MicroRNAs (miRNAs) play potential role in the development of various types of cancer conditions including pancreatic cancer (PC) targeting several cellular processes. Present study was aimed to evaluate function of miR-125b and the mechanism involved in PC.
METHODS:
Cell migration, MTT and BrdU study was done to establish the migration capability, cell viability and cell proliferation respectively. Binding sites for miR-125b were recognized by luciferase assay, expression of protein by western blot and immunofluorescence assay. In vivo study was done by BALB/c nude xenograft mice for evaluating the function of miR-125b.
RESULTS:
The study showed that expression of miR-125b was elevated in PC cells and tissues, and was correlated to proliferation and migration of cells. Also, over-expression of miR-125b encouraged migration, metastasis and proliferation of BxPC-3 cells, the suppression reversed it. We also noticed that thioredoxin-interacting protein (TXNIP) was the potential target of miR-125b. The outcomes also suggested that miR-125b governed the expression of TXNIP inversely via directly attaching to the 3-UTR activating hypoxia-inducible factor 1 (HIF1). Looking into the relation between HIF1 and TXNIP, we discovered that TXNIP caused the degradation and export of HIF1 by making a complex with it.
CONCLUSION:
The miR-125b-TXNIP-HIF1 pathway may serve useful strategy for diagnosing and treating PC.
Pancreatic ductal adeno-carcinoma (PDAC) is reported to be one of the main factor responsible for cancer-specific deaths due to scarcity in available treatment options [1, 2]. The available treatment options have improved the overall survival rate in PDAC, however in many cases the clinical outcomes still remain poor [2]. The present day treatment options are insufficient for improving the clinical results for PDAC [3, 4]. Delayed diagnosis of PDAC is a challenge due its character of showing distant metastasis in body organs like liver, lungs, stomach, intestine and spleen [5]. In some cases the diagnosis in final stage is accompanied with surgical excision making chances of survival diminished. However, to our knowledge approaches which would decrease the relapse and metastasis of PDAC are lacking, hence studies confirming the potential genes linked to metastasis and progression of PDAC are of greatest importance for successful treatment of PDAC.
MicroRNAs (miRs) are small non-coding, endogenous RNA molecules made up of at least 18–24 nucleotides, they are responsible for degradation or inhibition of translation via binding to the complementary sequences in the 3UTR region of target mRNAs [6, 7]. Numbers of studies have confirmed involvement of miRs in cellular processes of cancer [8, 9, 10, 11]. MiRs have been associated with cell death, proliferation, differentiation, cell death, migration, angiogenesis, invasion and metabolism in PDAC [8, 12, 13]. MiR-125b have recently been found to be over-expressed in glioma, it is reported to function as an oncogene responsible for pathogenesis of glioblastoma cells, inhibition of miR-125b leads to proliferation of cancer cells [14]. In a report earlier, miR-125b has been found to regulate the process of proliferation and differentiation and migration of neuronal stem cells via targeting Nestin [15]. In another report, miR-125b was reported to promote cell proliferation via directly targeting Lin28 in glioma cells, the cells showed low expression of miR-125b [16]. These reports hint that miR- 125b behaves as oncogene, however, the role of miR-125b in pancreatic cancer remains unexplored. In the present study, we directed to explore the role and mechanism of miR-125b in PDAC.
Materials and methods
Cell lines and tissue samples
For the study we selected CFPAC1, SW1990, Panac-1, Panac-2, BxPC-3 PDAC cell lines (American type culture collection (USA). hTERT-HPNE and H6c7 (Kerafast (Boston, MA, USA)) were used as normal pancreatic cells. All the cells were grown at 37C under CO (5%) in Dulbecco’s modified Eagle’s medium (DMEM) (Thermofisher, USA) with 99% realative humidity, without antibiotics. Tissue samples were obtained from patients undergoing surgery, all the subjects were educated about the study and informed consent was obtained and signed from them. About 43 pairs of human PDAC tissues were collected along with the adjacent non cancerous tissues form Xinxiang Central Hospital, Xinxiang, Henan, China. The experiments of the study were approved by the ethical committee of Xinxiang Central Hospital, Xinxiang, Henan, China the approval number was XCHHEC2019-22. One part of tissue was frozen (80C) for extraction of RNA while the other part of tissue was embedded in paraffin for obtaining tissue sections.
Cell transfection
In the process of transfection of PDAC cells the miR-125b mimic, miR-125b inhibitor, siRNAs and pcDNA3.1-TXNIP targeted for HIF1- and TXNIP were all obtained from Genescript (USA). The plasmid pGL4.20-HIF1 for HIF1 was obtained from Addgene (USA). Site directed mutagenesis was utilized for generating mutation in TXNIP-CRM1. The primer sequences for miR-125b were F: 5-CGTCCCTGAGACCCTAACTT-3 and R: 5-GCACTGGATACGACTCACAA-3.
Transduction of PDAC cells for stable miR-125b and TXNIP over-expressing cell lines
The miR-125b inhibitor or miR-125b over-expressing cell lines were established by transducting the BxPC-3 and SW1990 with lentvirus expressing miR125b or miR-125b anti-oligonucleotides. The BxPC3 cells received transfection of pLV3-224-GFP which were called as BxPC-3-miR-125b cells, whereas, the SW1990 cells were transfected with pLV3-224i-GFP and were called to be SW1990-miR-125bi. Quantitative real time PCR (qRT-PCR) was used to detect the expression of miR-125b or miR-125b inhibitor. The qRT-PCR reaction was done in a total volume comprising 25 l. The cycling conditions were as follows, 95C for time period of 5 minutes, 40 cycles; at 95C for time period of 20 seconds; 60C for time period of 20 seconds and at 72C for time period of 20 seconds. In the process to develop TXNIP stable over-expressing cell lines, the BxPC3 or BxPC3 miR-125b over-expressing cells received transfection of pcDNA3.1-TXNIP followed by exposure to Neomycin, after which the, obtained stable over-expressing BxPC3 cells were labeled as BxPC3-TXNIP and BxPC3-miR-125b-TXNIP cells.
Immunoblotting studies and immunofluorescence
Immunoblot analysis in cells was done by subjecting the cells to lysis using a lysis buffer, the total protein was estimated using protein estimation kit (Sigma Aldrich, USA). The proteins were isolated by electrophoresis on sodium dodecyl sulfate-polyacrylamide gel (10%). The antibodies used in the study are described in Table 1. For the immunofluorescence study the cells were fixed using p-formaldehyde and incubated for 12 hours using I antibodies.
Antibodies used for the study
Name of antibodies
Suppliers
Vimentin
Cell signaling
TXNIP
MBL
E-cadherin
Cell signaling
HIF1
ProteinTech
Ki-67
Abcam
Actin
Cell signaling
BrdU assay
The BrdU measurement was done for assessing cell proliferation as per the supplier’s instructions. The pancreatic cancer cells were added with 10 l/well of BrDU (10 M) and incubated for 5 hours at 37C. The cells were fixed and incubated for 30 min at 37C, the fixing solution was removed by tapping. Each well received 100 l/well of working solution of BrdU antibody conjugated with peroxidase followed by incubation of 1.5 hours at 37C. The wells were washed using washing solution followed by addition of substrate solution (100 l). The absorbance was recorded at 454 nm.
The MTT assay
MTT assay was done for cell viability studies. After the PDAC cells received treatment, 10 mL of MTT (5 mg/mL) was added and kept for 4 hours. The culture medium was removed and 100 mL dimethyl sulfoxide was added to cells followed by incubation of 2 hours. The absorbance maxima was recorded at 490 nm.
Transwell cell migration assay
Transwell assay was done for studying cell migration. About 2 10 cells were transfected followed by trypsinization and seeded in the topmost chamber of Transwell cell. The cells were incubated at room temperature for 24 hours, the cells which migrated to the other compartment were fixed using methanol followed by staining with 5% crystal violet and counted selecting 5 independent fields.
The miR-125b target prediction study
The prediction study for evaluating the favorable target of 125b was done opting in silico tools such as Targetscan predictions for human, mouse and zebrafish. The total context score and the PCT i.e Probability of Conserved Targeting for prediction scores were selected.
Luciferase assay
The BxPC-3 cells expressing miR-125b were transfected using TXNIP 3-UTR luciferase reporter. The SW1990 cells received transfection of miR-125 binhibitor along with TXNIP 3-UTR luciferase reporter. The Luciferase activity was measured using dual Luciferase (Renilla and firefly) reporter assay system (Promega).
Pancreatic cancer metastatic mouse model (In vivo studies)
For in vivo studies BALB/c nude mice were used. Mice aging 4–6 weeks, weighing 17–20 g were selected and segregated into 4 groups with each comprising 5 mice. The animals were housed in polypropylene cages maintained at 24C with dark–light cycle of 12 h. The mice received food and water Ad libitum. All the animal protocols were in accordance to the draft of Animal protection law of the People’s Republic of China-2009 for experimental animals. The animal studies were sanctioned by the institutional animal ethical committee of ‘Xinxiang Central Hospital, Xinxiang, Henan, China’. The approval number was XCHAEC2019-02, the in vivo studies adhered to ARRIVE guidelines. About 1 10 cells of BxPC-3-miR-125b, BxPC-3-miR-NC, BxPC-3-TXNIP, BxPC-3-miR-125b-TXNIP, SW1990-miR-125bi, or SW1990-miR-NC were injected via tail vein for establishing pancreatic cancer mouse model. The mice were sacrificed after 10 weeks mainly for observing the presence of metastatic foci in lungs. Hematoxylin and Eosin (H&E) staining of lung tissues was done.
Statistical analysis
All the results are presented as mean standard deviation ( 3). Statistical analysis was done using Minitab18 (Minitab, USA) for performing Student’s -test (two-tailed) with Pearson’s correlation coefficient. Value of 0.05 was regarded significant.
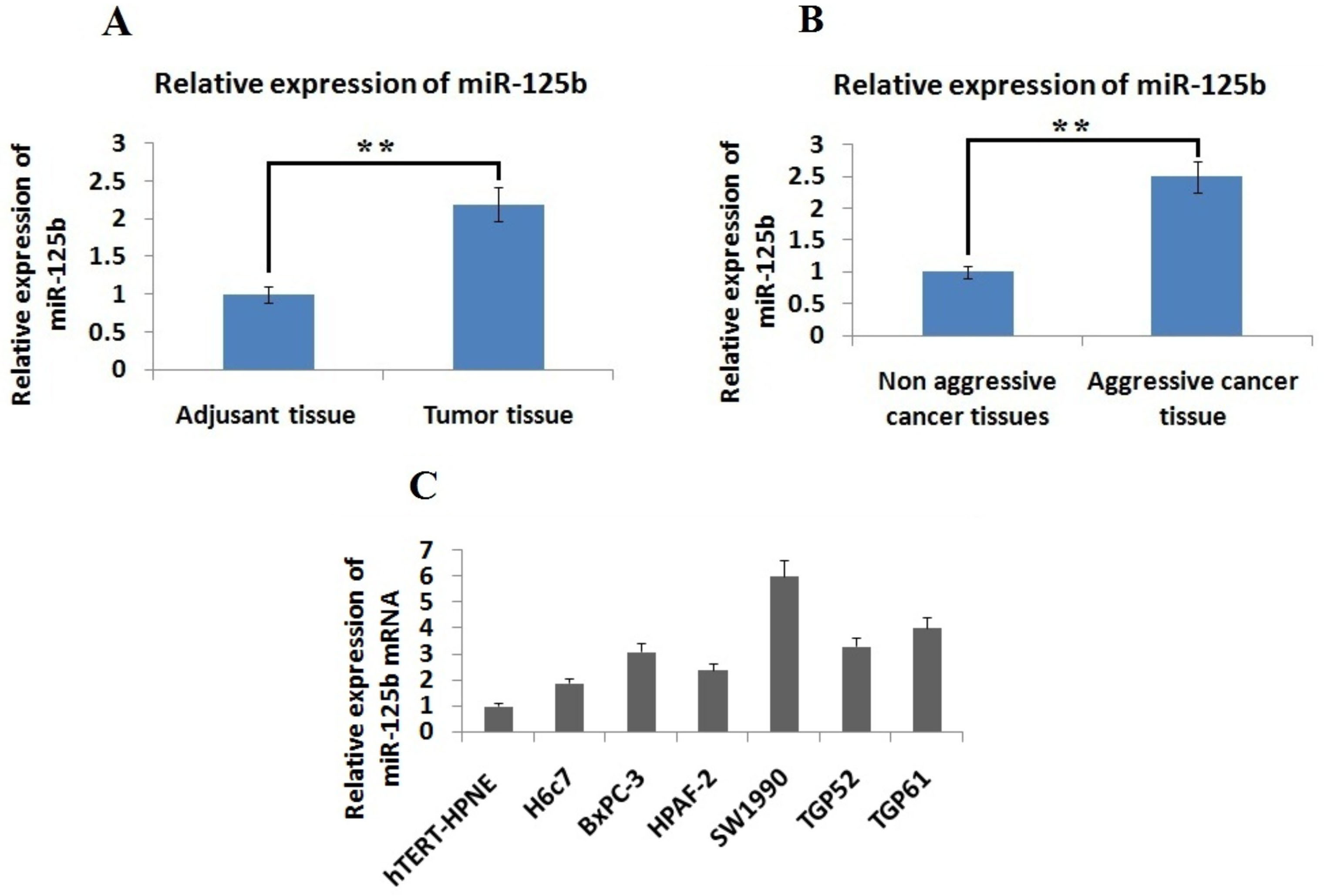
Expression of miR-125b was elevated in pancreatic cancer. Data showing comparison for expression of miR-125b in, A: In Pancreatic cancer tissues and adjacent non cancer tissues. ** 0.01 compared to adjacent tissues B: Non-aggressive and aggressive cancer tissues and C: Pancreatic cancer cell lines and normal pancreatic cells. ** 0.01 compared to non-aggressive cancer tissues.
Results
Expression of miR-125b is elevated in PDAC tissues and cells
To explore the involvement of miR-125b in progression of pancreatic cancer, we studied the levels of miR-125b via qRT-PCR. The levels of miR-125b were significantly elevated in PDAC tissues compared to the adjacent tissues ( 0.01) (Fig. 1A). The results suggested that, the levels of miR-125b were over-expressed in aggressive pancreatic cancer tissues compared to non aggressive tissues ( 0.01, Fig. 1B). We also studied the levels of miR-125b in different pancreatic cancer cells in comparison to normal epithelial cells i.e. hTERT-HPNE and H6c7. We found that levels of miR-125b were over-expressed significantly in the entire selected PC cell lines compared to normal epithelial cells (Fig. 1C). Altogether, the results suggested that miR-125b was over-expressed in PC cells and cancerous tissues compared to normal cells and adjacent non cancer tissues respectively. The experiment hence confirms up-regulation of miR-125b as a distinctive feature in development and progression of PDAC.
MiR-125b encourages proliferation and migration of pancreatic cancer cells
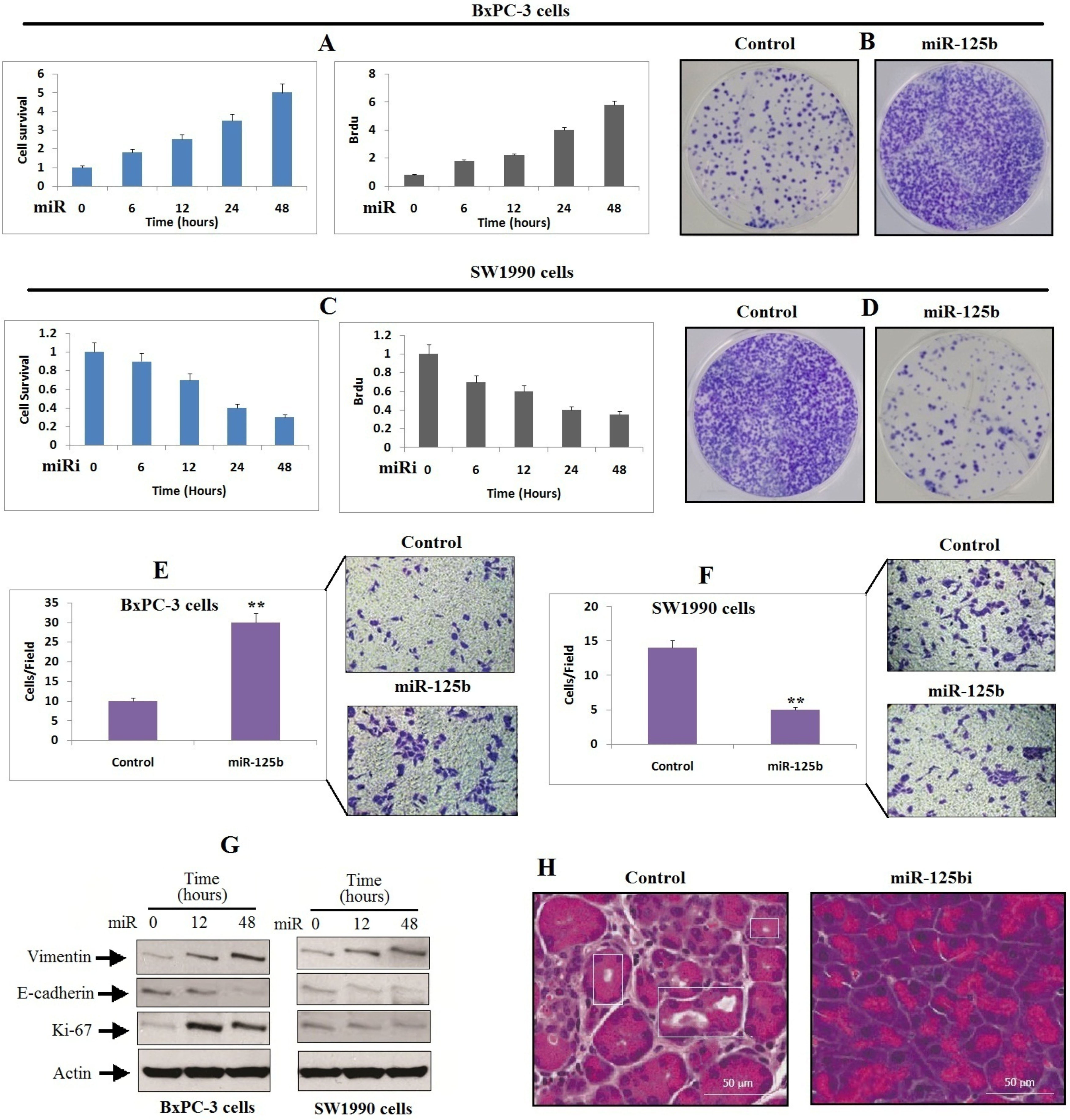
To further explore the involvement of miR-125b in development and progression of pancreatic cancer, BxPC-3 and SW1990 PDAC cell lines were chosen for the study as they exhibited variable expression of miR-125b (Fig. 1C). Both the PDAC cancer cell lines BxPC-3 and SW1990 were transfected with miR-125b mimic (miR) or inhibitor (miRi), the cell lines were assessed for proliferation and cell growth via BrDu and MTT assay. We found that, transfection of miR-125b in BxPC-3 caused a significant elevation in levels of miR-125b and caused a subsequent increase in cell proliferation with increasing time (Fig. 2A and B). In contrast, suppression of miR-125b by miR-125bi inhibited the levels of miR-125b, and halted the proliferation of BxPC-3 cells (Fig. 2C and D). A study earlier have evidenced that epithelial mesenchymal transition (EMT) is a potential regulator of metastasis [17], we hence evaluated whether miR-125b contributes in metastasis of pancreatic cancer by regulating EMT. We noticed that upregulation of miR-125b encouraged the migration of BxPC-3 cells (Figu. 2E), whereas inhibition of miR-125b via miRi halted the migration of SW1990 cells (Fig. 2F). In addition to this, the outcomes of Immunoblotting study revealed that upregulation of miR-125b suppressed the levels of E-cadherin (epithelial marker) but elevated the levels of Ki-67 (proliferation marker) and vimentin (mesenchymal marker) in BxPC-3 cells (Fig. 2G). In contrast, the decreased expression of miR-125b in SW1990 cells had a reverse impression on the levels of E-cadherin, Ki-67 and vimentin in SW1990 cells (Fig. 2G). To confirm these outcomes further, we extended the study in vivo on mice. The mice were injected with SW1990 cells overexpressing miR-125bi, BxPC-3-miR-125bi or SW1990-miR-NC into the vein of mice. The outcomes suggested that inhibition of miR-125b resulted in suppression of metastasis of PC to the lung (Fig. 2H). All these results confirm that miR-125b induces the migration and growth of pancreatic cancer cells.
MiR-125b promotes the migration and proliferation of pancreatic cancer cells. A: BxPC-3 cells were transfected with miR-125b mimics followed by MTT assay and BrdU assay at various time intervals. B: The BxPC-3 cells were treated for 48 h as described earlier in (A) followed by determination of cell proliferation by crytal violet staining method. C: The SW1990 cells were transfected with miR-125b inhibitor (miRi) followed by MTT analysis and BrdU assay at various time intervals. D: BxPC-3 cells transfected with miRi for 48 h and cell proliferation was estimated by crystal violet staining. E: BxPC-3 cells were treated as described earlier in (A) followed by Transwell migration assay. F: SW1990 cells were transfected with miRi followed by Transwell migration assay. G: Both BxPC-3 and SW1990 cells received transfection of miR or miRi and expression of proteins was studied by western blot analysis. H: The BxPC-3-miR-125bi or BxPC-3-miR-125b-Negative control cells were injected to mice via tail vein. The mice were sacrificed after 10 weeks of injecting cells, the development of metastasis in the lungs was evaluated by H&E staining. ** 0.01 compared to control.
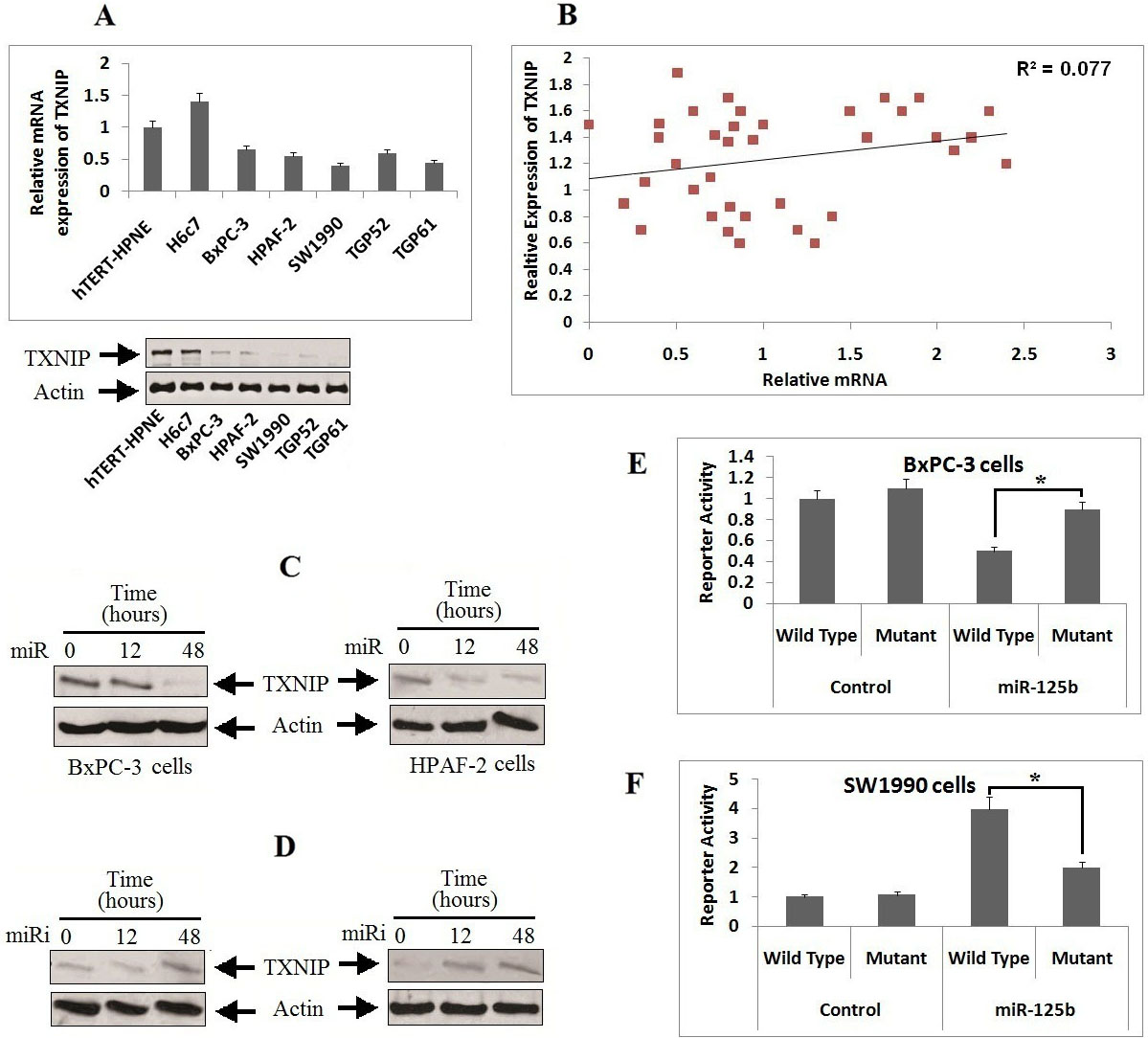
TXNIP is a potential target of miR-125b in pancreatic cancer cells. A: The protein (lower) and mRNA expression (upper) of TXNIP was evaluated in various PC cell lines by qRT-PCR and western blot analyses. Both the levels of mRNA and TXNIP were normalized against the normal pancreatic cells (hTERT-HPNE cells). B: Pearson’s correlation coefficient analysis was done for establishing correlation between miR-125b and TXNIP in PC tissue samples from patients. ( 0.0019). C: BxPC-3 and HPAF-2 cells received transfection of TXNIP and miR-125b, and the expression levels was evaluated by western blot analysis. D: SW1990 and TGP61cells received transfection with miRi-125b and TXNIP, the expression level was evaluated by western blot analysis. E: miR-125b mutant or TXNIP 3-UTR wild type, luciferase activity in BxPC-3 cells received transfection of miR-125b. F: miR-125b mutant or TXNIP 3-UTR wild type, luciferase activity in SW1990 cells received transfection of miR-125b. * 0.05 compared to wild type.
TXNIP was the potential target of miR-125b in pancreatic cancer cells
For identifying the direct target of miR-125b in silico, TargetScan (Algorithm) was employed for identifying the possible mechanism by which miR-125b acts on migration of pancreatic cancer cells. The results of algorithm TargetScan version 7.2 (http://www.targetscan. org/vert_72/) hinted sequence match between miR-125b and 3UTR of TXNIP. We found that expression of protein and mRNA levels TXNIP were suppressed in all the selected five pancreatic cancer cell lines compared to normal cells (Fig. 3A), the results were opposite to the levels of miR-125b in these cells. Further, the elevated levels of miR-125b in both the pancreatic cancer cell lines (BxPC-3 and SW1990 cells) was associated with suppressed expression of TXNIP (Fig. 3A). However, a significantly inverse correlation existed between the expression levels of miR-125b and TXNIP mRNA in pancreatic cancer tissues ( 0.001, Fig. 3B), this outcomes suggested TXNIP may as a potential target of miR-125b in pancreatic cancer. To affirm these outcomes, we evaluated expression of TXNIP in different pancreatic cells treated with miR-125b or miR-125bi. Under these conditions, the levels of TXNIP were suppressed by miR-125b, but the levels were upregulated upon exposure with miR-125b inhibitor (Fig. 3C and D).
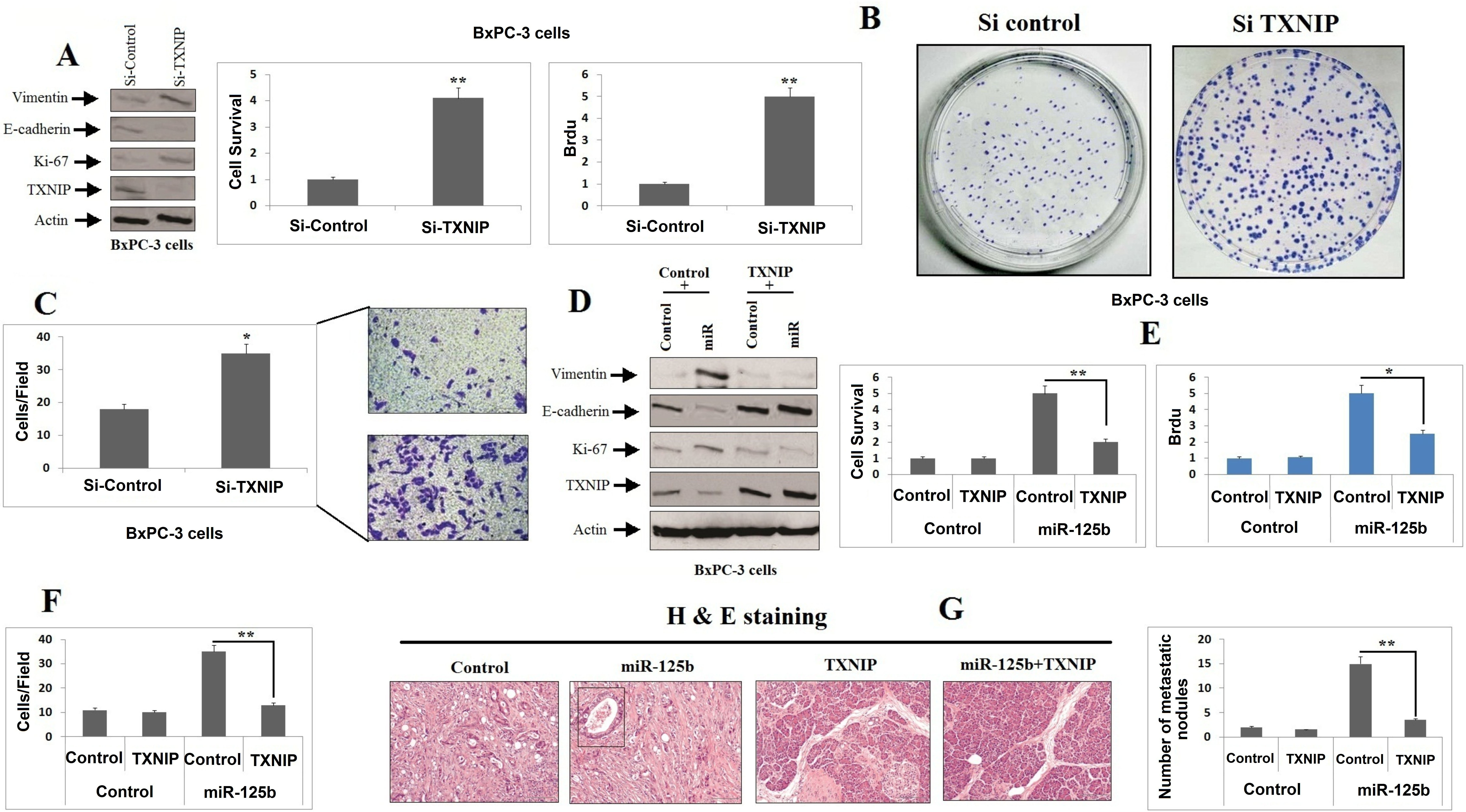
miR-125b encourages cell proliferation by suppressing TXNIP in PC cells. A: BxPC-3 cells received transfection of TXNIP siRNA, the expression proteins was analyzed by western blot analysis. B: MTT assay was carried for studying proliferation of BxPC-3 cells after receiving transfection of TXNIP siRNA treated as in (A) was tested by an MTT assay (left), BrdU assay (middle), and crystal violet staining (right) at 48 h. (C) The migration of BxPC-3 cells treated as in (A) was tested by Transwell assays. (D) BxPC-3 cells were co-transfected with miR and TXNIP plasmid, and the expression of the indicated proteins was analyzed by western blotting. (E) The proliferation of BxPC-3 cells treated as in (D) was tested by MTT (left) and BrdU (right) assays at 48 h. (F) The migration of BxPC-3 cells treated as in (D) was tested by Transwell assays. (G) Histological analysis of the pulmonary metastases in the animal model as described. * 0.05; ** 0.01.
Furthermore, the upregulation of miR-125b resulted in decreased luciferase activity, whereas the activity remained unaffected upon mutation of sequences of 3-UTR for the complementary sites in seed region of miR-125b (Fig. 3E). The miR-125b inhibitor caused a significant increase in Luciferase activity upon transfection with constructed vectors in the pancreatic cancer cells, the outcomes however suggested that luciferase activity remained unchanged when the 3-UTR of TXNIP in the seeding site of miR-125b was mutated (Fig. 3F). The findings confirm TXNIP as a potential target of miR-125b in pancreatic cell lines.
miR-125b encourages the growth and migration of pancreatic cancer via targeting and suppressing TXNIP in vitro and in vivo
The outcomes confirmed that TXNIP is favorable target of miR-125b contributing in development of pancreatic cancer; we supposed that the oncogene character of miR-125b was associated to blockade of TXNIP. The decrease in levels of TXNIP with the presence of two siRNAs also caused the inhibition of E-cadherin and also increased the levels of Ki-67 and vimentin in BxPC-3 cell lines (Fig. 4A). Consequently, the blockade of TXNIP caused migration and proliferation of BxPC-3 cells (Fig. 4B and C), which showed same consequences with over-expression of miR-125b. The results also showed that, upon transfecting TXNIP with miR-125b mimics a reversal in expression of Ki-67, vimentin and E-cadherin (Fig. 4D) was observed along with elevation in growth mediated by miR-125b (Fig. 4E and F).
To further provide concrete to our study in vivo, the mice were injected with BxPC-3-miR125b+TXNIP (miR-125b and TXNIP co-expressing PC cells) and BxPC-3-miR-NC cells (Control cells) via lateral vein. Our findings suggested that, the miR-125b over-expressing group of mice showed higher metastasis in the form of lung nodules upon microscopic examination (Fig. 4G). However, the mice injected with cells exhibiting co-expression of TXNIP along with miR-125b decreased the number of lung nodules (Fig. 4G). All together the in vitro and in vivo experiments suggested that miR-125b contributed in growth and migration of pancreatic cancer via suppressing the levels of TXNIP.
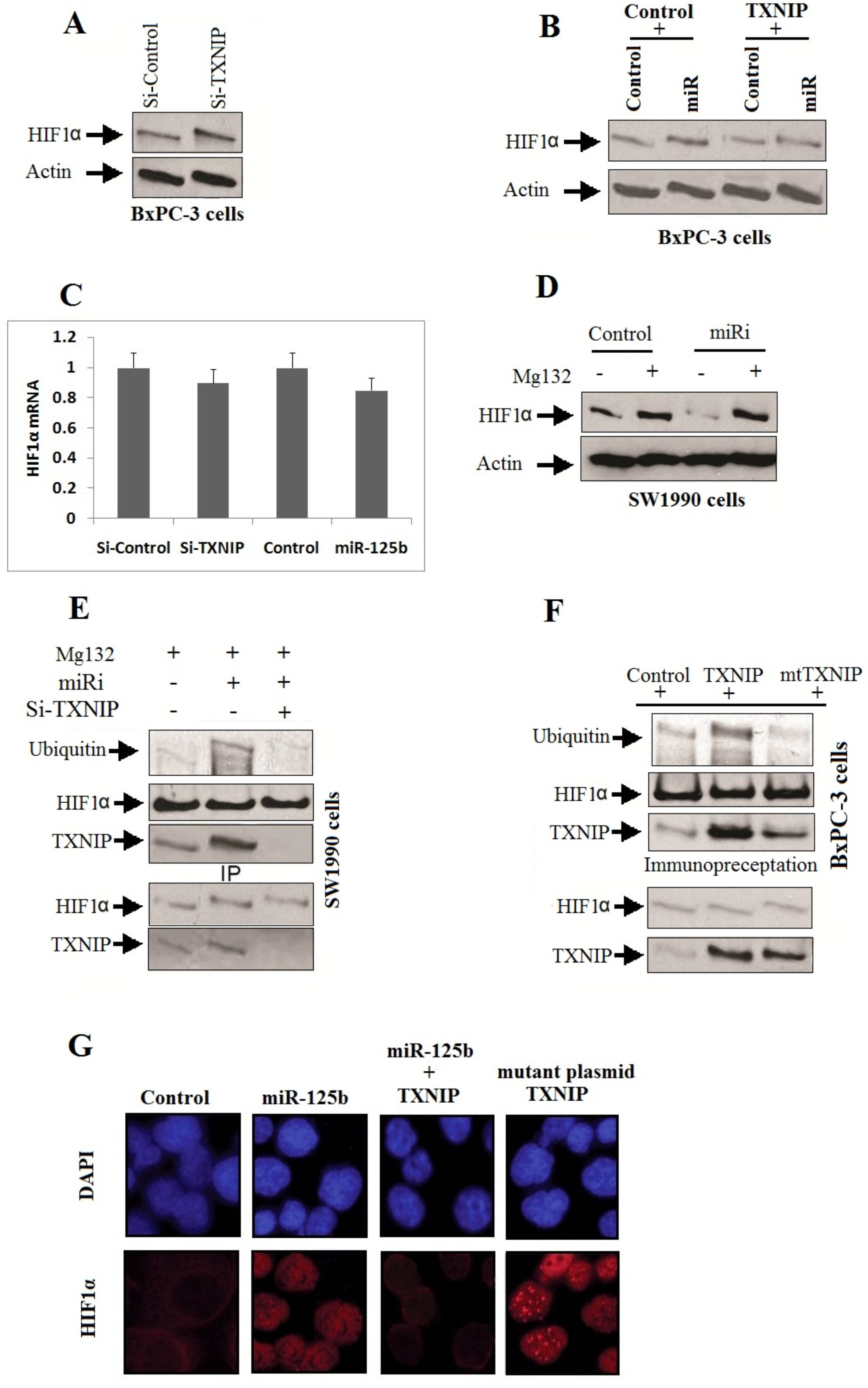
MiR-125b downregulates the expression of TXNIP for activating HIF1. A: BxPC-3 cells received transfection of TXNIP siRNA, the expression of HIF1 was evaluated by western blot analysis. B: The BxPC-3 cells received co-transfection of miR-125b and TXNIP plasmid, the expression of HIF1 was evaluated by western blot analysis. C: The levels of HIF1-mRNA observed after the BxPC-3 cells were transfected with TXNIP siRNA or miR-125b. D: SW1990 cells received pretreatment of protease inhibitor (MG132) followed by transfection with miR-125b inhibitor for next 6 hours, the expression of HIF1 was evaluated by western blot analysis. E: SW1990 cells were treated with protease inhibitor (MG132) for 2 hours followed by co-transfection with miR-125b inhibitor and TXNIP siRNA. The ubiquitylation of HIF1 and interaction between TXNIP and HIF1 were evaluated using anti-HIF1 antibody. F: The BxPC-3cells received pretreatment of MG132 for 2 h followed by co-transfection with miR-125b and TXNIP Wild type or mutant plasmid TXNIP. After 6 hours, ubiquitylation of HIF1 and interaction between TXNIP and HIF1 were evaluated using anti-HIF1 antibody. G: The BxPC-3 cells were co-transfected with miR-125b and TXNIP wild type or mutant plasmid TXNIP. Immunofluorescence study was done for studying the expression and cellular localization of HIF1.
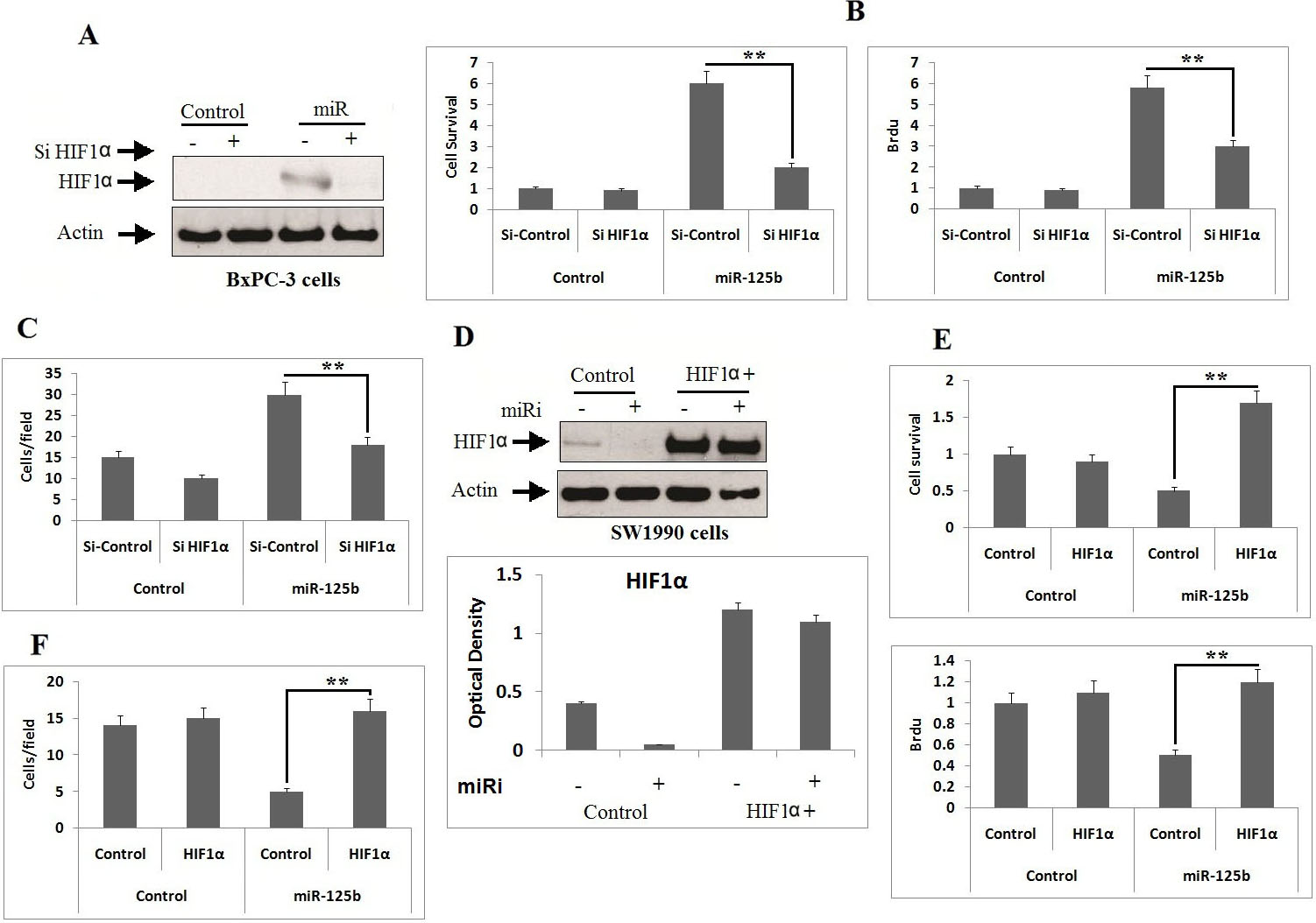
HIF1 is crucial factor for miR-125b in proliferation and migration of PC cell. A: The BxPC-3 cells received transfection of miR-125b and Si-HIF1, the suppressing role of HIF1 was evaluated by western blot analysis. B: MTT and BrdU assay was done after 48 hours for evaluating the proliferation of BxPC-3cells transfected with miR-125b and HIF1 siRNA. ** 0.001 compared to Si-HIF1. C: The extent of cell migration of BxPC-3 cells transfected with miR-125b and Si-HIF1 was analyzed by Transwell assays. ** 0.001 compared to Si-HIF1. D: The SW1990 cells received co-transfection of miR-125b inhibitor and HIF1 plasmid, the expression of HIF1 was evaluated by western blot analysis. E: The extent of proliferation of SW1990 cells co-transfected with miR-125b inhibitor and HIF1 plasmid were analyzed for MTT and BrdU assay after 48 hours of transfection. ** 0.001 compared to control. F: The extent of migration in SW1990 cells co-transfected with miR-125b inhibitor and HIF1 plasmid were tested by Transwell migration assay. ** 0.001 compared to control.
miR-125b stabilizes HIF1 and via TXNIP cascade
HIF1 has been identified to be an important factor in the stimulating migration, proliferation and metastasis of tumors [18], HIF1 is found to be degraded under normal conditions by von Hippel–Lindau protein (pVHL) [19]. In the present study, we showed that, siRNA mediated decrease in levels of TXNIP elevated the levels of HIF1 in BxPC-3 cells (Fig. 5A). Contrarily, the increased levels of TXNIP settled the induction of HIF1 after the transfection of miR-125b (Fig. 5B). Nevertheless, upon blocking TXNIP or by transfection of miR-125b caused no substantial alterations in levels of mRNA of HIF1 (Fig. 5C), which suggested that TXNIP along with miR-125b stabilizes levels of HIF1. The suppression of HIF1 using an inhibitor of miR-125b was reclaimed following pretreatment using MG-132 i.e a protease inhibitor (Fig. 5D). The results of our study further confirmed that, pretreatment of miR-125b inhibitor increased the interaction between HIF1 and TXNIP, and ubiquitylation of HIF1 (Fig. 5E), The blockade of TXNIP halted the ubiquitylation of HIF1, suggesting that the HIF1 and TXNIP interaction leads to its degradation. Looking into the reports suggesting that the CRM1 site is valuable for the building of TXNIP-pVHL-HIF1 complex [20], the SW1990 cells were transfected with TXNIP construct having CRM1 binding site mutation or a TXNIP construct (Wild type). The outcomes showed that, elevated expression of TXNIP construct (Wild type) enhanced the interaction with HIF1 and its ubiquitylation, which was not shown by the transfected mutant TXNIP (Fig. 5F). In addition to this, the transfection of miR-125b resulted in stabilization and translocation of HIF1 in the nucleus, which was blocked when transfected with TXNIP (Wild type) and not with the mutant (Fig. 5G). All together, the experiment suggested that miR-125b suppresses the expression of TXNIP leading to stabilization and release of HIF1.
HIF1 acts a crucial factor in miR-125b arbitrated growth and migration of pancreatic cells
To evaluate the involvement of HIF1 in miR-125b induced cell growth, proliferation and migration. As expected, up-regulation of miR-125b in BxPC-3 cell lines enhanced the levels of HIF1, which in turn was associated for the increase in migration and proliferation of PC cells (Fig. 6A–C). On the other hand, blockade of HIF1 settled the growth mediated by transfection of miR-125b (Fig. 6A–C) and down-regulation of miR-125b with its inhibitor suppressed the migration, proliferation and expression levels of HIF1 in cells (Fig. 6D–F). The outcomes suggest that miR-125b elevates the levels of HIF1 via suppressing TXNIP, which is crucial for the growth and migration of pancreatic cancer cells
Discussion
Presently, selective expression of miRs in cancerous and non cancerous biological samples showing specific prognostic features has been employed as potential predictive markers, they also serves as targets for treating malignancies [20, 21]. Reports have emerged suggesting profiling of several miRs in pancreatic cancer tissues [6], however there are no reports describing the correlation between miR-125b and pancreatic cancer. In the present work, we showed that level of miR-125b was increased in pancreatic cancer tissue samples compared to normal tissues. Our findings demonstrated that miR-125b resulted in development and progression of pancreatic cancer, increased migration and proliferation of pancreatic cancer cells in vitro and also in animal model in vivo. The results hence confirmed that miR-125b acted as oncogenes promoting development and progression of PC. The outcomes were consistent with the function of miR-125b in other studies associated with malignancies such as Glioma [22, 26] and prostate cancer [27]. Though over-expression of miR-125b is observed in various malignancies such [22, 26, 27], the underlying mechanism involved in cancer are still unclear. In the present study, we established a novel mechanism describing role of miR-125b in PC, suggesting that miR-125b leads to cell growth in PC via miR-125b-TXNIP- HIF1 pathway. Here we also demonstrated that TXNIP was the favorable target of miR-25b and is responsible for the suppression of TXNIP in pancreatic cancer cells. We also proved that this suppression of TXNIP mediated by miR-125b may promote growth of pancreatic cancer. The outcomes also showed that miR-125b mediated suppression of TXNIP causes nuclear translocation of very important HIF1 (cancer progression stimulator).
Reports have confirmed that blocking TXNIP is associated with advanced stage various cancers [19, 28, 29], but the studies related to genetic alterations of TXNIP are missing. In the present work, we showed that miR-125b potentially aims TXNIP and inhibits its regulation in PC. TXNIP have shown suppressive effect on cancer metastasis [18, 30]. Parallel to this, in the present work it was found that elevated expression of TXNIP halted the miR-125b mediated cell proliferation and metastasis in PC. TXNIP is a protein that blocks thioredoxin, it was evidenced previously that thioredoxin causes promotion for progression of tumor by triggering angiogenesis and blocking apoptosis [31, 32, 33]. These reports confirm TXNIP as a potential tumor suppressor gene. Numbers of studies have come up suggesting effects of TXNIP in various pathways [34, 35, 20]. Moreover, many reports also suggest that over-expression of TXNIP inhibits the hypoxia induced factor mediated activity in various malignancies [19, 34, 36]. The studies also established that the CRM1 region of TXNIP binds to NF- B-HIF1 and pVHL forming complex which causes degradation of HIF1 [26]. In light of these findings, the present study demonstrated that TXNIP regulates the expression of HIF1 negatively, irrespective of its interaction with reactive oxygen species and thioredoxin as there was no notable change in mRNA levels of HIF1. In contradiction to this, the findings of our study suggested that TXNIP forms a complex with HIF1 causing degradation, blockade and its translocation in nucleus.
Attenuation of HIF1 is necessary for managing the regulation of number of genes responsible for reaction involving hypoxia, transport of oxygen, angiogenesis, glucose metabolism which is responsible for proliferation, migration and tumor invasion. HIF1 has been recorded to regulate various pathways in cells such as translation, transcription and degradation [37, 38]. Studies have emerged establishing various miRs responsible for modulating expression of HIF1 [39, 40]. In the current study we proved that miR-125b elevated the levels of HIF1 and caused activation of HIF1 via TXNIP pathway. The expression levels of TXNIP halts the up-regulation of HIF1 mediated by miR-125b. In contrast to previous reports, which suggest that miR-373 resulted in activation of HIF1 via scavenging of reactive oxygen species, the outcomes of present study suggest that miR-125b induce the expression of HIF1 via increasing the stability and its translocation. It has been demonstrated that degradation and localization of HIF1 happens based on type of cells majorly in cytoplasm, similarly in I endothelial cells the degradation occurs in cytoplasm and in nucleus. Hence, the results of our study demonstrate that the miR-125b-TXNIP-HIF1 pathway is potential for growth and migration of pancreatic cancer cells, creating new strategies for treating pancreatic cancer.
In conclusion, the outcomes indicate that miR-125b drives the proliferation and migration of pancreatic cancer cells. MiR-125b promotes the metastasis and progression of PC by stabilizing HIF1 via TXNIP cascade in pancreatic cancer, this pathway can be employed as possible target for diagnosing and treating pancreatic cancer.
Footnotes
Conflict of interest
The authors declare no conflict of interest.
References
1.
MatsumotoK.TakedaY.OnoyamaT.KawataS.KurumiH.KodaH.YamashitaT. and IsomotoH., Endoscopic treatment for distal malignant biliary obstruction, Ann Transl Med5 (2017), 190.
2.
ChrystojaC.C.DiamandisE.P.BrandR.RuckertF.HaunR. and MolinaR., Pancreatic cancer, Clin Chem59 (2013), 41–46.
3.
BournetB.BuscailC.MuscariF.CordelierP. and BuscailL., Targeting KRAS for diagnosis, prognosis, and treatment of pancreatic cancer: Hopes and realities, Eur J Cancer54 (2016), 75–83.
4.
DengT.YuanY.ZhangC.ZhangC.YaoW.WangC.LiuR. and BaY., Identification of circulating miR-25 as a potential biomarker for pancreatic cancer diagnosis, Cell Physiol Biochem39 (2016), 1716–1722.
5.
JafarnejadS.M. and LiG., Regulation of p53 by ING family members in suppression of tumor initiation and progression, Cancer Metastasis Rev31 (2012), 55–73.
6.
BartelD.P., MicroRNAs: Genomics, biogenesis, mechanism, and function, Cell116 (2004), 281–297.
7.
WinterJ.JungS.KellerS.GregoryR.I. and DiederichsS., Many roads to maturity: microRNA biogenesis pathways and their regulation, Nat Cell Biol11 (2009), 228–234.
8.
YonemoriK.KuraharaH.MaemuraK. and NatsugoeS., MicroRNA in pancreatic cancer, J Hum Genet62 (2017), 33- 40.
9.
ZhouB.SunC.HuX.ZhanH.ZouH.FengY.QiuF.ZhangS.WuL. and ZhangB., MicroRNA-195 suppresses the progression of pancreatic cancer by targeting DCLK1 cell physio biochem44 (2017), 1867–1881.
10.
QianY.FengL.WuW.WengT.HuC.HongB.WangF.X.ShenL.WangQ. and JinX., MicroRNA expression profiling of pancreatic cancer cell line L3 6p1 following B7-H4 knockdown, Cell Physio Biochem44 (2017), 494–504.
11.
YangW.YangY.XiaL.YangY.WangF.SongM.ChenX.LiuJ.SongY. and ZhaoY., MiR-221 promotes Capan-2 pancreatic ductal adenocarcinoma cells proliferation by targeting PTEN-Akt, Cell Physio Biochem38 (2016), 2366–2374.
12.
ZhaoL.SunH.KongH.ChenZ.ChenB. and ZhouM., The Lncrna-TUG1/EZH2 axis promotes pancreatic cancer cell proliferation, migration and EMT phenotype formation through sponging Mir-382, Cell Physio Biochem42 (2017), 2145–2158.
13.
LiH.HaoX.WangH.LiuZ.HeY.PuM.ZhangH.YuH.DuanJ. and QuS., Circular RNA expression profile of pancreatic ductal adenocarcinoma revealed by microarray, Cell Physio Biochem40 (2016), 1334–1344.
14.
ShiL.ZhangJ.PanT.ZhouJ.GongW.LiuN. et al., MiR-125b is critical for the suppression of human U251 glioma stem cell proliferation, Brain Res1312 (1312) 120–126.
15.
CuiY.XiaoZ.HanJ.SunJ.DingW.ZhaoY. et al., MiR-125b orchestrates cell proliferation, differentiation and migration in neural stem/progenitor cells by targeting Nestin, BMC Neurosci13 (2012) 116.
16.
WanY.SunG.WangZ.GuoJ. and ShiL., miR-125b promotes cell proliferation by directly targeting Lin28 in glioblastoma stem cells with low expression levels of miR-125b, Neuroreport25(5) (2014), 289–296.
17.
JayachandranA.DhungelB. and SteelJ.C., Epithelial-to-mesenchymal plasticity of cancer stem cells: Therapeutic targets in hepatocellular carcinoma, J Hematol Oncol9 (2016), 74.
18.
SchofieldC.J. and RatcliffeP.J., Oxygen sensing by HIF hydroxylases. Nature reviews, Molecular Cell Biology5 (2004), 343–354.
19.
ShinD.JeonJ.H.JeongM.SuhH.W.KimS.KimH.C.MoonO.S.KimY.S.ChungJ.W.YoonS.R.KimW.H. and ChoiI., VDUP1 mediates nuclear export of HIF-1alpha via CRM1-dependent pathway, Biochimica et Biophysica Acta1783 (2008), 838–848.
20.
SpindelO.N.WorldC. and BerkB.C., Thioredoxin interacting protein: Redox dependent and independent regulatory mechanisms, Antioxid Redox Signal16 (2012), 587–596.
21.
GarzonR. and MarcucciG., Potential of microRNAs for cancer diagnostics, prognostication and therapy, Current Opinion in Oncology24 (2012), 655–659.
22.
AlbulescuR.NeaguM.AlbulescuL. and TanaseC., Tissular and soluble miRNAs for diagnostic and therapy improvement in digestive tract cancers, Expert Review of Molecular Diagnostics11 (2011), 101–120.
23.
ShiL.ZhangJ.PanT.ZhouJ.GongW.LiuN.FuZ. and YouY., MiR-125b is critical for the suppression of human U251 glioma stem cell proliferation, Brain Res1312 (2010), 120–126.
24.
ShiL.ZhangS.FengK.WuF.WanY.WangZ.ZhangJ.WangY.YanW.FuZ. and YouY., MicroRNA-125b-2 confers human glioblastoma stem cells resistance to temozolomide through the mitochondrial pathway of apoptosis, Int J Oncol40(1) (2012), 119–129.
25.
ShiX.B.XueL.MaA.H.TepperC.G. and KungH.J., White RW. miR-125b promotes growth of prostate cancer xenograft tumor through targeting pro-apoptotic genes, Prostate71 (2011).
26.
BaiJ.NaH.HuaX.WeiY.YeT.ZhangY.JianG.ZengW.YanL. and TangQ., A retrospective study of NENs and miR-224 promotes apoptosis of BON-1 cells by targeting PCSK9 inhibition, Oncotarget8 (2017), 6929–6939.
27.
HeC.WangL.ZhangJ. and XuH., Hypoxia-inducible microRNA-224 promotes the cell growth, migration and invasion by directly targeting RASSF8 in gastric cancer, Molecular cancer16 (2017), 35.
28.
NishizawaK.NishiyamaH.MatsuiY.KobayashiT.SaitoR.KotaniH.MasutaniH.OishiS.TodaY.FujiiN.YodoiJ. and OgawaO., Thioredoxin-interacting protein suppresses bladder carcinogenesis, Carcinogenesis32 (2011), 1459–1466.
29.
WoolstonC.M.MadhusudanS.SoomroI.N.LoboD.N.Reece-SmithA.M.ParsonsS.L. and MartinS.G., Thioredoxin interacting protein and its association with clinical outcome in gastro-oesophageal adenocarcinoma, Redox Biology1 (2013), 285–291.
30.
SchofieldC.J. and RatcliffeP.J., Oxygen sensing by HIF hydroxylases. Nature reviews, Molecular Cell Biology5 (2004), 343–354.
31.
NishiyamaA.MatsuiM.IwataS.HirotaK.MasutaniH.NakamuraH.TakagiY.SonoH.GonY. and YodoiJ., Identification of thioredoxin-binding protein-2/vitamin D(3) up-regulated protein 1 as a negative regulator of thioredoxin function and expression, J Biol Chem274 (1999), 21645–21650.
32.
WelshS.J.BellamyW.T.BriehlM.M. and PowisG., The redox protein thioredoxin-1 (Trx-1) increases hypoxia-inducible factor 1alpha protein expression: Trx-1 overexpression results in increased vascular endothelial growth factor production and enhanced tumor angiogenesis, Cancer Res62 (2002), 5089–5095.
33.
SaitohM.NishitohH.FujiiM.TakedaK.TobiumeK.SawadaY.KawabataM.MiyazonoK. and IchijoH., Mammalian thioredoxin is a direct inhibitor of apoptosis signal-regulating kinase (ASK) 1, EMBO J17 (1998), 2596–2606.
34.
ChenD.DangB.L.HuangJ.Z.ChenM.WuD.XuM.L.LiR. and YanG.R., MiR-373 drives the epithelial-to-mesenchymal transition and metastasis via the miR-373-TXNIP-HIF1alpha-TWIST signaling axis in breast cancer, Oncotarget6 (2015), 32701–32712.
35.
WaldhartA.N.DykstraH.PeckA.S.BoguslawskiE.A.MadajZ.B.WenJ.VeldkampK.HollowellM.ZhengB.CantleyL.C.McGrawT.E. and WuN., Phosphorylation of TXNIP by AKT mediates acute influx of glucose in response to insulin, Cell Rep19 (2017), 2005–2013.
36.
WongR.W. and HagenT., Mechanistic target of rapamycin (mTOR) dependent regulation of thioredoxin interacting protein (TXNIP) transcription in hypoxia, Biochem Biophys Res Commun433 (2013), 40–46.
37.
YangM.H.WuM.Z.ChiouS.H.ChenP.M.ChangS.Y.LiuC.J.TengS.C. and WuK.J., Direct regulation of TWIST by HIF-1alpha promotes metastasis, Nat Cell Biol10 (2008), 295–305.
38.
GaoP.ZhangH.DinavahiR.LiF.XiangY.RamanV.BhujwallaZ.M.FelsherD.W.ChengL.PevsnerJ.LeeL.A.SemenzaG.L. and DangC.V., HIF-dependent antitumorigenic effect of antioxidants in vivo, Cancer Cell12 (2007), 230–238.
39.
MadaneckiP.KapoorN.BebokZ.OchockaR.CollawnJ.F. and BartoszewskiR., Regulation of angiogenesis by hypoxia: The role of microRNA, Cell Mol Biol Lett18 (2013), 47–57.
40.
GrecoS. and MartelliF., MicroRNAs in hypoxia response, Antioxid Redox Signal21 (2014), 1164–1166.