
Abstract
BACKGROUND:
Muscle stretch reflexes are widely considered to beneficially influence joint stability and power generation in the lower limbs. While in the upper limbs and especially in the muscles surrounding the shoulder joint such evidence is lacking.
OBJECTIVE:
To quantify the electromyographical response in the muscles crossing the shoulder of specifically trained overhead athletes to an anterior perturbation force.
METHODS:
Twenty healthy male participants performed six sets of different external shoulder rotation stretches on an isokinetic dynamometer over a range of amplitudes and muscle pre-activation moment levels. All stretches were applied with a dynamometer acceleration of 10,000
RESULTS:
Consistent reflexes were not observed in all experimental conditions. The reflex latencies revealed a significant muscle main effect (F (2,228)
CONCLUSION:
Short latency muscle reflexes showed a tendency to an anterior to posterior muscle recruitment whereby the main internal rotator muscles of the shoulder revealed the most consistent results.
Keywords
Introduction
High-speed over-head throwing requires a complex sequence of individual joint movements involving the whole body. Several investigations examined which joints and joint-specific angular motions are predominantly involved in the generation of high projectile velocities [1, 2, 3, 4]. Roach et al. [5] and Roach and Lieberman [6] promoted the hypothesis of the ‘kinetic chain’ which transfers momentum, generated in adjacent, connected body segments from joint to joint. The resulting interaction moments are evidence of high angular velocities during throwing in line with velocity-dependent torso rotation and shoulder internal rotation [2]. Studies that analysed muscular activity in throwing and related tasks indicate high muscle activity during internal shoulder rotation throughout the acceleration phase of the movement [7, 8]. However, some findings emphasized that only approximately 50% of the generated shoulder power can be explained by muscle activation patterns [5]. First explanatory approaches suggested that the storage and release of elastic energy also cause significant contribution to power enhancement within the upper body due to a ‘pre-stretching’ of numerous throwing related muscles just prior to their activation [9]. The stretch shortening cycle (SSC) of muscle function has been biomechanically analysed by several investigators over the last decades. Convincing evidence indicates that a SSC results in an enhancement of performance within the final phase of movement (concentric phase) when compared to isolated concentric action [10, 11]. However, so far most investigations have focused on the lower limbs. As described by Roach et al. [5] the posture of the arm at the end of the ‘cocking-phase’ (time of maximum external shoulder rotation) passively stretches elastic elements and therefore enables elastic energy storage and release at the shoulder. The exact magnitude of the rapid stretch contribution to power enhancement still remains largely unknown, especially within the upper extremity.
Former research has outlined an existing reflex arc within the feline model with an extremely short latency (2.7–3.1 ms) between the shoulder capsule and the internal rotator muscles of the shoulder evoked by electrical stimulation of the shoulder capsule [12]. In accordance with these findings the existence of various proprioceptors such as free nerve endings and Golgi mechanoreceptors in the human glenohumeral joint has been reported [13, 14]. Purposeful investigations of the monosynaptic short latency stretch reflex (SLR) indicated that it has considerable importance in stiffness regulation of the muscle, which would consequently enhance joint moments and power during SSCs movements [15, 16]. However only a few studies have used a joint perturbation to simulate specific tasks in the upper extremity which is associated with rapid angular motions of the glenoid joint. An exception was a study by Latimer et al. [17] analysing muscle response to an anterior perturbation force. Observed latencies ranged from 110 to 220 ms whereby the anterior muscles responded first followed by the posterior muscles. Furthermore, Myers et al. [18] demonstrated that muscle reflexive response considerably relied on muscle pre-activation which was expressed in a latency shortening. However, the results of these investigations were in disagreement to the findings of Roach et al. [5]. For an elite thrower up to 70% of the achieved projectile release velocity is developed in the last 100 ms of the movement [19]. The actual arm acceleration of a pitching motion which is initiated at the point of maximal external shoulder rotation and ends at ball release took only 42–58 ms of the total pitch temporal sequence (foot contact to ball release: 145
The purpose of the current research was (i.) to examine the capability of isokinetic dynamometry for the evaluation of muscular stretch reflexes in the muscles surrounding the shoulder and (ii.) to find evidence for SSC mechanisms at the shoulder to be similar to those described for the lower limbs [21]. We hypothesised that (a) muscle pre-activation moment and stretch amplitude would significantly influence reflex latency, reflex amplitude and muscle activation duration in terms of the stretch, (b) that the clavicular part of the pectoralis major muscle and the anterior part of the deltoid muscle in their function as major internal rotators of the shoulder would show the fastest and most consistent response to the anterior perturbation by the dynamometer as formerly reported in the literature [17].
A subject positioned in the dynamometer during familiarisation. Left: Starting position (90/90/90-position). Right: iMER (position of individual maximal external shoulder rotation).
Participants
Twenty healthy right-handed men (age: 22.9
Experimental protocol
The study consisted of one familiarisation and one test session. Subjects were seated on an isokinetic dynamometer (IsoMed2000, D&R GmbH, Hemau, Germany) in an upright position. A special back pad was used to stabilize the left (non-dominant) shoulder and ensure a sufficient abutment for the moving dominant arm throughout the application of the stretches. The right (also the dominant in all subjects) arm was 90
We determined maximum isometric moment of force (MVC) of the pectoral muscle, the deltoid muscle and the latissimus dorsi muscle from at least 3 trials of maximum isometric contractions of each subject in the 90/90/90-position with 2 minutes break between trials. The highest moment value during a steady plateau was used as MVC.
To minimise the risk of overstrain the individual maximum external stretch amplitude (iMER) for each participant was determined within the familiarisation. We used the same stretch velocity as in the test session (150
Experimental conditions. The order of stretch conditions was randomized
Experimental conditions. The order of stretch conditions was randomized
iMER: individual maximal external shoulder rotation, MVC: maximum voluntary contraction.
During the test session external shoulder rotation stretches were applied using the formerly determined individual maximal stretch amplitudes (50% of iMER and 100% of iMER) and three different pre-activation moment levels (0% MVC, 25% MVC and 50% MVC) according to the moment values of the MVC-test (Table 1). This set-up results in six different testing conditions. For each condition a minimum of 10 trials was accomplished with a rest period of 10 s between trials to avoid habituation and fatigue of the response [23] and possible influence of post activation depression [24]. Each stretch was applied with a dynamometer acceleration of 10
Surface EMG (sEMG) activity of the primary internal rotator muscles of the shoulder was recorded at a sampling frequency of 2000 Hz via an A/D board (National Instruments, Austin, USA) and Templo 7.1 Software (Contemplas GmbH, Kempten, Germany). Data were stored on a computer for further processing. Bipolar, rectangular surface electrode sensors (Bagnoli, Delsys Inc., Boston, USA) with an interelectrode distance of 1 cm were used. In accordance to the SENIAM recommendations [27] the EMG sensors were attached to the subjects skin. One sensor was attached to the clavicular muscle head of the pectoralis muscle (pars clavicularis, PCL), one to the sternal muscle head (pars sternocostalis, PST), one to the anterior deltoid (pars clavicularis, DCL) and one to the latissimus dorsi muscle (LTD) respectively. A 5
Data analysis
Surface EMG signals were demeaned and bandpass zero-lag filtered (10–450 Hz) with a digital fourth-order Butterworth filter for further analysis. The SLR was manually identified. The onset of the SLR was defined as the first consistent rapid increase in sEMG activity following the trigger stimulus. The second zero crossing of the signal determined the end of the SLR (Fig. 2). The peak-to-peak amplitude was calculated for the selected part of the signal. SLR values were normalized to the peak-to-peak amplitude of the MVC trial, which was selected out of a 1 s plateau. Trials with no obvious stretch reflex response were excluded from further analysis.
Descriptive statistics (mean
standard deviation) for muscle reflex latencies (in ms), muscle reflex amplitudes (in % of MVC) and muscle activation duration (in ms)
Descriptive statistics (mean
Schematic diagram showing the pectoral clavicular head reflexive response (sEMG, upper trace) to an external shoulder perturbation and the dynamometer angle (lower trace) during a 25% MVC 50% of iMER. The mechanical delay was determined via two accelerometers, muscle reflex latency was defined as time from actual perturbation to first muscle response.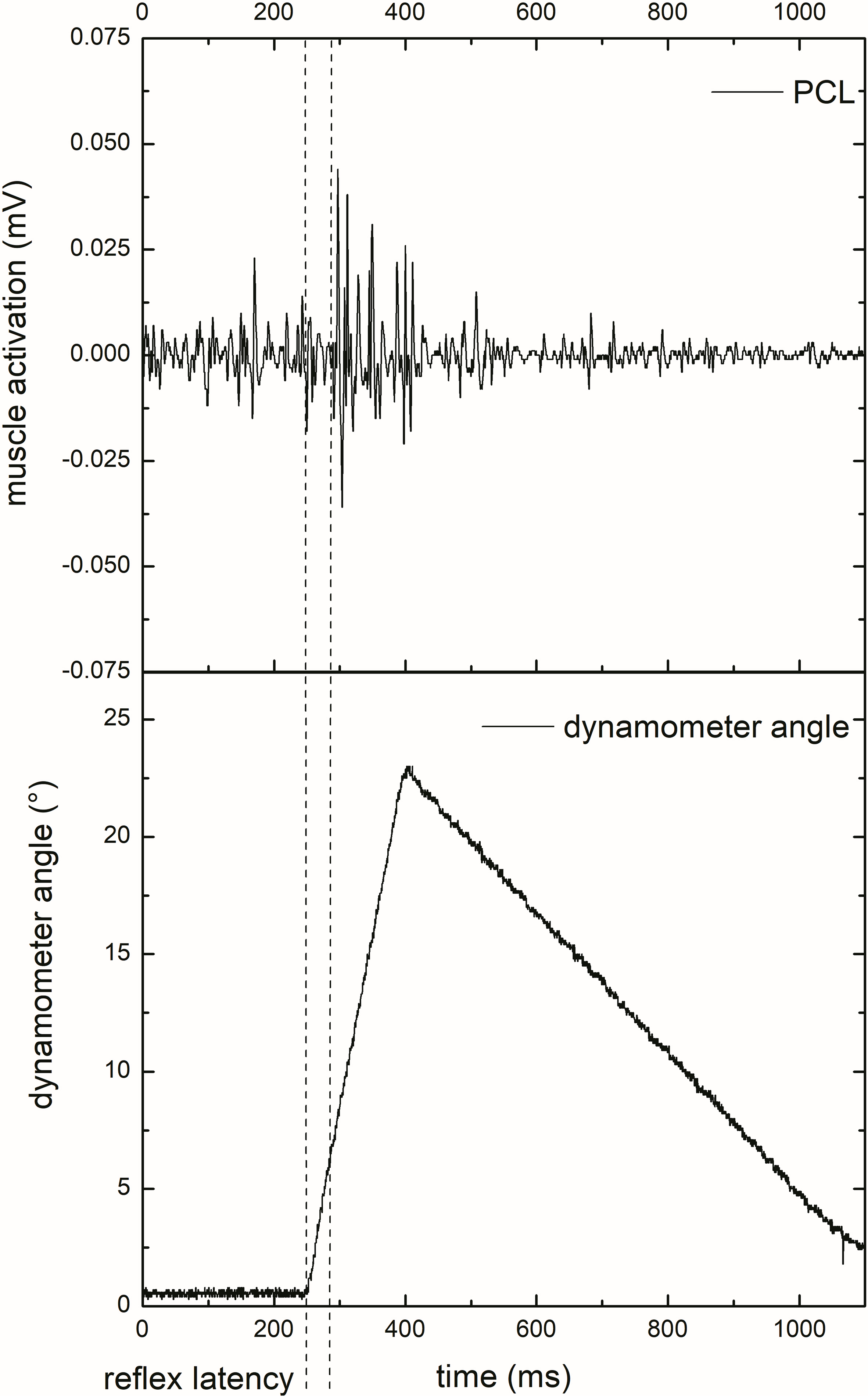
Top: Schematic diagram showing dynamometer angle and stretch velocity in 50% of iMER condition; bottom: Schematic diagram showing dynamometer angle and stretch velocity in 100% of iMER condition.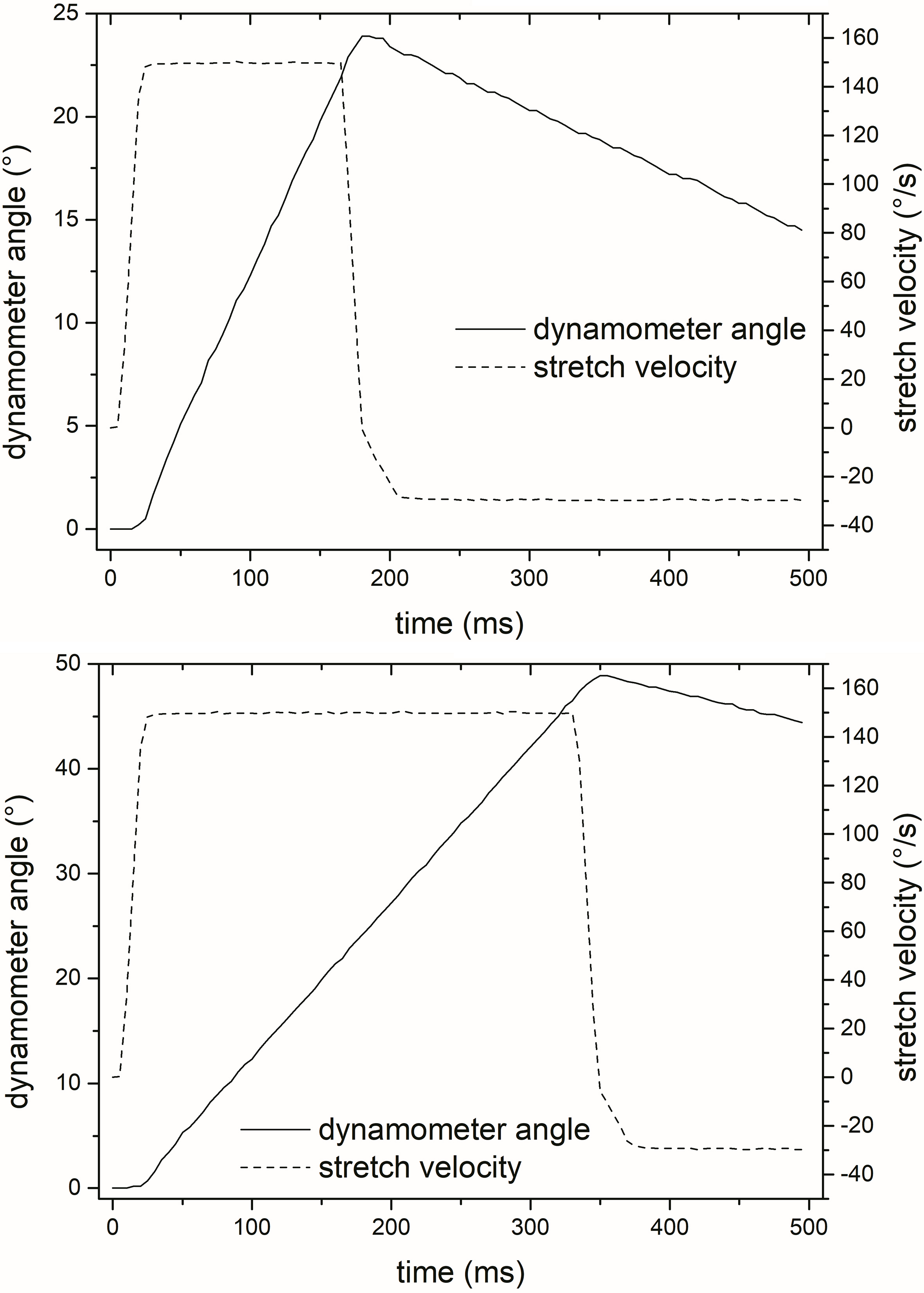
Shoulder muscle reflex latencies in all tested conditions and muscles. 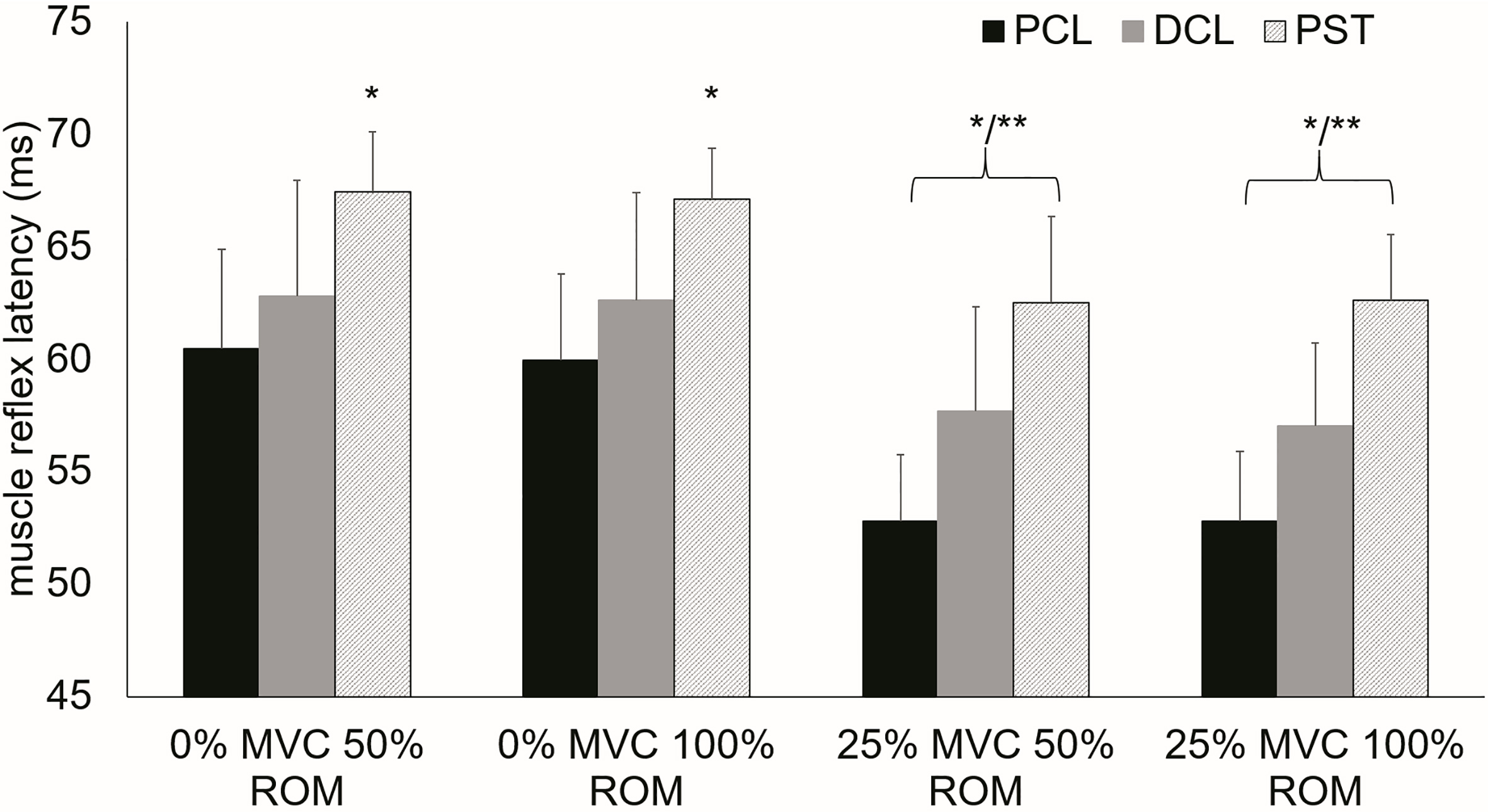
Shoulder muscle reflex amplitude in all tested conditions and muscles. 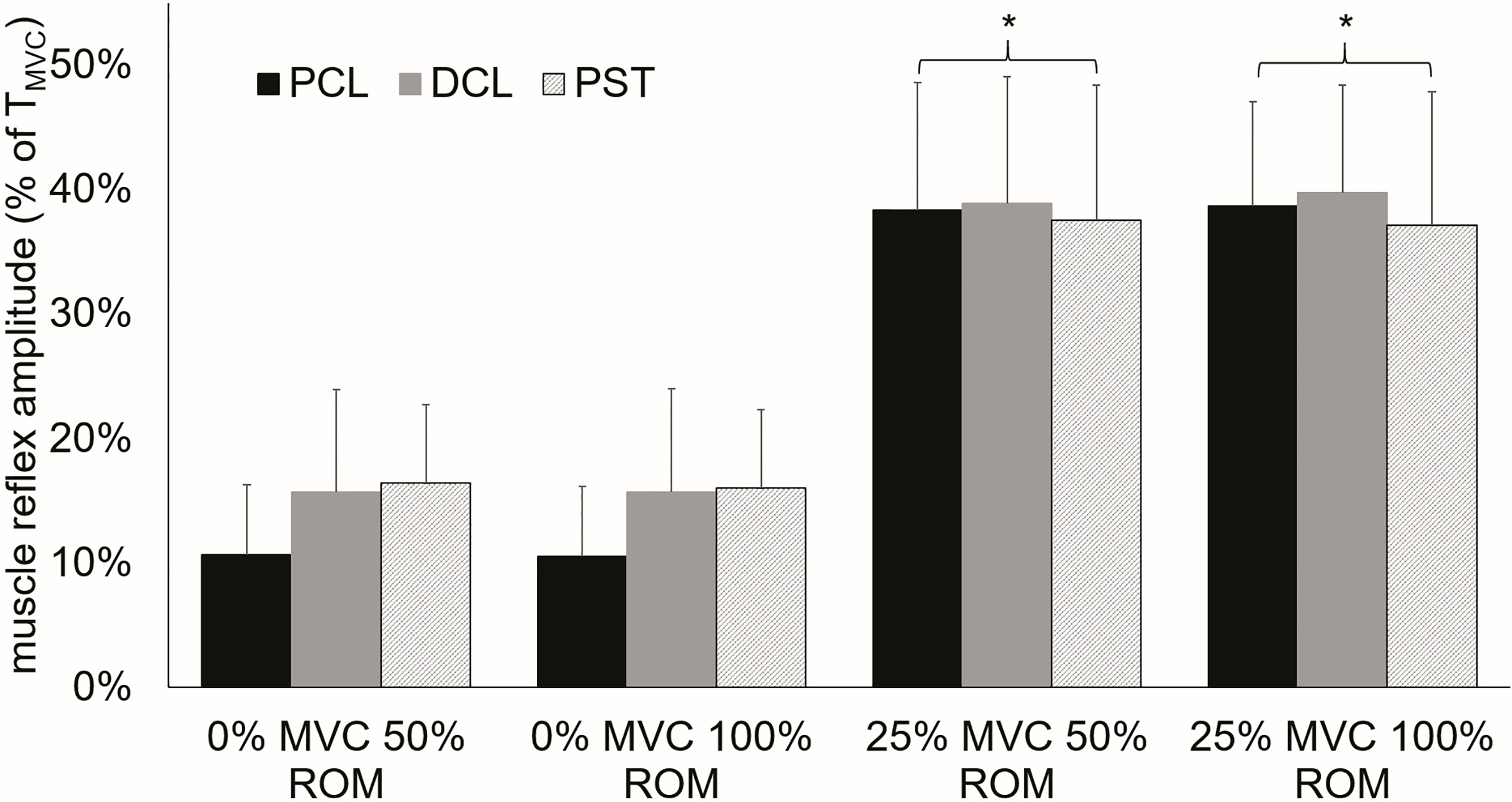
Shoulder muscle activation duration following stretch reflex onset. Significant increased activation duration between 50% of iMER and 100% of iMER in 0% MVC condition. Significant longer activation duration in trials with 25% MVC pre-activation compared to 0% MVC trials. No difference between different stretch amplitudes in 25% MVC condition.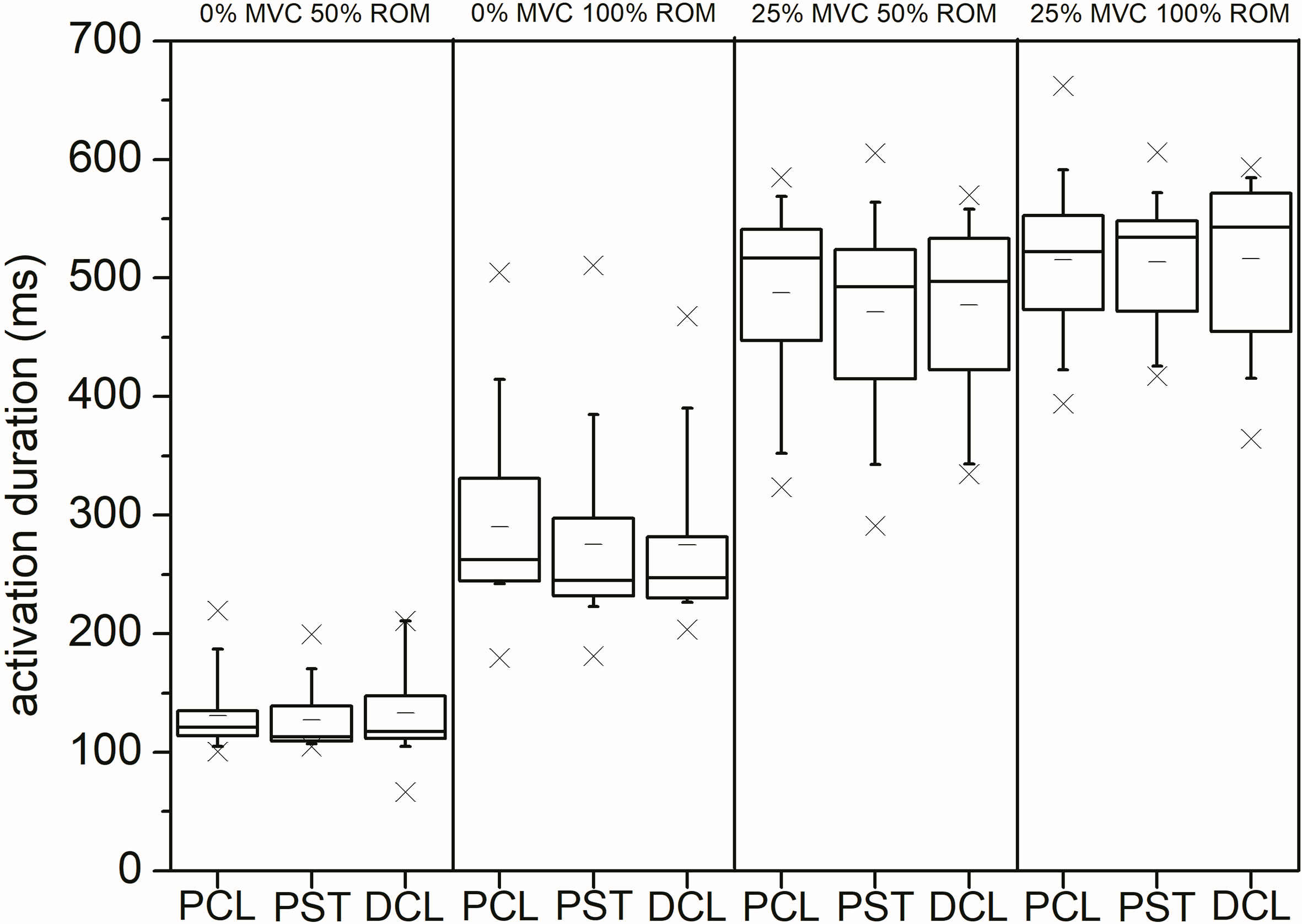
All statistical data analyses were carried out on SPSS 25 (IBM Corporation, Armonk, USA). Three within-factor repeated measures ANOVA with Bonferroni post-hoc tests were performed to determine main effects and factor interactions. The within-factors were the level of pre-activation moment (0% MVC, 25% MVC, 50% MVC), the applied stretch amplitude (50% of iMER, 100% of iMER) and the tested muscle (6 muscles). The significance level was set at
Results
In all trials the targeted dynamometer acceleration of 10
Discussion
The results from the current research indicate that muscle pre-activation has a significant effect on the short latency stretch reflex. Contraction levels of 25% of MVC led to a significant quickening of the reflex latencies due to the applied stretches. Significant faster muscle responses during pre-activated test conditions were observed in the clavicular and the sternal head of the pectoral muscle and the anterior head of the deltoid (Fig. 4). In addition pre-activation led to a significant increase of stretch reflex amplitude in all muscles and to an increase of muscle activation duration as an indication of increased excitation of the motoneurons [28]. Variation of the stretch amplitude did not show an effect on reflex latency or amplitude. However, an increase in stretch amplitude influenced muscle activation duration by increasing activation duration significantly. This observation blurred in the presence of muscle pre-activation. Overall the observed reflex latencies of specifically trained overhead athletes tended to be faster than formerly reported latencies [17, 18, 29] for the internal shoulder rotator muscles.
The stretch velocity as an important factor that influences the outcome size of the reflex, since the sEMG amplitude is highly related to the velocity of the stretch [cf. 30], was set to 150
It is well established that muscle pre-activation decreases the latency of the stretch reflex [cf. 30]. The decrease in reflex latency can be explained by an increase in muscle spindle sensitivity to intramuscular lengthening [15, 33] resulting in a prompt detection of the stretch caused by the anterior perturbation moment and therefore evoking a swifter muscular response. The significantly longer reflex latency of the sternal head of the pectoral muscle seems logical when taking pectoral muscle architecture into account. Despite its relatively small morphological part of total pectoral muscle (approximately 19% of total volume) [34] the clavicular head is the major generator of an internal rotation muscle moment at the shoulder. The pectoral tendon consists of two components one of which is only connected to the clavicular head. The sternal head of the muscle is substantially bigger with a volume of approximately 81% of total muscle volume [34]. Yet it is contracting synergistically during internal shoulder rotation. The normalized reflex amplitude elicited by the stretch allows a rough indication of the proportion of the motoneuronal pool that was excited by the stretch of the muscle. Significant increases in the observed reflex amplitude in 25% MVC pre-activated muscles can be explained by a full activation of the fusimotor system which has been suggested to be reached at the chosen pre-activation level [35]. Furthermore
In contrast to previous research, in our study a muscle pre-activation level of 50% MVC yielded no consistent stretch reflex in any investigated muscle [18]. We attributed the lack of consistent reflexive muscle responses to the considerably high background activation achieved during the pre-activation phase. Since electromyography measures the degree of muscle activation and the electrophysiological activation of a muscle initiates force generation strong evidence indicates that higher EMG levels correlate with higher muscle force [37]. During isometric contraction the biceps brachii muscle showed a non-linear relationship between measured force and muscle activation. There are also different EMG-force relationships due to differences in muscle architecture reported in the literature [cf. 37]. Furthermore Cronin et al. [38] revealed a negative correlation between higher forces and short-latency stretch reflex behaviour. They showed a decline in fascicle stretch velocity of over 50% between passive conditions and maximal force levels coinciding with progressively smaller muscular stretch response [cf. 38].
The latissimus dorsi muscle showed no response to the applied external shoulder rotation stretches. Given that the subjects were asked to produce an isometric internal rotation force to release the external shoulder perturbation trigger and taking previous findings into account [17, 18] we expected a reflexive response of the muscle. The observed muscle activation patterns during over-head throwing might contribute an explanation for the lack of reflexive muscle response. Jobe et al. [39] and Escamilla and Andrews [8] illustrated that the latissimus muscle is particularly activated during the late cocking phase and the early acceleration phase. In contrast to the starting position in our test setting the shoulder in that case was already located in a state of maximal external rotation which indicates that the reflex threshold for the latissimus dorsi was potentially not reached within our test setting. Furthermore, we cannot exclude a slight stretch evasion of the subjects by arching their backs in the late phase of the external shoulder perturbation which might have had an inhibiting influence on latissimus stretch response.
The relatively limited knowledge about reflex activity at the shoulder results in difficult comparisons among studies. To the best of our knowledge only four publications with comparable test settings to assess reflex latency at the shoulder exist [17, 18, 29, 40]. Wallace et al. [29] tested subjects with and without unstable shoulders. Although they used an accelerometer attached to the dynamometer to determine perturbation onset the results of this investigations are difficult to compare to our findings. The tested subjects showed significantly longer reflex latencies. No pre-activation condition was examined and moreover the authors did not reveal the acceleration which was used to apply the stretches. Latimer et al. [17] assessed muscular reflex response by a rope and pulley system using different weights to simulate an anterior translation force to the humerus. Subjects were placed in a supine position. Further methodological incomparability between this study and our research led to a difficult if not impossible comparability of the results. The results of Myers et al. [18] are similar to those of the current study. The tested muscles showed a significant pre-activation main effect showing the fastest reflexive response in the 20% pre-activation condition. The overall slightly longer latencies can be explained by the non-specifically trained participants examined since Brindle et al. [40] showed that a comparison between healthy subjects and trained intercollegiate pitchers express in faster reflex latencies for the pitchers. However, Brindle et al. [40] assessed reflex with an internal perturbation in contrast to the externally induced acceleration in our test setting.
Although this study suggests some interesting areas for further research, there were some limitations to the study design. One limiting factor was the fixation of the participants. As mentioned before the external shoulder rotations were applied in an upright sitting position by the use of a custom-made back pad to stabilize the non-dominant shoulder of the subjects. This set-up mimics the functional position of over-head throwing but still it cannot be ruled out that evasive trunk movements led to an increase of reflex latency. The increase might be explained by a variation of the limb position and as a consequence an inhibition of the length excitability of the motoneuron pool [cf. 30]. Furthermore, the chosen 90/90/90-starting position resulted in a slight pre-stretch of the internal shoulder rotator muscles of the shoulder joint. The increased muscle length and the pre-activation moment could reduce the impact of muscle slack which might limit the comparison of our outcomes to studies that chose different start settings. The potential influence of various starting positions must be investigated in further research. Finally the steering of the pre-activation moment level was a limiting factor of the study design. An extensive familiarisation was necessary to ensure reproducible pre-activation moment levels throughout the test session. Further research could facilitate this procedure by the use of a visual feedback system.
Conclusion
The results of the current research indicate that the chosen measurement setting was capable of eliciting muscular stretch reflexes in the internal shoulder rotator muscles. This observation further enhanced our knowledge of the stretch reflex induction and thus offers a more complex method to analyse neural muscle function for clinical applications such as stroke and spasticity treatment. Although the reflex latencies during pre-activation behaved as hypothesised the question still remains how these observations fit into the model of the ‘kinetic chain’ for high-speed over-head throwing. To get closer to a solution the influence of limb positioning, different stretch velocities and acceleration of the stretches needs to be further investigated to get a better understanding of the role of shoulder muscle stretch and energy storage at the shoulder during specific high-speed tasks such as over-head throwing.
Footnotes
Acknowledgments
The study was not funded and there is no conflict of interest. The authors thank Florian Weiser for assistance in customizing the MATLAB 2016a software for EMG-data processing.
Conflict of interest
None to report.