKD is estimated to be around one in 20,000 to one in 40,000 live births with a carrier rate of approximately 1/70 [3]. ARPKD is caused by mutations in PKHD1, which encodes fibrocystin or polyductin (FPC), a large protein with a single transmembrane spanning domain. Missense, frameshift, and truncating mutations have all been described [4], and nearly all patients are compound heterozygotes [5–7]. FPC colocalizes with polycystin-2 (PC2), the protein product of PKD2, at the basal bodies of primary cilia although the function of this complex is unknown.
ADPKD, the most common genetic renal disease occurs at a rate of 1/400 to 1/1000 live births [8] and affects 12.5 million individuals worldwide [9]. ADPKD classically presents with slow, progressive replacement of kidney tissue with fluid-filled cysts leading to end-stage renal disease (ESRD) most commonly by the sixth decade of life. ADPKD can manifest in utero, in infants, and in children and can be a significant cause of morbidity and mortality in this age group [10–12]. ADPKD is a heterogenic disease caused by mutations in two genes. PKD1 (OMIM *601313), encoding polycystin 1 (PC1) accounts for 80% of all ADPKD and PKD2 (OMIM *173910) responsible for 15% of ADPKD encoding PC2 [8]. The remaining 5% of ADPKD families do not map to either the PKD1 or PKD2 locus, suggesting the possibility of a third ADPKD gene; GANAB or DNAJB11 may account for some of these cases [13–15].
There is extreme allelic heterogeneity in PKD with over 1200 known pathogenic mutations in PKD1 (http://pkdb.mayo.edu). In contrast to PKD1, the PKD2 gene is much smaller and less complex and it has simpler features and structure [16]. However, a significant degree of allelic heterogeneity exists in PKD2 as well with over 200 pathogenic mutations identified to date (http://pkdb.mayo.edu). PC1 has a cytoplasmic C terminus that regulates multiple signaling cascades [17] and is also known to interact with PC2. PC2 is a calcium-permeable channel with six transmembrane spanning elements; both the amino and carboxy termini face the cytoplasmic compartment. Interaction with PC1 is required for full-length PC2 to leave the ER and reach the cilia [18, 19].
Pathophysiology of cyst formation
The process of cyst formation and growth is multidimensional and dynamic with multiple independent contributing components. PKD proteins are localized to apical and basolateral plasma membranes, desmosomes, focal adhesions, basal bodies, and primary cilium. In mouse models of PKD, cyst initiation and expansion are regulated by Pkd1 gene dosage [20–22]. Tubular epithelial cells have a threshold expression level of Pkd1 that is necessary to maintain normal tubular size and non-proliferative state. When the level of functional PC1 falls below the cystogenic threshold, the proliferation of tubular epithelial cells and cyst formation follows. Recent studies have demonstrated that the rate of cyst growth is regulated and can be adjusted by altering levels of functional PC1 [22]. Overexpression of Pkd1 can rescue a mutant Pkhd1. Hence PC1 dosage modifies the severity of both ADPKD and ARPKD.
The role of the primary ciliary structure in PKD cyst formation has recently been interrogated using conditional and inducible mouse models that allow structural ciliary genes (Kif3a and Ift20) and PKD genes (Pkd1, Pkd2, and Pkhd1), to be inactivated or deleted in a spatiotemporal manner, either alone or in combination. These studies specifically examined the functional role of PC1 and PC2 proteins on the cilia and revealed a cilia-dependent proliferation (cyst-promoting) pathway that is inhibited by a normal PC1/PC2 complex on the cilia [23, 24]. Furthermore, it is shown that Tubby-related protein 3 (Tulp3) and Daz interacting protein-1 (DZIP1L) impact the efficiency of the PC1/PC2 insertion into the cilia. Murine knock-outs of either Tulp3 and Dzip1l leads to collecting duct cysts [25, 26].
The focal nature of cyst formation in ADPKD adds additional complexity to the pathophysiology. Despite all cells in the body having the same germ-line mutation, only 5 to 10% of the nephrons become cystic. This puzzling fact led to the development of the two-hit theory which hypothesizes that a second mutation (i.e., a somatic mutation) is required for the initiation of cyst formation. Hence ADPKD behaves as a recessive disease at the molecular level. Substantial evidence exists to support this theory [27–30] however, additional factors have been shown to influence disease progression and severity [31]. These include: 1) the developmental timing of PKD1 inactivation [32–35]; 2) reduction in functional PC1 levels [20, 37]; and 3) one cyst among neighboring cells and nephrons can create a toxic environment resulting in a “snowball effect” leading to cyst development in neighboring nephrons [38].
The complex interactions between the three PKD proteins make gene-targeted therapies a daunting task. As a result, most therapies at present are aimed at interrupting abnormal signaling pathways driving cyst growth and associated interstitial inflammation and fibrosis.
Targeting the cystic phenotype
During cyst formation normal renal epithelial cells change from mature, differentiated, nonproliferative, absorptive cells to partially de-differentiated, secretory cystic cells characterized by specific polarity defects and increased rates of proliferation [8]. Cyst formation is associated with proliferation of cells lining the expanding cysts, fluid secretion into cysts that have lost their connection to the nephron and remodeling of the extracellular matrix surrounding the expanding tubules.
Multiple signaling molecules and pathways have been identified in the phenotypic change of normal tubular epithelial to the cystic phenotype [8]. The precise mechanisms by which defective PKD proteins alter these signaling pathways are not fully understood. Despite this knowledge gap, key pathogenic features common in ARPKD and ADPKD have been identified and form the basis of current therapeutic interventions and clinical trials. These features include: Altered intracellular cAMP levels coupled with decreased intracellular calcium leading to increased proliferation and fluid secretion. Abnormalities in expression, localization, and activity of the epidermal growth factor (EGFR) family of receptors and ligands (EGFR axis) leading to increased proliferation. Increased activity of cSrc, a critical mediator of cross-talk between the EGFR axis and G-protein-cAMP pathways which is also required for STAT3 activation by the PC1 tail. Alterations in cell-cell adhesion, and cell-matrix interactions. Chronic inflammation that leads to progressive fibrosis.
Clinical trials to date have been largely based on targeting these aberrant, complex cellular signaling elements. Therefore, a brief discussion of each along with the results of clinical trials targeting these elements to date will follow. Figure 1 depicts major signaling pathways that comprise the phenotype of the cystic cell. The main proliferative pathways are shown, but other entry points into these pathways are possible and likely. A list of current and completed clinical trials can be found at http://clinicaltrials.gov/
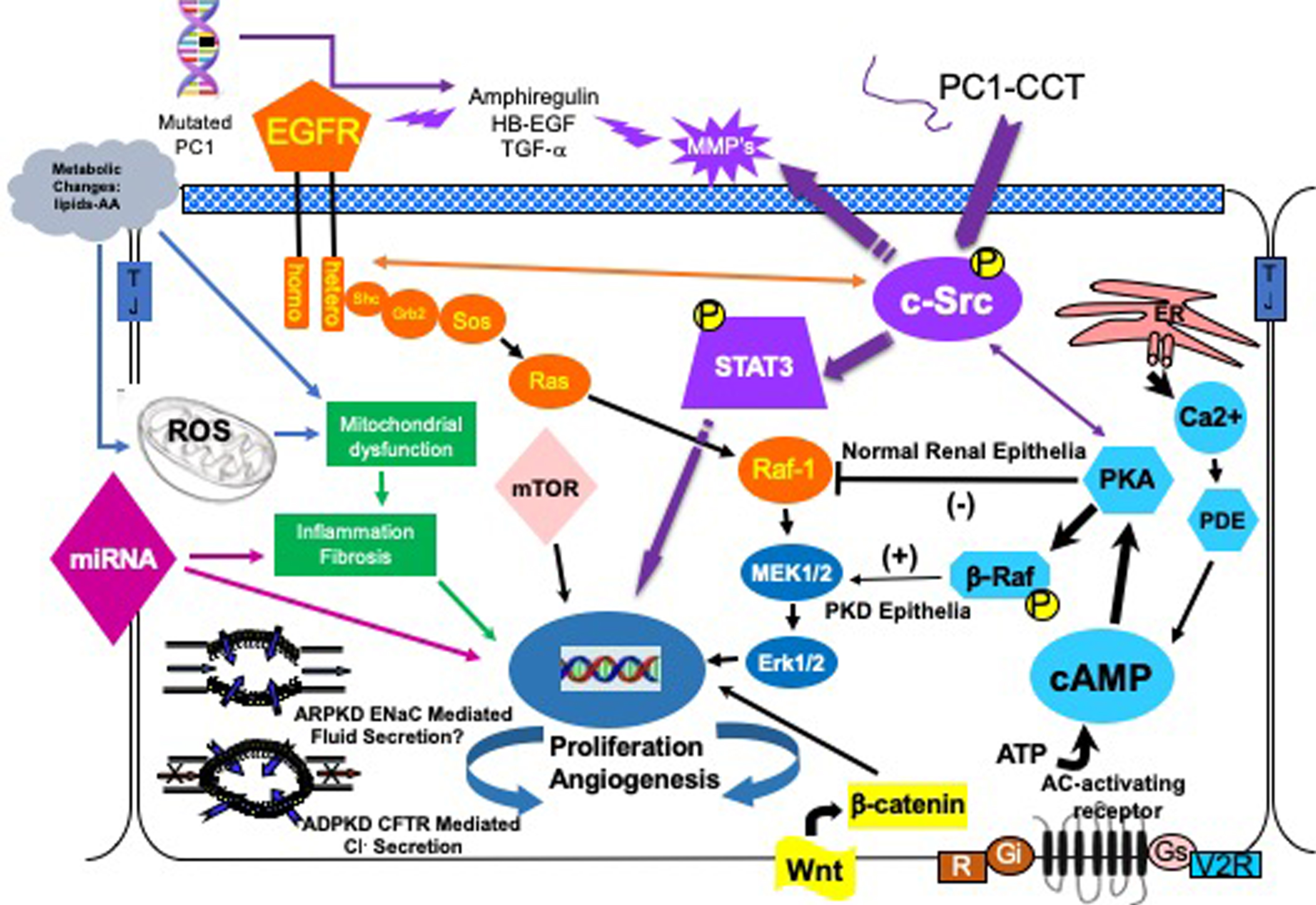
The cystic cellular phenotype. This cartoon is an abridged composite of the abnormal signal transduction pathways reported to be active in PKD. Two main conduits that lead to unchecked proliferation are 1) the EGFR axis (orange), and 2) a G-protein axis (aqua blue path) that leads to increased cAMP and a switch in the phenotypic response of renal epithelia to cAMP. The pathways suggest a possible pathogenic process as follows: in ADPKD (purple path) a mutated PC1 leads to increase amphiregulin, activating EGFR, resulting in increased cSrc phosphorylation; in both ARPKD and ADPKD, cSrc activation alters the cellular response of cAMP resulting in increased proliferation and secretion; in addition, the cytoplasmic tail, PC1-p30, is overexpressed. PC1-p30 interaction with Src causes Src-dependent activation of STAT3 by tyrosine phosphorylation; and in ARPKD, an apical EGFR becomes activated resulting in reciprocal phosphorylation of the nonreceptor tyrosine kinase cSrc leading to increased cAMP-mediated, and EGFR driven proliferation. Increased activity of EGFR and increased cAMP amplify the activation of Src-STAT3 pathway. There are multiple entry points that can augment these two main pathways including the WNT/B-Catenin pathway, metabolic changes leading to the generation of reactive oxygen species and mTOR activation and abnormalities of miRNAs.
The cAMP-dependent pathway, or the adenylate cyclase pathway, is a G-protein-coupled receptor signaling pathway. In normal renal epithelia, increased intracellular cAMP results in a signal to reduce epithelial proliferation, but in cystic renal epithelia this blocking signal is bypassed, and cAMP is needed to maintain a pro-proliferative phenotype. Both low intracellular calcium level, (theoretically due to a defective PC1/PC2 calcium channel) and cSrc dependent phosphorylation of β-Raf, allows the cell to bypass Raf-1 and increase ERK phosphorylation to stimulate cell proliferation [39, 40]. Increased cAMP in a cell with an active MAPK pathway also changes the cell from an absorptive to a secretory cell which contributes significantly to the progressive cyst enlargementin ADPKD [41].
Intracellular cAMP is pharmacologically reduced by vasopressin R2 receptor antagonists such as tolvaptan (Samsca or Jinarc®) or somatostatin (and its long-acting analogs such as lanreotide) that inhibit adenylate cyclase [42, 43]. Tolvaptan has undergone extensive human clinical trials. Tolvaptan has proven to slow the increase in total kidney volume and reduce the rate of decline in loss of renal function in ADPKD patients [44, 45]. However, in patients with later-stage ADPKD, treatment with lanreotide compared with standard care did not slow the decline in kidney function over 2.5 years of follow-up [46]. Jinarc® (Tolvaptan) has been approved in Europe, Japan, Canada and Australia for treatment of ADPKD in adults. Tolvaptan has recently been approved in the USA under the trade name JYNARQUE®. Due to FDA concerns regarding potentially serious adverse liver events, in the USA, JYNARQUE is only available through a restricted distribution program called the JYNARQUE Risk Evaluation and Mitigation Strategy designed to closely monitor patients for liver or other adverse events(www.fda.gov/safety; https://www.jynarque.com). Tolvaptan is not currently recommended for use in children due to concerns regarding the risk of fluid imbalance although clinical trials are underway in Europe [NCT02964273 Belgium and Italy].
EGFR (ErbB) axis
EGFR axis consists of the epidermal growth factor/ErbB family of receptors and ligands. A large body of evidence indicates that one or more members of the ErbB receptor family play a role in renal cystic epithelial proliferation. This includes EGFR, as well as the related receptors, ErbB2 and ErbB4 [47–50]. In human ADPKD and ARPKD and every rodent model of PKD published to date, cystic kidneys display characteristic abnormalities in EGFR-axis. Inhibition or reduction of EGFR activity through genetic manipulation [51] or administration of tyrosine kinase inhibitors [52, 53], reduces cyst formation and enlargement in animal models.
Microarray profiling in human PKD1 cystic cells demonstrated the EGF/ErbB family receptor ErbB4, is a major driver of cyst growth in ADPKD. ErbB4 levels may also serve as a prognostic biomarker for disease progression [50]. Recent studies provide a direct link between PC1 and PC2 proteins and the EGFR axis; EGF reduces the threshold of PKD2 activation by releasing it from PIP(2)-mediated inhibition [54].
cSrc
cSrc is a critical intermediate that integrates signals between two major proliferative pathways in a cystic epithelial cell, the EGFR axis and cyclic AMP pathway [55] (Fig. 1). Also, a proteolytic fragment of the PC1 cytoplasmic tail, PC1-p30, interacts with cSrc and activates STAT3 stimulating further increase in proliferation [56–58]. Src activation of STAT3 is amplified by increase of both EGFR axis activity and cAMP levels resulting in a self-perpetuating cycle that accelerates proliferation [58]. Further, a mutated PC1 increases amphiregulin expression, resulting in activation of the EGFR axis and reciprocal activation of Src which amplifies inputs from EGFR, and cAMP signaling pathways to activate STAT3 [58]. In a clinical trial on ADPKD patients, Bosutinib, a cSrc inhibitor, (ClinicalTrials.gov Identifier NCT01233869) was very effective in reducing total kidney volume but did not reduce the decline in renal function [59].
mTOR
The mammalian target of rapamycin (mTOR) pathway integrates signals from growth factors (including EGFR), G-protein coupled receptors (which generate cAMP), cellular energy levels, and stress conditions to stimulate protein synthesis and cell growth through phosphorylation of S6K1 and eIF4E [60, 61]. In human ADPKD, ARPKD and a variety of animal models, cyst-lining epithelium reveals an increased activity of mTOR [61–64]. The mTOR inhibitors rapamycin and everolimus, therefore, have been tested in human clinical trials and were found to be ineffective in slowing total kidney volume or the progressive loss of renal function [65, 66].
In an effort to explain the disappointing results of the mTOR trials, it has been suggested that ineffective renal concentrations of mTOR inhibitors may be responsible for the unsatisfactory clinical results. Based on the finding that cystic cells express folate receptors, Shillingford et al. conjugated folate to the mTOR inhibitor rapamycin [67] and showed that in a Pkd1 animal model when rapamycin was conjugated with folate, target specificity was increased and the effectiveness of mTOR inhibition was dramatically improved [67]. This concept of linking a therapeutic agent with a protein that has a receptor on the cell of interest is not new but is one that deserves further investigation in the field of PKD therapy. In ARPKD, cystic lesions occur largely in collecting ducts. The ability to specifically target collecting duct cells allows for lower systemic exposure of the therapy while increasing the concentration at the site of disease.
Multi-kinase inhibitor
Multi-kinase inhibitor therapy takes advantage of a compound that inhibits the activity of multiple proteins known to be aberrantly active in cystic disease process. One such compound Tesevatinib (TSV) previously known as KD-019, is a unique multi-kinase inhibitor that targets EGFR, c-Src and KDR (VEGFR2). TSV was effective in slowing the proliferation of renal epithelia and cyst growth, progression of fibrosis and the loss of renal function in the BPK and orthologous PCK rat models of ARPKD [68].
TSV is currently being tested on adults in two ADPKD Phase II-III clinical trials (ClinicalTrials.gov Identifier NCT01559363 and NCT03203642). TSV is also the first specific disease-targeted therapy to receive FDA approval for a clinical trial in pediatric ARPKD (ClinicalTrials.gov Identifier NCT03096080). All three clinical trials are still in progress.
20-HETE
Metabolic changes are known to occur during the development of ARPKD and ADPKD. Bioactive lipids such as 20-HETE, a P450-produced metabolite of arachidonic acid was shown to correlate with eGFR and TKV in ADPKD patients [69]. Inhibitors of 20-HETE have been effective in reducing cyst formation in animal models of PKD [70–72]. Therefore, inhibition of 20-HETE production may be a potential therapeutic agent for slowing PKD progression. In addition, urinary 20-HETE levels may serve as a non-invasive biomarker of disease severity [69].
Future approaches
miRNA
miRNAs are short non-coding RNA molecules (∼22 nucleotides) that regulate gene expression. There are emerging data that support the integral role of miRNAs in the pathogenesis of PKD [73–76]. Recent studies in human ADPKD have linked a single miRNA to the progression of disease [50], and more recently a unique interstitial miRNA signature was found to play a pivotal role in the pathogenesis of renal fibrosis in an orthologous animal model of ADPKD [77]. Inhibitors of miRNA are powerful candidates for therapeutic intervention against various pathological conditions. One advantage of such therapies is the small size that makes it easier to get to target disease tissue [78]. Pharmaceutical companies are actively pursuing the development of miRNA-based therapies for numerous human diseases [79].
Targeting cilia
The primary cilium has an exceptionally high density of receptors on its membrane that are important for sensing and transducing extracellular stimuli. Moreover, the primary cilium exists as a separate cellular compartment from the cytosol, providing for unique spatial and temporal regulation of signaling molecules to initiate downstream events. Many cilia proteins are found at other sites and have non-ciliary functions. Therefore care must be exercised in attributing biological effects of a protein solely to the function of the cilia. The complexity of the spatial and temporal integration of proteins into the cilia must certainly be important in the proper function of the cilia and the efficiency in which signals are transmitted to the cell. Proteins and mechanisms that impact the efficiency or order of protein trafficking would theoretically be modifiable events and as such would present novel therapeutic targets.
Recent studies of cilia structure and function have identified proteins of interest, such as Tulp3 and Dzip1l, that may be targeted in PKD due to their ability to increase the efficiency of PC1/PC2 integration into the cilia. As the molecular biogenesis of the cilia becomes further characterized, opportunities to alter the efficiency or function of the cilia may become part of the therapeutic arsenal for PKD and other ciliopathies.
Conclusion
The clinical trials to date have shown that targeting a single molecule of an aberrant pathway in PKD has been ineffective in preventing the loss of renal function. In the short term, the most promising therapies will likely target key signaling intermediates that appear to integrate multiple pathways. These might include Src, (Fig. 1) [55, 58] and/or a combination therapy approach where multiple compounds are used to target multiple pathways simultaneously or a single compound is used to target multiple pathways such as a multi-kinase inhibitors (MKI) like tesevatinib (TSV).
Rational therapies will require knowledge of both the extent of disease and the rate of disease progression. Therapeutic targets will be stage-specific and change as the disease progresses. Therapies will become increasingly focused on treating PKD in early childhood where they are likely to provide the greatest long-term benefit. Two clinical trials for children with PKD are underway: One for children with ADPKD [Tolvaptan [NCT02964273 Belgium and Italy] and one for the use of TSV in ARPKD (ClinicalTrials.gov Identifier NCT03096080). Therapies that target abnormal signaling pathways will need to be carefully tailored, so that pathway activity is reduced to normal basal levels rather than eliminated. Promising compounds will be modified to direct the molecule to the site of disease, making these therapies highly specific with low levels of toxicity [67]. Epigenetic and dietary factors that slow or accelerate the progression of PKD will likely be discovered, and adherence or avoidance of such factors may slow the rate of progression and eliminate the need for pharmacological intervention or renal replacement therapy for some.
The importance of gene dosage, modifier genes and somatic mutations in the clinical course of a disease in an individual provide a compelling rationale for personalized medicine. Personalized medicine will mean tailored approaches that modulate functional gene dosage and consider not only individual genotypes but also the response of the kidney to disease and the unanticipated response of the kidney to therapy. This unanticipated response to therapy may be active in tolvaptan treatment which reportedly results in increased shedding of HB-EGF [80]. The results of this shedding are unknown but would suggest that combining Tolvaptan with a tyrosine kinase inhibitor may improve clinical responses. Therapeutically improving the efficiency of PKD1 processing in the ER (neutral alpha-glucosidase designated AB, (GANAB)) or improving the efficiency of its insertion at the cilia (TULP3) may have significant impact on the disease expression.
Conflict of interest
The authors declare that they have no conflicts of interest concerning research, authorship and publication of this manuscript.
Footnotes
Acknowledgments
This work was supported, in part, by the Lillian Goldman Charitable Trust; the Amy P. Goldman Family Trust, the EllsworthFamily Fund; and the Children’s Research Institute at the Children’s Hospital of Wisconsin.