
Abstract
BACKGROUND:
Non-small cell lung cancer (NSCLC) is the most commonly diagnosed solid tumor. Natural killer (NK) cell-based immunotherapy is a promising anti-tumor strategy in various cancers including NSCLC.
OBJECTIVE:
We aimed to investigate the specific mechanisms that regulate the killing effect of NK cells to NSCLC cells.
METHODS:
Reverse transcription-quantitative PCR (RT-qPCR) assay was applied to measure the levels of hsa-microRNA (miR)-301a-3p and Runt-related transcription factor 3 (RUNX3). Enzyme-linked immunosorbent assay (ELISA) was used to measure the levels of IFN-
RESULTS:
A low expression of hsa-miR-301a-3p was observed in NK cells stimulated by IL-2. The levels of IFN-
CONCLUSIONS:
Hsa-miR-301a-3p suppressed the killing effect of NK cells on NSCLC cells by targeting RUNX3, which may provide promising strategies for NK cell-based antitumor therapies.
Introduction
Lung cancer (LC) is the leading cause of cancer-related death worldwide. Among them, non-small cell lung cancer (NSCLC) is the most common type of LC. Although effective advances in therapeutic outcomes have been achieved, the prognosis for patients with NSCLC is still poor [1]. The pathogenesis of NSCLC is extremely complex, and various signaling cascades including Akt and mTOR are also involved in regulating the progression of NSCLC [2]. Targeted therapy is widely used in LC with the advent of directed therapy toward certain driver genetic alterations [3]. Recently, clinical trials have also demonstrated the effectiveness of immunotherapy in NSCLC, and this evidence supports the critical role of enhancing host immunity in fighting NSCLC [4]. For example, the KEYNOTE-024 trial compared the 5-year outcome of pembrolizumab with chemotherapy as first-line treatment in advanced NSCLC patients with
As a type of innate immune cell, natural killer (NK) cells can swiftly kill multiple adjacent cells if these show surface markers associated with oncogenic transformation [8]. This property, which is unique among immune cells, and their capacity to enhance antibody and T cell responses support a role for NK cells as anticancer agents [9]. NK cells play critical roles in antitumor immunity in cancers, including pancreatic carcinoma and breast cancer [10, 11]. In the last years, NK cell-based immunotherapy has emerged as a promising therapeutic approach for solid tumors and hematological malignancies [9, 12]. NK cell-based immunotherapies mainly include combined cytokine, complement-dependent cytotoxicity (CDC) and antibody-dependent cell-mediated cytotoxicity (ADCC), NK-92, killer cell immunoglobulin-like receptor (KIR) mismatch and chimeric antigen receptor (CAR) approaches [13]. Among them, the concept of treating cancer with cytokine stimulated NK cells is long-established. NK cell activation involves a variety of signals such as interleukin-2 (IL-2). IL-2 activates NK cells to induce the secretion of cytokines and chemokines [14]. IL-2 stimulates the proliferative and cytotoxic activity of NK cells in pre-clinical models, NK cell-based therapies have been revealed to prevent the development of lung metastases [15]. Some researchers also found that patients who received an infusion of NK-92 cells could produce some encouraging responses including patients with advanced LC [16]. However, the therapeutic efficacy of NK cells is clinically limited due to the poor infiltration in NSCLC and the biological mechanism of NK cells in NSCLC remains controversial. Therefore, understanding of these mechanisms can contribute to approaches that enhance NK cell-mediated tumor cell clearance.
As small non-coding RNAs, microRNAs (miRNAs) act as oncogenes or tumor suppressors [17]. They are essential regulators of various cellular processes by targeting messenger RNA (mRNA) resulting in mRNA degradation and protein translation inhibition [18]. Aberrant expression of miRNA is associated with the progression of various cancers. For example, hsa-miR-301a-3p plays an oncogenic role in triple-negative breast cancer by regulating MEOX2 expression [19]. Further, the upregulation of hsa-miR-301a-3p can facilitate the growth and metastasis of colorectal cancer cells [20]. More importantly, hsa-miR-301a-3p can promote growth and aerobic glycolysis of NSCLC cells [21, 22]. Therefore, elucidating the mechanism of hsa-miR-301a-3p in NSCLC is urgently needed.
Runt-related transcription factor 3 (RUNX3), a DNA-binding transcription factor, is involved in many diseases and cell physiological processes [23]. Su et al. observed that RUNX3 was reduced in NSCLC tissues and cells [24]. Moreover, Fang et al. reported that RUNX3 could regulate the killing ability of NK cells [25]. Combining this evidence, we speculate that RUNX3 may affect the cytotoxicity of NK cells in NSCLC. Using bioinformatics, we found the targeted binding sites between hsa-miR-301a-3p and RUNX3. Therefore, hsa-miR-301a-3p may influence the cytotoxicity of NK cells in NSCLC cells by RUNX3.
This study found that hsa-miR-301a-3p was downregulated in activated NK cells and could inhibit the cytotoxicity of NK cells by RUNX3 in NSCLC. The findings provide promising targets for antitumor therapies based on NK cells.
Materials and methods
Cell treatment
NSCLC (A549 and H1299), 293T and NK (NK-92; ATCC Number: CRL-2407
Cell transfection
pcDNA-RUNX3 was synthesized in GenePharma (Shanghai, China) by inserting the full-length sequences of RUNX3 into pcDNA.3.1 vector. Hsa-miR-301a-3p mimic and NC mimic were purchased from GenePharma. In vitro experiment, cell transfection was conducted with Lipofectamine
RT-qPCR assay
The total RNA of the tissues and cells was extracted using TRIzol (cat. no. 15596026; Thermo Fisher Scientific) according to the manufacturer’s instructions [29]. In brief added 1 ml of TRIzol reagent per 100 mg of tissue to the sample. Added 1 ml of TRIzol reagent per 1
Western blot
Total proteins were extracted from tissues and cells using RIPA lysis buffer (Beyotime Institute of Biotechnology) containing protease inhibitor [34]. In brief added 1 ml of cold RIPA lysis buffer per 1
Enzymelinked immunosorbent assay (ELISA)
After centrifugation, the supernatants of the NK-92 cells were obtained for IFN-
Cytotoxicity assay
For co-cultured experiments of NK cells and cancer cells, some methods in other reports were referred to by us. Wei et al transfected NK cells were co-incubated with MGC-803 at effector cell/target cell (E:T) ratios of 5:1 for 6 h [38]. The report of Fang and his partners demonstrated that IL-2 activated NK92 cells with different transfection were co-cultured with cancer cells for 4 h at 37
Dualluciferase reporter assay
The target reaction between hsa-miR-301a-3p and RUNX3 was predicted using TargetScan (
RNA immunoprecipitation (RIP)
The lysate of the transfected NK-92 cells was cultured with protein A/G magnetic beads and Argonaute2 antibody (Ago2; Abcam). IgG (Abcam) was used as a control. Then, RNA in the immunoprecipitated complex was purified with RNase-free Dnase I (Thermo Fisher Scientific) and proteinase K (Solarbio, Beijing, China). The mRNA levels of RUNX3 in different group cells were measured with RT-qPCR. The expression of RUNX3 in Ago and IgG group was compared with the input group.
Xenograft model of NSCLC
The experiment was approved by the Institutional Animal Care and Use Committee of Zhongshan City People’s Hospital (Approval No. ZSCPH-IACUC-2022-B0801). For the model construction, A549 cells (6
Statistical analysis
All experiments, except for in vivo procedures, were performed in triplicate. Data were processed using the GraphPad Prism software (version 8.0) and presented as mean
Results
Hsa-miR-301a-3p signal was decreased in NK-92 cells activated by IL-2
To study the effect of hsa-miR-301a-3p in NK-92 cells on NSCLC cells, we activated NK-92 cells with IL-2. Significantly elevated levels of IFN-
Decreased hsa-miR-301a-3p expression in NK-92 cells activated by IL-2. NK-92 cells were treated with IL-2 (20 ng/ml) for 24 h. (A) The secretions of IFN-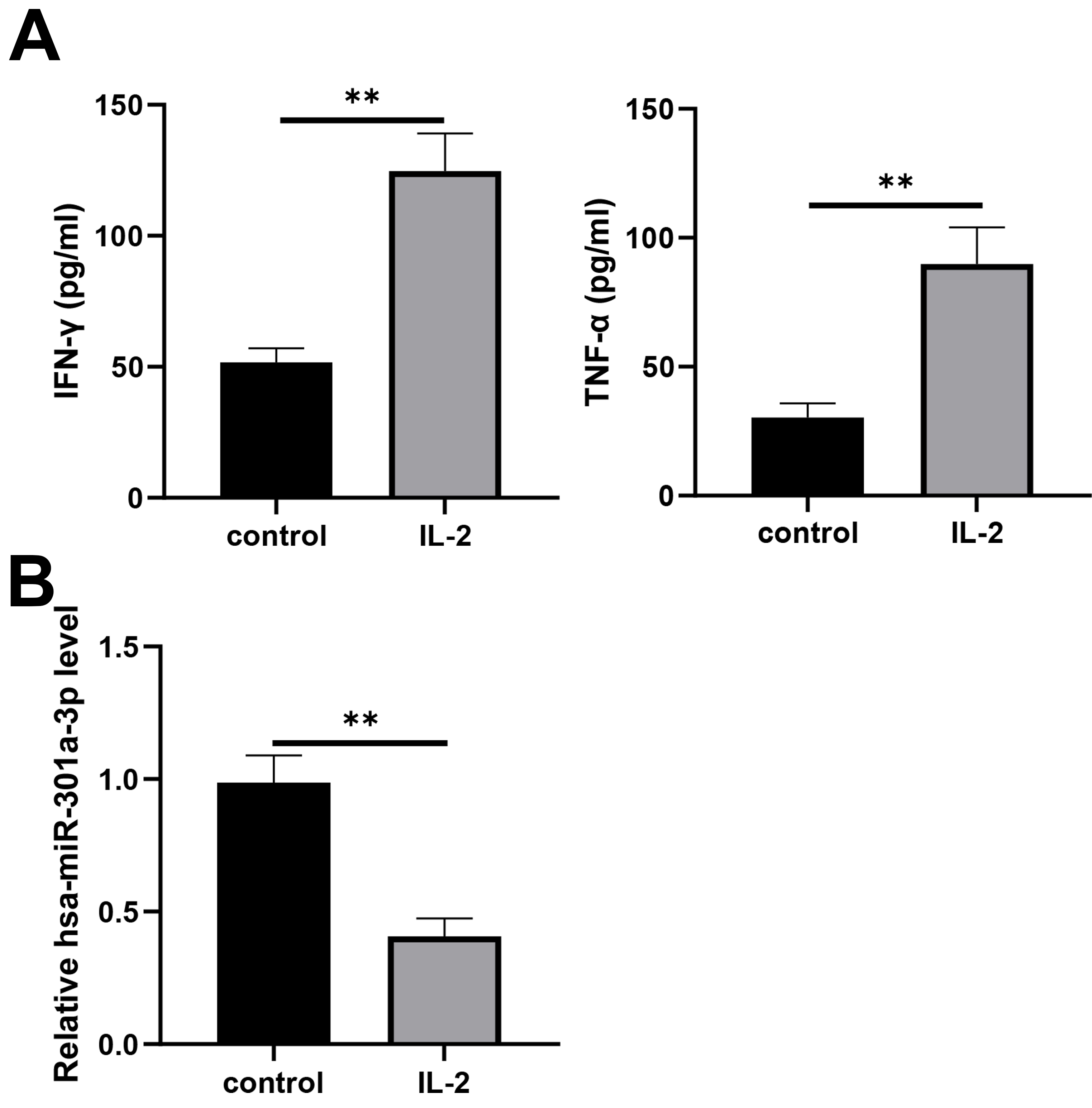
Due to the downregulation of hsa-miR-301a-3p on NK-92 cells activated by IL-2, we speculated that hsa-miR-301a-3p played a role in the cytotoxicity of NK-92 cells against NSCLC cells. After the activation with IL-2, NK-92 cells were treated with hsa-miR-301a-3p mimic for the increase in hsa-miR-301a-3p expression (Fig. 2A) and a decrease in IFN-
Hsa-miR-301a-3p inhibited the cytotoxicity of NK-92 cells against NSCLC cells. NK-92 cells were treated with IL-2 (20 ng/ml), followed by transfection of a hsa-miR-301a-3p mimic. (A) Hsa-miR-301a-3p expression was investigated with RT-qPCR. (B) The secretions of IFN-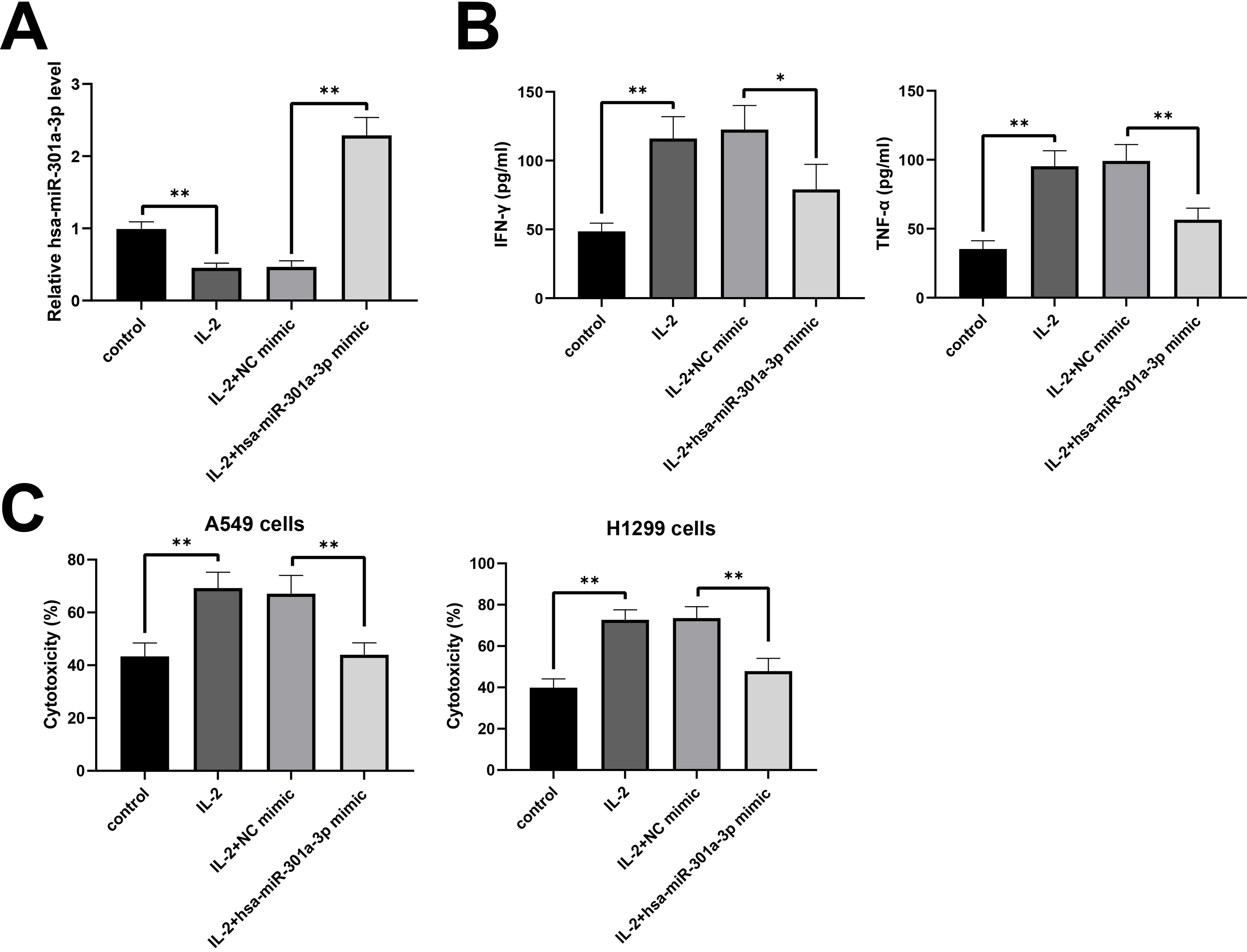
To further explore the specific molecular mechanism, the miRNA target prediction of hsa-miR-301a-3p was evaluated using TargetScan. There were highly conserved combination sites between the 3
RUNX3 as a direct target gene of hsa-miR-301a-3p. (A) TargetScan was used to predict the binding between hsa-miR-301a-3p and RUNX3. (B) The verification of the interaction between hsa-miR-301a-3p and RUNX3 by dual-luciferase reporter assay. (C) RUNX3 protein expression in NK cells transfected with hsa-miR-301a-3p mimic and relevant control was detected by western blot. (D) The enrichment of RUNX3 in NK-92 cells with the hsa-miR-301a-3p mimic was detected with the RNA immunoprecipitation (RIP) assay. 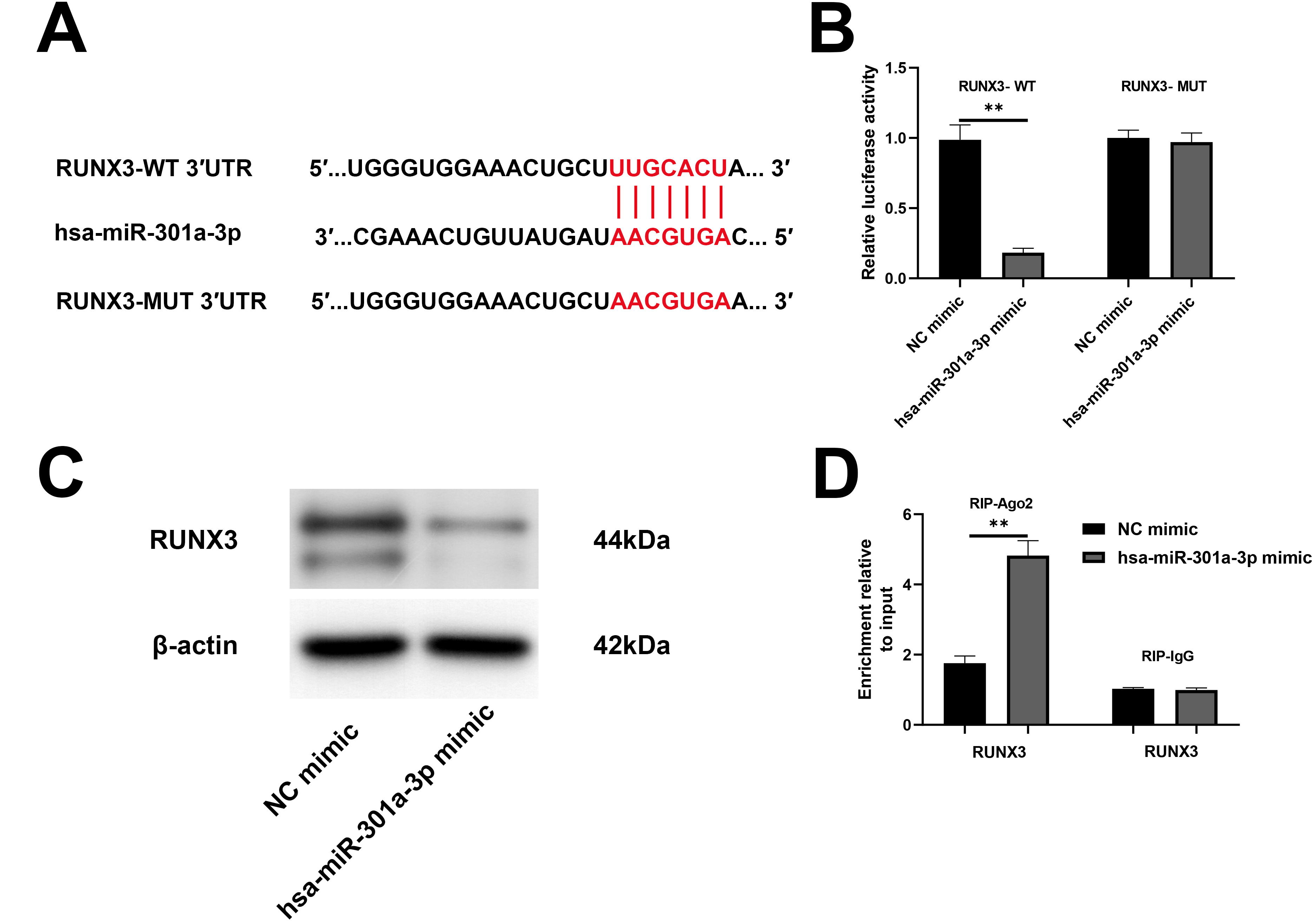
To demonstrate whether hsa-miR-301a-3p suppressed the cytotoxicity of NK-92 cells by RUNX3, activated NK-92 cells were treated with hsa-miR-301a-3p mimic and/or pcDNA3.1-RUNX3. As illustrated in Fig. 4A, hsa-miR-301a-3p was upregulated in the hsa-miR-301a-3p mimic group and not affected by pcDNA3.1-RUNX3. RUNX3 was downregulated in the hsa-miR-301a-3p mimic group, and pcDNA3.1-RUNX3 reversed the inhibitory of the hsa-miR-301a-3p mimic (Fig. 4B). IFN-
Hsa-miR-301a-3p inhibited the cytotoxicity of NK-92 cells through RUNX3. NK-92 cells were treated with IL-2 (20 ng/ml) and hsa-miR-301a-3p mimic and/or pcDNA3.1-RUNX3. (A) Hsa-miR-301a-3p expression was investigated using RT-qPCR. (B) RUNX3 expression was detected by western blotting. (C) The secretions of IFN-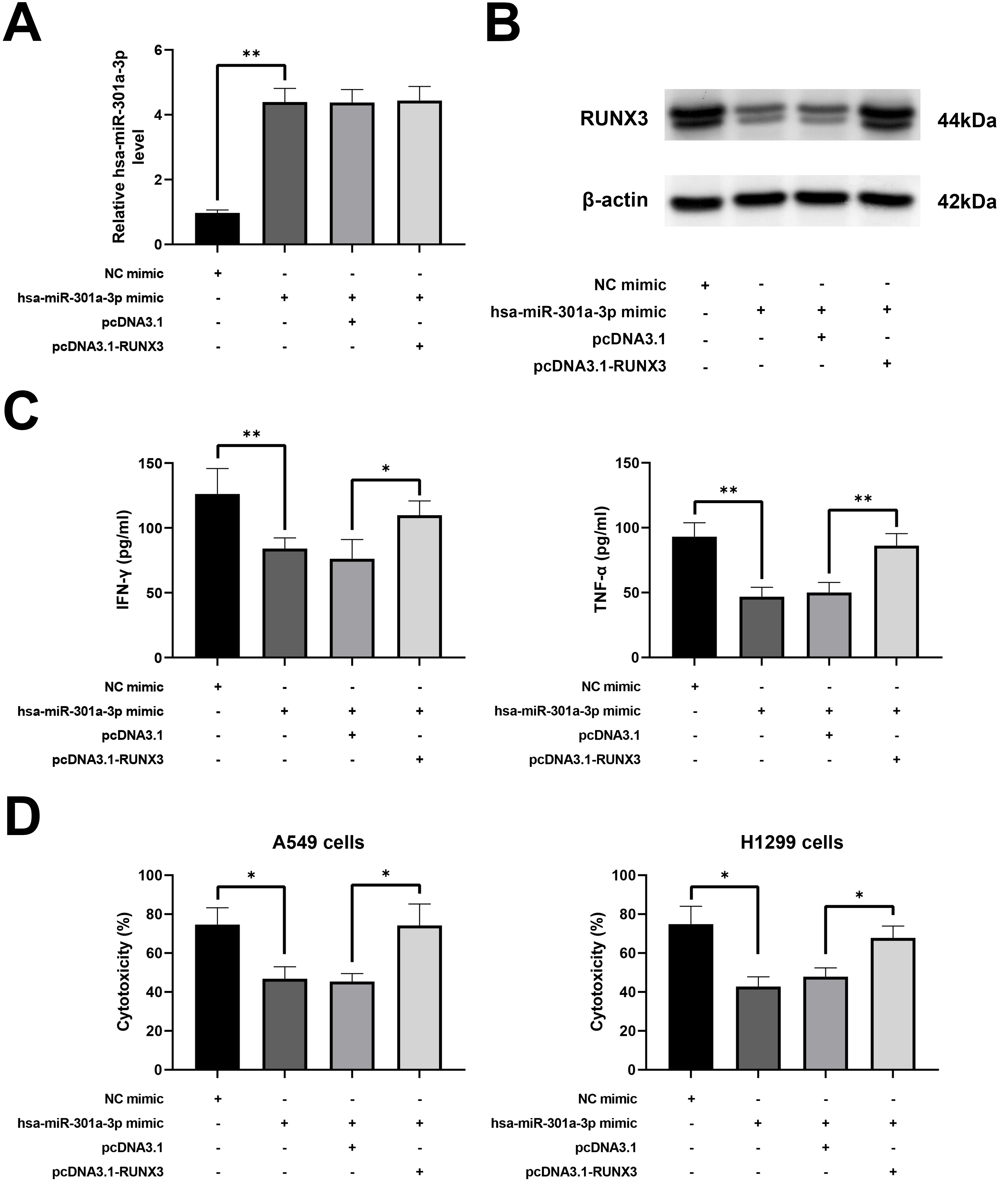
The effect of hsa-miR-301a-3p was verified in vivo Activated NK-92 cells treated with hsa-miR-301a-3p were introduced into the BALB/c nude with A549 cell subcutaneous xenograft tumors. Treatment with the lenti hsa-miR-301a-3p mimic increased the tumor volume (Fig. 5A). The tumors in mice treated with the lenti hsa-miR-301a-3p mimic were larger and heavier than those treated with the NC mimic (Fig. 5B and C). Moreover, hsa-miR-301a-3p expression was increased and RUNX3 expression was decreased in lenti hsa-miR-301a-3p mimic group (Fig. 5D and E). Collectively, these results provided evidence that hsa-miR-301a-3p promoted tumor growth by suppressing the killing effect of NK cells against NSCLC cells.
Hsa-miR-301a-3p inhibited NK-92 cell-mediated killing of NSCLC cells in vivo. (A-C) Tumor volume was examined every 3 days with slide calipers (0.5 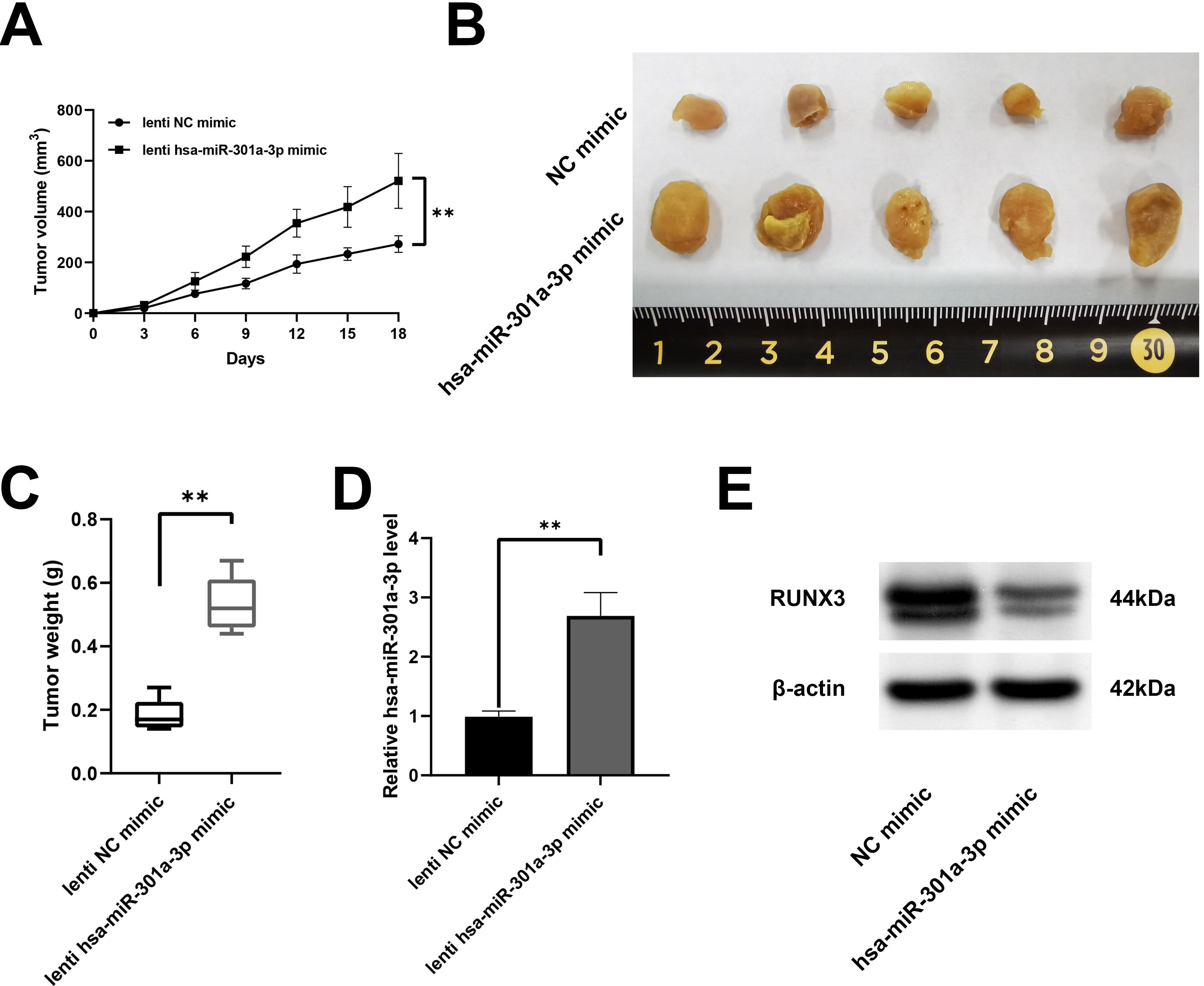
In our study, hsa-miR-301a-3p was remarkably decreased, and IFN-
As one of the vital immune cells, NK cells are crucial for eliminating tumor cells [41]. Although its cytolytic potential for NSCLC is limited, enhancing the function of NK cells may enhance its tumor regression effect [42]. Moreover, increased NK cell infiltration suppressed the development of NSCLC [43]. Activated NK cells induced cytolysis of tumor cells by regulating the levels of proinflammatory cytokines [44]. Therefore, the activated NK cells observed in this study could inhibit the development of NSCLC.
RUNX3 is a member of the RUNX family, which plays a significant role in cell growth and differentiation [45]. Abnormal expression of RUNX3 has been found in malignant tumors, and it played an inhibitory part in the carcinogenesis of different cancer types through various signaling pathways [46]. RUNX3 is associated with the progression of NSCLC [24, 47, 48]. In addition, RUNX3 expression was found in NK cells, while long non-coding RNA (LncRNA) GAS5/ hsa-miR-544/RUNX3 enhanced the elimination of NK cells in liver cancer [25]. This study confirmed that RUNX3 could enhance the cytotoxicity of activated NK cells on NSCLC cells.
As a member of the hsa-miR-301 family, hsa-miR-301a-3p is related to various tumors [49, 50, 51]. The expression of hsa-miR-301a-3p was abnormally high in NSCLC tissues [21, 22]. In addition, some researchers have found that RUNX3 was targeted by hsa-miR-301a-3p in some types of cancers [20]RUNX3 was verified to be a direct target gene of hsa-miR-301a-3p in colorectal cancer cell [20]. In prostate cancer, some researchers revealed that RUNX3 was a target of hsa-miR-301a-3p, which was confirmed by dual-luciferase reporter assay [52]. Combined with the above reports and the results of our study, we assumed that hsa-miR-301a-3p regulated RUNX3 to suppress the cytotoxicity of NK cells on NSCLC and promoted the development of NSCLC. At the same time, Li and his partners found that down-regulated expression of hsa-miR-301a enhanced infiltration of CD8
However, there are some limitations in this research. First, the regulation of hsa-miR-301a-3p/RUNX3 axis on the tumor microenvironment of NSCLC is multifaceted, while this study only proved its regulatory effect on NK cells. The mouse model in the present study also could not reflect the real tumor microenvironment. A stable mouse model with immune reconstitution needs to be established in the future. In addition, future studies can focus on the role of hsa-miR-301a-3p/RUNX3 in various immune cells to find new therapeutic strategies for NSCLC.
Conclusion
In summary, hsa-miR-301a-3p was underexpressed in activated NK cells and restrained RUNX3. Moreover, the elimination of NK cells to NSCLC was suppressed by hsa-miR-301a-3p/RUNX3. Hsa-miR-301a-3p may be as a potential immune therapeutic target of LC.
Author contributions
Conception: Zhihua Ye. Interpretation or analysis of data: Junkai Zhang and Zhihua Ye Preparation of the manuscript: Yingyu Yang, Ying Wei, Lamei Li and Xinyi Wang. Revision for important intellectual content: Junkai Zhang and Zhihua Ye. Supervision: Zhihua Ye.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Ethics approval and consent to participate
The experiments were approved by the Institutional Animal Care and Use Committee of Zhongshan City People’s Hospital (Approval No. ZSCPH-IACUC-2022-B0801).
Consent for publication
Not applicable.
Competing interests
The authors declare no conflict of interest.
Footnotes
Acknowledgments
The present study was supported by a grant (no. 81903029) from the National Natural Science Foundation of China Youth Project.