
Abstract
Parkinson’s disease (PD) is a common neurodegenerative disease whose pathogenesis remains unknown. TMEM230 gene, encoding a transmembrane protein in secretory and recycling vesicle, has been recently identified as a novel disease-causing gene of autosomal dominant PD with Lewy pathology and typical clinical symptoms. Although its mutation and variants seem to be rare in PD patients, functional studies have indicated that TMEM230 protein probably plays an important role in secretory and recycling pathway and may be involved in Lewy pathological mechanism. Here we summarize current genetic and functional reports about TMEM230 and focus on its relation with PD.
INTRODUCTION
Parkinson’s disease (PD) (OMIM 168600) is the second most common neurodegenerative disease after Alzheimer’s disease (AD), with incidence rate of 0.014% per year in total population and 0.16% per year in people over the age of 65 in high-income countries [1]. The typical clinical symptoms of PD include progressive bradykinesia in combination with rest tremor, rigidity, postural instability, and numerous non-motor symptoms [2, 3]. The loss of dopaminergic neurons in the substantia nigra pars compacta (SNpc) and the Lewy pathology including Lewy bodies and Lewy neurites have been identified as pathological features of PD [2]. PD was once thought to be a nongenetic disease and mainly caused by environmental factors, but as a result of advances in genetic research, this notion has gradually changed and there is growing recognition of genetic mechanisms in the pathogenesis of the disease [4]. To date, at least 23 disease-causing loci and 19 genes have been reported in monogenic PD pedigree, though the list of risk-associated genes is steadily growing [3, 5]. PD is now viewed as a complex neurodegenerative disorder resulting from genetic, environmental and other, yet unknown factors [4].
Unlike sporadic PD which accounts for about 90% of total PD cases, a considerable number of patients with monogenic form of PD have been found to have young-onset of symptoms, atypical clinical features, and lack Lewy pathology [1, 7]. Only a few of PD causing mutations had been reported to be related to clinically typical PD with Lewy pathology [6]. Recently, Deng et al. reported that transmembrane protein 230 gene (TMEM230), may play a pathogenic role in a rare autosomal dominant PD (ADPD) with typical motor features and Lewy body pathology [8]. This finding provides new insights into the pathogenesis of PD-related neurodegeneration. The primary aim of this article is to review current genetic and functional data about TMEM230 and suggest how the discovery of this disease-causing gene can lead to pathogenesis-targeted therapy for PD.
PARK21 AND THE TMEM230 GENE
In 2014, a large Canadian Mennonite family with a PD phenotype was reported with c.2564A > G (p.N855S) variant in the DNAJC13 gene, located on chromosome 3q22 [9]. But this variant did not fully cosegregate with the disease as demonstrated by its presence in one unaffected individual died at the age of 87 years and absence in two PD cases and one parkinsonism case with progressive supranuclear palsy pathology, who belong to different branches and couldn’t be explained as sporadic cases because extremely low possibility (<10- 3) [8]. The causal gene locus was termed as PARK21 (OMIM 616361) in Online Mendelian Inheritance in Man (OMIM) according to the chronology of identification of the disease-causing gene loci. However, in 2016, Deng and colleagues proposed that TMEM230 c.422G > T (p.R141L) mutation, mapped to chromosome 20p13-p12.3, was the pathogenic mutation of PD in the same family with 13 available patients [8]. Though a TMEM230 mutation–freed patient with atypical parkinsonism phenotype which was still mild after 23 years of disease progression was evidenced, the TMEM230 mutation was the best genetic explanation for PD in this family under current known data, which may be explained by other conditions such as environmental or inconsistent genetic factors [10, 11]. The Canadian Mennonite family of mixed European ancestry included 14 enrolled family members with ADPD, mean age at onset of 67.0 years, and typical presentation of late-onset, levodopa-responsive PD [8, 12]. These patients had rigidity, bradykinesia, and rest tremor (in 57% patients), and dementia was present in 21% of cases [13]. Additionally, neuronal loss in substantia nigra and nucleus basalis, and Lewy bodies were found in brainstem at autopsies including α-synuclein stains, performed in three of cases [10, 13].
The TMEM230 gene, also called as chromosome 20 open reading frame 30 (C20orf30), covers a genomic region of about 13.2 kb with five exons [14]. TMEM230 mRNA expression is high in many tissues, including several regions of nervous system, such as midbrain, cerebellum, neocortex and spinal cord [8, 15]. Its four mRNA transcriptional variants encode two protein isoforms: the isoform-1 of 183 amino acids and the isoform-2 of 120 amino acids [8]. The isoform-2 accounts for more than 95% of total protein isoforms in humans, and presents alone in species spanning zebrafish to most mammals [8]. The highly conserved amino acid sequence of isoform-2 contains two transmembrane segments, with N-terminal and C-terminal regions exposing to the cytosol [8]. There is no other known protein with sequence identical or similar to TMEM230.
TMEM230 VARIANTS IDENTIFIED IN PD
Three other PD-associated TMEM230 variants, including two variants (p.Y92C and p.*184Wext*5) which were found through analyzing 832 North American PD cases and one variant (p.*184PGext*5) which were detected by 9 PD cases of 7 families from China were reported in original Deng et al.’s study [8]. The asymptomatic carriers with p.Y92C and p.*184PGext*5 variants suggested incomplete penetrance, similar to LRRK2 p.G2019S variant [8, 16].
Mutation/variants associated with PD detected in coding region of the TMEM230 gene
FPD, familial PD; SPD, sporadic PD; Het, heterozygous; Hom, homozygous; MAF, minor allele frequency; ExAC, Exome Aggregation Consortium database; gnomAD, Genome Aggregation Database. Data in reference 31 was not extracted due to the lack of detailed information [31].
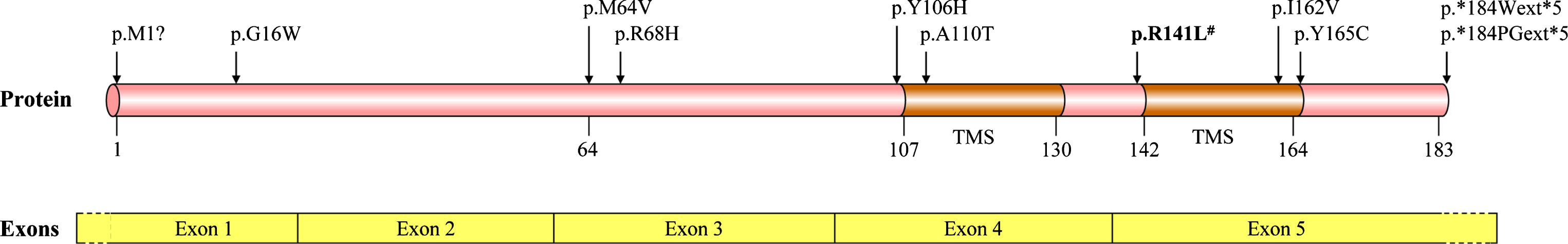
Missense mutation and variants associated with PD detected in TMEM230 coding regions. # The initial mutation detected in a large family with autosomal dominant and Lewy pathology confirmed PD. TMS, transmembrane segment.
THE POTENTIAL PATHOLOGICAL MECHANISM OF TMEM230 IN PD
The TMEM230 protein is localized to vesicle structures in human SH-SY5Y cells, and the mouse ortholog distributes to the same subcelluar structures in brain neurons including dopaminergic neurons in the substantia nigra [8]. These vesicle structures predominantly co-localize with STX6, a protein mainly enriched in the trans-Golgi network (TGN), and with vesicular monoamine transporter type 2 (VMAT2), vacuolar protein sorting-35 (VPS35), Rab5a and Rab11a proteins, suggesting that TMEM230-positive vesicles are involved in the function of synaptic vesicles and recycling of endosomes [8]. Thus, TMEM230 appears to play an important role in many cellular functions, including synaptic vesicles trafficking, retromer trafficking, secretory autophagy and Golgi-derived vesicle secretion. As such, it shares in pathogenic pathways implicated other PD-causing genes, such as the SNCA, LRRK2, VPS35 and PINK1 [32].
Interaction with SNCA
Mutations in synuclein alpha (SNCA) gene, which encodes α-synuclein protein, the key component of Lewy body inclusions, have resulted in ADPD [6, 33]. As the first PD-related gene identified and labeled PARK1, the typical PD clinical phenotype of patients was also associated with characteristic Lewy body pathology [6, 33]. Although the physiological function of α-synuclein protein remains enigmatic, mounting evidence suggests a regulatory function in synapse, such as vesicle trafficking, synaptic vesicle pool maintenance and neurotransmitter release [33]. TMEM230 protein was detected in α-synuclein-positive Lewy bodies and Lewy neurites both in sporadic PD and dementia with Lewy bodies (DLB) cases [8]. Similar to α-synuclein, the TMEM230 protein was observed in the synaptic vesicle pool region in the rat brain neuron presynapse [8, 34]. Expression of PD-related TMEM230 variants resulted in significantly slower movement of synaptic vesicles and increased α-synuclein protein level compared to wild-type protein possibly due to impairment of autophagy-mediated clearance [8, 32]. In addition, the Rab8a protein whose function is connected with TMEM230, also interacts with α-synuclein, and its overexpression reduces α-synuclein-induced toxicity in vitro and improves α-synuclein-induced behavioral defects in fruit flies [32, 35]. Intriguingly, tmem230a, the zebrafish ortholog of human TMEM230, could affect angiogenic blood vessel growth though Delta/Notch signaling pathway [36], which may be involved in neurodegenerative disease and reduced by overexpressive or mutant α-synuclein protein [37, 38].
Interaction with LRRK2
Leucine-rich repeat kinase 2 (LRRK2) mutations represent the most common genetic cause of ADPD and nearly half of LRRK2-related PD cases had Lewy bodies [6, 39]. LRRK2 protein has been found to phosphorylate several members of Rab family which plays a key role in all forms of intracellular vesicular trafficking [40]. The Rab8a protein is one of substrates of LRRK2, and its phosphorylation may be increased 2-3 fold by LRRK2 p.G2019S mutation [40]. Thus LRRK2 kinase activity-dependent phosphorylation may lead to deficits in cell polarization, neurite outgrowth and directed migration [41]. The Rab8a-mediated secretory vesicle and retromer trafficking were impaired when TMEM230 lost function, similar to lack of LRRK2 protein [32]. This suggests that TMEM230 and LRRK2 may share Rab8a-mediated vesicle trafficking pathway in development of PD and Lewy pathology. Additionally, Notch signaling pathway which may be associated with TMEM230 was also regulated by LRRK2 through endosomal pathway [36, 42].
Interaction with VPS35
VPS35, a causal gene linked to ADPD, encodes a subunit of retromer complex [43]. The TMEM230 protein partially co-localizes with VPS35 protein and both regulate retromer trafficking function [8]. The expression of TMEM230-R141L mutant protein changed VPS35 and itself from perinuclear to punctate cytoplasmic distribution [32]. However, only one VPS35-PD autopsy report showed no immunostaining for α-synuclein and there was no neuronal loss or intraneuronal inclusions in the cortex and basal ganglia; the substantia nigra tissue was not available [44]. Further studies are warranted to clarify the similarities and differences between TMEM230 and VPS35 in pathogenesis of PD.
Interaction with PINK1
Many mutations of phosphatase and tensin homolog-induced putative kinase 1 (PINK1) gene have been identified in different families with autosomal recessive PD [45]. One early-onset PD patient with two compound heterozygous PINK1 mutations was reported to have neuronal loss and Lewy pathology in the SNpc [46]. This gene encodes PINK1 protein, a serine/threonine protein kinase whose activation caused phosphorylation of Rab8a at residue of serine 111 and significantly impaired Rab8a activation [47]. Further studies of Rab8a-involved pathway may help to elucidate the association between TMEM230 and PINK1 in pathogenesis of PD.
In summary, there is a growing body of evidence that TMEM230 protein and its interaction with Rab8a, SNCA, LRRK2 and PINK1 may lead to PD-related neurodegeneration (Fig. 2).
The potential pathological mechanisms and associated proteins of TMEM230. TGN, trans-Golgi network; TMEM230, transmembrane protein 230; PINK1, phosphatase and tensin homolog-induced putative kinase 1; LRRK2, leucine-rich repeat kinase 2; VPS35, vacuolar protein sorting-35.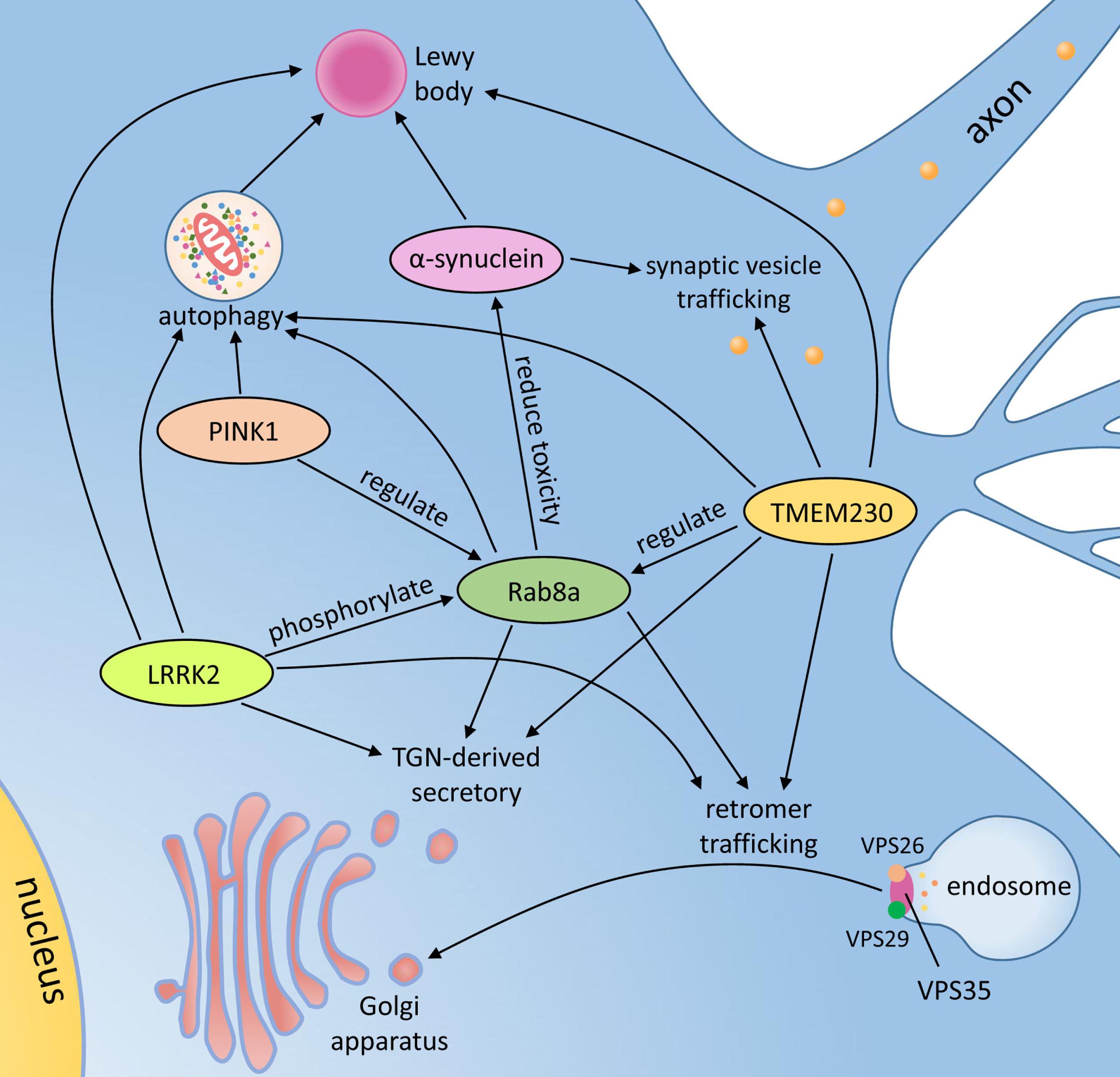
THE POTENTIAL ROLE OF TMEM230 IN OTHER DISEASES
The TMEM230 gene may be potentially related with other neurodegenerative diseases with Lewy pathology such as DLB and AD, multiple system atrophy (MSA) [48, 49]. In AD patients, the TMEM230 protein was increased in hippocampal neurons and aggregated in granulovacuolar and dystrophic neurites, two prominent pathological features of AD [50]. No MSA-risk variants have been found in the TMEM230 gene in 110 cases of MSA [51]. Furthermore, He et al. did not find stop codon variants in the TMEM230 gene in 200 Chinese patients with essential tremor [26].
CONCLUSION
Even after exciting acceleration of PD research during the past 50 years since the discovery of levodopa, the pathogenesis of this complex disorder remains enigmatic [2]. Notable discoveries, especially the advances in genetics of PD in recent 20 years have greatly changed our understanding on etiology and pathogenesis of PD [1]. It seems increasingly clear that PD is a highly complex neurological disease with heterogeneous clinical presentation, variable pathological features, and multifactorial causes [52]. Reveal of the association of phenotype-genotype, especially analysis of protein interaction network involving in Lewy body-confirmed PD-related genes, which highly mimics idiopathic PD, will help to understand the main underlying pathogenic mechanism of this complex disorder [52]. But only a few of PD-related mutations have been associated to PD with Lewy body pathology which remains a core feature of most PD cases, without precise mechanism known [1, 6].
Only a few PD-related TMEM230 variants could not cover up its significance in discovering pathogenesis of PD. Copy number variations including duplication and triplication in the TMEM230 gene, perhaps share a similar mechanism resulting in PD with dementia as the SNCA gene [53], as well as its epigenetic or non-coding regulatory factors, cannot be ignored in future studies. The application of quantitative PCR, digital PCR, whole genome sequencing and epigenetic strategies may help to identify more pathogenic mechanisms involving TMEM230 in PD, especially phenotype with dementia and other neurodegenerative disorders. Future research should focus on development of TMEM230 genetic animal models to better understand the role of TMEM230 in pathogenesis of neurodegeneration. These studies may also provide insight into potential treatment and prevention of PD and related disorders.
CONFLICT OF INTEREST
The authors have no conflict of interest to report.
Footnotes
ACKNOWLEDGMENTS
H.D.’s research was supported by the National Key Research and Development Program of China [grant number 2016YFC1306604], the National Natural Science Foundation of China [grant number 81670216], the Natural Science Foundation of Hunan Province [grant numbers 2015JJ4088, 2016JJ2166 and 2017JJ3469], Grant for the Foster Key Subject of the Third Xiangya Hospital of Central South University [Clinical Laboratory Diagnostics], and the New Xiangya Talent Project of the Third Xiangya Hospital of Central South University [grant number 20150301], China. J.J. was supported by the Parkinson Foundation and Michael J. Fox Foundation for Parkinson Research.