Patients with acute leukemia (AL) refractory to induction or reinduction chemotherapy show poor prognoses if they do not undergo allogeneic hematopoietic stem-cell transplantation (AHSCT). The present study aims to investigate whether donor natural killer (NK) cells and interleukin-2 (IL-2) gene modification exert anti-relapse effects on AHSCT after establishing a mouse model of AL.
METHODS:
C57BL/6 (H-2) mice were selected as donor mice to obtain NK cells and hematopoietic stem cells, while BALB/c (H-2) mice were selected as the recipient mice for AHSCT. The AHSCT-treated mice were then injected with the donor NK cells, recombinant adenovirus expressing IL-2 (AdIL-2), or the NK cells infected by AdIL-2. Flow cytometry was performed to detect the cell transplantation rate, immune cell number, and cell immunogenicity. Enzyme-linked immunosorbent assay (ELISA) was employed to quantify the secretion of IL-2 in spleen cells, and the level of peripheral blood factors, including interferon- (IFN-), tumor necrosis factor- (TNF-), IL-35, transforming growth factor (TGF-), and IL-10.
RESULTS:
In our experiments, promotional effects of NK cells and AdIL-2 were found on cell transplantation rate, immune reconstitution ability, cell immunogenicity, IL-2 secretion, as well as increased peripheral blood factor levels in the recipient mice treated with AHSCT, with improved pathological changes observed. Moreover, the aforementioned changes were further promoted in the AHSCT-treated recipient mice injected with the AdIL-2-infected NK cells.
CONCLUSIONS:
These results uncover that the donor NK cells and IL-2 gene modification could inhibit the relapse of AL mice underwent AHSCT, hereby providing a new target for leukemia treatment.
Acute leukemia (AL) is a malignant disease of the bone marrow and blood with an alarming prevalence worldwide, which is characterized by an uncontrolled accumulation of blood cells [1]. It could be fundamentally categorized into two types: lymphoblastic leukemia (ALL) and acute myeloid leukemia (AML) [2]. Unsatisfactory outcomes were reported in AML patients older than 60 years, including low remission rates and poor overall survival [3]. A few risk factors have been demonstrated to be associated with AL, among which genetics and environmental exposure are imperative [4]. Currently, allogeneic hematopoietic stem cell transplantation (AHSCT) has been established as an effective therapeutic protocol for patients diagnosed with several hematologic malignancies, such as bone marrow failure syndromes and congenital immune deficiencies [5]. AML recurrence has been a persistent cause of treatment failure after AHSCT [6]. Occasionally, after chimerism, the donor’s hematopoietic cells completely replace the recipient’s hematopoietic cells; but in some cases, a concurrent relationship was established between them in the blood. This could potentially trigger graft-vs-host disease (GVHD) and further lead to AML relapse [7, 8]. Therefore, it warrants an investigation to find new options to rectify treatment failure due to recurrence following AHSCT [6].
Natural killer (NK) cells, which critically are cytotoxic lymphocytes, serve as a bridge between the innate immune system and adaptive immune systems [9]. NK cells activate numerous immune-receptors and trigger them to eliminate the infected, transformed, or stressed cells, and effectively produce interferon- (IFN-) [10]. They moderately contribute to immune defense via their effector functions, including cytotoxicity and cytokine secretion, and relatively function as regulators of the antigen-presenting cells (APC) and adaptive responses of T cells [11]. NK cells can critically influence AHSCT thus helping prevent leukemia relapse, rejection, and GVHD [12]. Interestingly, in recent years, genome-wide analysis has revealed an association between AL and genetic alterations, hereby speculating gene modification as a new direction for research [13]. Interleukin-2 (IL-2), a pleiotropic cytokine produced by T cells after antigen activation, involves in mediating T cell growth and proliferation [14]. Previous studies have revealed the functionality of IL-2 as a regulator of immune response [15, 16]. Based on a study of patients who underwent AHSCT and treated with IL-2, increased Treg expansion and NK cells were found without inducing GVHD [17]. Although extensive research has highlighted the effect of donor NK cells and IL-2 gene modification on AL respectively [18, 19, 20], little attention has been dedicated to the role of donor NK cells in combination with IL-2 gene modification on AHSCT in AL. Consequently, a study is trivial to explore the anti-relapse effects of donor NK cells and IL-2 gene modification on AHSCT in a mouse model of AL, and we hope this study could introduce a new target for superior treatment protocols for leukemia relapse after AHSCT.
Procedures for recipient mice in each group
Group
Procedure
AHSCT
Tail vein injection of 1 10 hematopoietic stem cells
AHSCT NK
Tail vein injection of 1 10 hematopoietic stem cells and 5 10 NK cells
AHSCT AdIL-2
Tail vein injection of 1 10 hematopoietic stem cells, followed by administration of 1 ug AdIL-2 per day for
7 days
AHSCT NK AdIL-2
Tail vein injection of 1 10 hematopoietic stem cells and 5 10 AdIL-2-infected NK cells
The study was conducted with approval of the Animal Ethics Committee of the First Affiliated Hospital of Hebei North University. Animal experiments were conducted in strict accordance with the animal protocols and guidelines established by the Medical Ethics Review committee for animal experiments [21]. Optimal measures were taken to minimize the suffering of animals during the experiment.
Study subjects
Sixty specific pathogen free (SPF) grade BALB/C (H-2) mice (age: 7–8 weeks; weight: 20–24 g) were chosen as the recipient mice (30 male and 30 female) and 60 SPF grade C57BL/6 (H-2) mice (age: 7–8 weeks; weight: 20–24 g) were chosen as the donor mice (30 males and 30 females). The mice were housed in a clean animal facility with Co radiation sterilization, which were fed without any restrictions having a normal circadian rhythm at room temperature of 22–25C. All animals were purchased from Shanghai SLAC Laboratory Animal Co. Ltd. (Shanghai, China). The recombinant adenovirus carrying mouse IL-2 gene (AdIL-2) was provided by the Department of molecular biological therapy, Japan Cancer Research Association.
Cell preparation, transplantation and grouping
Thirty C57BL/6 (H-2) mice were euthanized by cervical dislocation to isolate the femur, tibia, and spine under sterile conditions, followed by grounding and filtration through nylon net to prepare cell suspension. Red blood cells were lysed using red blood cell lysis buffer and rinsed in phosphate buffer saline (PBS). Following this, cell density was adjusted to 1 10 cells/mL to prepare hematopoietic stem cells. The remaining 30 C57BL/6 mice were also euthanized by cervical dislocation to harvest the femur, tibia, spine, and spleen under sterile conditions, which were also prepared as cell suspension, followed by centrifugation at 300 g to pellet the cells. Subsequently, NK cells were isolated according to previous reported protocols [22]. In brief, the obtained cells were depleted for T cells using CD90 (Thy-1.2) MicroBeads and LD depletion column placed in MidiMACS (Miltenyi Biotec, Bergisch-Gladbach, Ger- many). Then, the CD90-depleted cells were positively selected with antiCD49b (DX5) MicroBeads and MS column using MiniMACS magnetic field, which could obtain 95% highly-enriched NK cells. The enriched NK cells were then resuspended in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% fetal calf serum (FCS), 50 lM 2-ME, 10 mM HEPES, 2 mM l-glutamine, 10 ng/mL rhIL-2, and 100 ng/mL rhIL-15 (Gibco, Grand Island, NY, USA). Subsequently, AdIL-2 and donor NK cells were introduced to the culture dish at a ratio of 1 g: 1 mL to prepare AdIL-2-infected NK cells. The cell density of the donor NK cells injected into the recipient mice was identified by cell counting (5 10 cells/mouse). The BALB/c (H-2) mice served as the recipient mice were subjected to AHSCT via tail vein injection of the prepared hematopoietic stem cells (1 10/mouse), which were then assigned into 4 groups (15 mice in each group): AHSCT group, AHSCT NK group, AHSCT AdIL-2 group, and AHSCT AdIL-2 NK group. Detailed procedures for each group are listed in Table 1. After surgery, all recipient mice were housed in a quiet and hygienic facility for four months before euthanasia.
Flow cytometry
Detection of cell transplantation rate
Dimethylsulfoxide (DMSO)-dissolved carboxyfluorescein diacetate succinimidyl (CFSE) (Molecular Probes Inc., Eugene, Oregon, USA) was diluted to 5 mol/L CFSE work fluid using serum free DMEM (Gibco). Mouse NK cell suspension was prepared and added with an equal volume of the CFSE working fluid. After incubation at 37C for 10 min, the reaction was terminated by the addition of 40% cold fetal bovine serum (FBS). Then, the cells were incubated with the anti-H2KPE antibody (BD Pharmingen, San Jose, CA, USA) for 20 min followed by detection using flow cytometry (BD Pharmingen). The cell transplantation rate of the recipient mice was analyzed using Cell quest software (Cell quest, Largo, FL, USA). The experiment was conducted three times independently.
Measurement of immune cell number
After 4 months of AHSCT, the recipient mice were euthanized, with spleens harvested to prepare splenocyte suspension. Fluorescence-activated cell sorting (FACS) buffer (PBS containing 0.5% BSA and 0.1% sodium azide; BD Pharmingen) was added to resuspend the splenocytes, followed by cell counting. Then, the splenocytes were incubated with CD16/CD32 antibodies (BD Pharmingen) at 4C for 20 min, and further incubated with phycoerythrin (PE) or fluorescein isothiocyanate (FITC)-labeled anti-CD3, anti-CD4, anti-CD8, anti-NK1.1, anti-CD11b, anti-CD19, anti-H-2K, anti-H-2K antibodies (BD Pharmingen) at 4C for 20 min. After being centrifuged, precipitated, washed using FACS buffer, and resuspended, the cells were detected by flow cytometry. Cells were sorted at a speed of 4000 cells/second and analyzed by cell quest software. The number of detected cells was no less than 2000. The number of CD T cells, CD T cells, B cells, NK cells, and macrophage cells was further examined using the statistic software CS2000. The experiment was conducted three times independently.
Detection of cell immunogenicity
After 4 months of AHSCT, the recipient mice were euthanized, with bone marrows collected. The bone marrows were divided into 3 tubes and each tube was supplemented with the fluorescent antibodies (BD Pharmingen), including major histocompatibility complex class II (MHCII) antibody (MHCII-APC), co-stimulatory molecule (CD40L and CD28) antibodies (CD40L-APC and CD28-APC), and adherent molecule intercellular adhesion molecule (ICAM) antibody (ICAM-APC). After incubation for 30 min at room temperature in the dark, red blood cell lysis was added into each tube, followed by centrifugation at 402 g for 5 min. Finally, the cells were analyzed using flow cytometry. The experiment was conducted three times independently.
Enzyme-linked immunosorbent assay (ELISA)
Detection of IL-2 secretion in spleen cells
The splenocytes of recipient mice were isolated, ground, and centrifuged to obtain supernatant. The level of IL-2 in the supernatant was measured according to the instructions of ELISA kit (eBioscience, San Diego, CA, USA). ELISA kit was equilibrated at room temperature for 20 min, and then utilized to prepare the washing buffer. After dissolution, 100 L of standard sample was added to the reaction plate for incubation at 37C for 90 min to draw standard curve. Subsequently, 100 L of biotinylated antibody solution was added for incubation at 37C for 60 min and 100 L of enzyme binding reactant solution was added for incubation at 37C for 30 min, followed by addition of 100 L of tetramethylbenzidine (TMB) substrate. After reaction termination, the optical density (OD) value was detected using a multi-mode microplate reader (BioTek Synergy 2, BioTek Instruments, Inc., Winooski, VT, USA) at an excitation wavelength of 450 nm. The standard curve was plotted based on the OD values to analyze the IL-2 secretion in splenocytes of recipient mice. The experiment was conducted three times independently.
Detection of peripheral blood factor level
After 4 months of AHSCT, orbital blood was collected from recipient mice before euthanasia. Then, the peripheral blood supernatant was used for ELISA detection. The detail procedures were the same as above mentioned. Enzyme labelled antibodies including IFN-, TNF-, IL-35, TGF-, and IL-10 were used. The experiment was conducted three times independently.
Hematoxylin and eosin (HE) staining
The liver, colon, and skin samples were harvested from the euthanized recipient mice. These samples were fixed using 4% formalin, dehydrated using gradient ethanol, embedded in paraffin, and sectioned into 5 m. The sections were stained using HE (hematoxylin staining for 3 min and eosin counterstaining for 3 min) (Boster Biotechnology, Wuhan, Hubei, China). The stained sample sections were then mounted and observed under the microscope (Olympus, Tokyo, Japan). The experiments were conducted three times independently.
Statistical analyses
Data were analyzed using SPSS version 21.0 (IBM Corp, Armonk, NY, USA). Measurement data were expressed as mean standard deviation, which were analyzed by test. Enumerated data were expressed as ratios and percentages, which were compared using Chi-square test. Differences among multiple groups were analyzed by one-way analysis of variance (ANOVA), followed by Tukey’s post hoc test. A value of 0.05 was considered to be statistically significant.
Donor NK cells and AdIL-2 promoted the transplantation rate of AHSCT in recipient mice. A, the transplantation rate of AHSCT in each group detected by flow cytometry; B, statistical analysis of Panel A; *, 0.05, vs. the AHSCT group; #, 0.05, vs. the AHSCT NK group; &, 0.05, vs. the AHSCT AdIL-2 group; Differences among groups were analyzed by one-way analysis of variance, followed by Tukey’s post hoc test. AHSCT, allogeneic hematopoietic stem cell transplantation; NK: natural killer; AdIL-2, adenovector-mediated delivery of interleukin-2.
Results
Donor NK cells and AdIL-2 promote transplantation rate of AHSCT
The transplantation rate of AHSCT in recipient mice was measured by FITC-H-2K and PE-H-2K flow cytometry. The results indicated that, compared with the AHSCT group, the transplantation rate of AHSCT was significantly increased in the AHSCT NK, AHSCT AdIL-2, and AHSCT NK AdIL-2 groups (all 0.05), while no significant difference was observed between the AHSCT NK group and the AHSCT AdIL-2 group ( 0.05). The transplantation rate in the AHSCT NK AdIL-2 group was significantly higher than that in the AHSCT NK and AHSCT AdIL-2 groups (all 0.05) (Fig. 1A and B). These data revealed that donor NK cells and IL-2 gene modification could improve the transplantation rate of AHSCT.
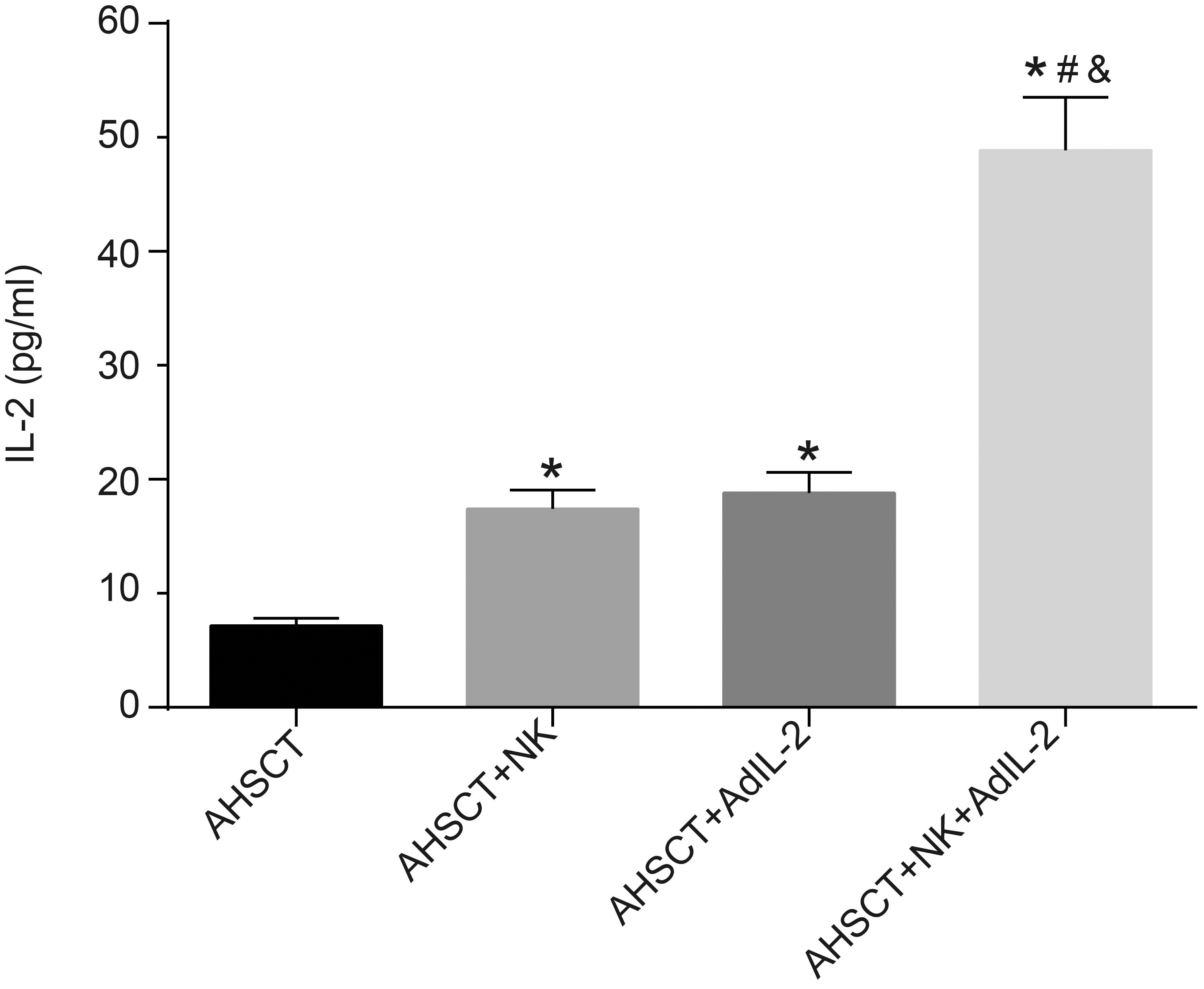
Donor NK cells and AdIL-2 promote IL-2 secretion in splenocytes of recipient mice. The IL-2 secretion in each group detected by ELISA; *, 0.05, vs. the AHSCT group; #, 0.05, vs. the AHSCT NK group; &, 0.05, vs. the AHSCT AdIL-2 group; Differences among groups were analyzed by one-way analysis of variance, followed by Tukey’s post hoc test. AHSCT: allogeneic hematopoietic stem cell transplantation; NK: natural killer; AdIL-2, adenovector-mediated delivery of interleukin-2.
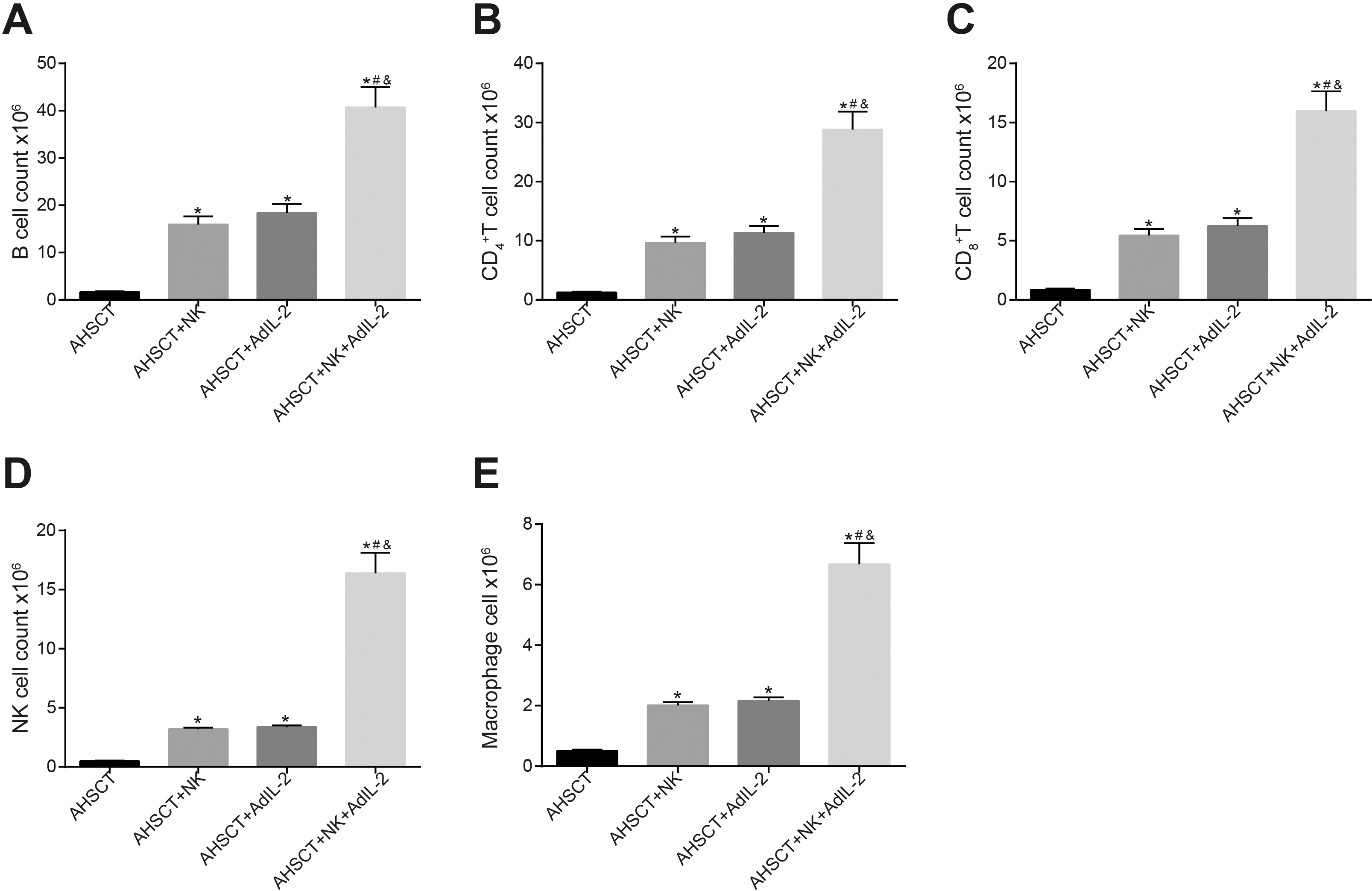
Donor NK cells and AdIL-2 could promote spleen immune reconstitution of recipient mice. Panel A–E: the number of B cells, CD T cells, CD T cells, NK cells and macrophage cells in recipient mice after transplantation detected by flow cytometry is increased by IL-2 recombinant adenovirus infected donor NK cells; *, 0.05, vs. the AHSCT group; #, 0.05, vs. the AHSCT NK group; &, 0.05, vs. the AHSCT AdIL-2 group; Differences among groups were analyzed by one-way analysis of variance, followed by Tukey’s post hoc test. AHSCT: allogeneic hematopoietic stem cell transplantation; NK: natural killer; AdIL-2, adenovector-mediated delivery of interleukin-2.
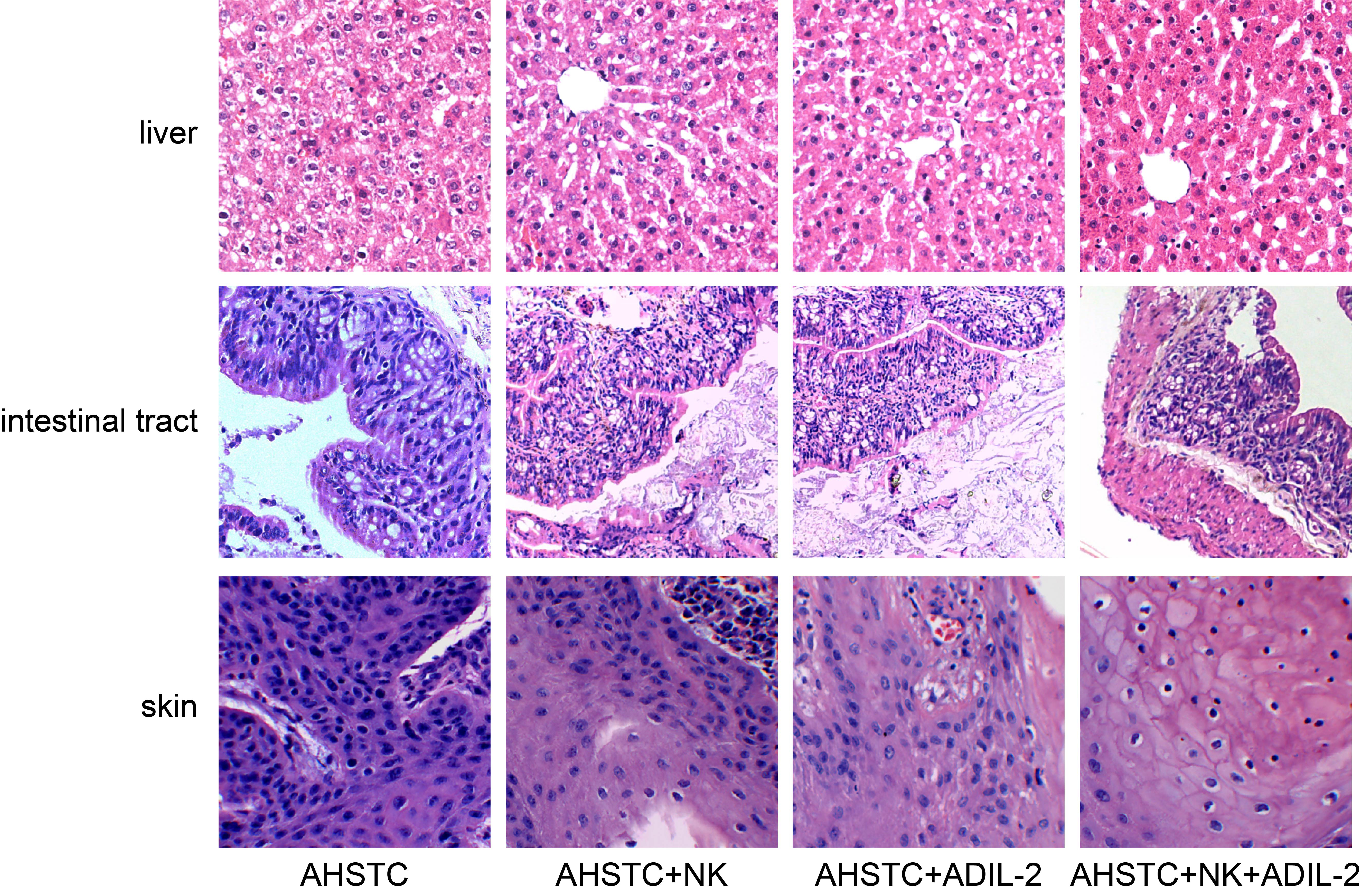
Donor NK cells and IL-2 gene modification inhibit the deterioration of tissue pathogenesis in recipient mice receiving AHSCT ( 400). AHSCT group: hydropic degeneration of hepatocytes, apparent lymphocytic infiltration in the portal area, sinus expansion and hyperemia in the hepatic vascular, edema in the intestinal submucosa, structure destruction accompanied by a lymphocyte infiltration in the lamina propria, severe vacuolar degeneration in the epidermal layer, focus dissolution in the basal layer, and apparent lymphocytic infiltration. The AHSCT NK, AHSCT AdIL-2, and AHSCT NK AdIL-2 groups showed better hydropic degeneration in mouse hepatocytes and lymphocyte infiltration, smaller intestinal structure and lymphocyte infiltration in the lamina propria, as well as lower vacuolar degeneration of the epidermal layer and focus dissolution of the basal layer. AHSCT: allogeneic hematopoietic stem cell transplantation; NK: natural killer; AdIL-2, adenovector-mediated delivery of interleukin-2.
Donor NK cells and AdIL-2 promote IL-2 secretion in splenocytes of recipient mice
To evaluate the level of allogeneic reaction in the recipient mice, ELISA was adopted to detect the content of IL-2 secreted by the mouse splenocytes. The results demonstrated a notable increased IL-2 secretion in the AHSCT NK, AHSCT AdIL-2, and AHSCT NK AdIL-2 groups (all 0.05), while there was no significant difference between the AHSCT NK group and the AHSCT AdIL-2 group ( 0.05). IL-2 secretion in the AHSCT NK AdIL-2 group was significantly higher than that in the AHSCT NK and AHSCT AdIL-2 groups (all 0.05) (Fig. 2). The data indicated that donor NK cells and IL-2 gene modification could enhance the IL-2 secretion in splenocytes of recipient mice.
Donor NK cells and AdIL-2 promote spleen immune reconstitution of recipient mice
In order to observe the changes in spleen immune reconstitution of recipient mice, flow cytometry was conducted to detect the number of immune cells from recipient mice after transplantation. The results revealed that compared with the AHSCT group, the number of B cells, CD T cells, CD T cells, NK cells, and macrophages was significantly elevated in the AHSCT NK, AHSCT AdIL-2, and AHSCT NK AdIL-2 groups (all 0.05), while no significant difference existed between the AHSCT NK group and the AHSCT AdIL-2 group ( 0.05). The number of immune cells in the AHSCT NK AdIL-2 group was significantly higher than that in the AHSCT NK and AHSCT AdIL-2 groups (all 0.05) (Fig. 3A–E). The aforementioned data illustrated the ability of donor NK cells and IL-2 gene modification to promote immune reconstitution of recipient mice.
Donor NK cells and IL-2 gene modification inhibit the deterioration of tissue pathogenesis of recipient mice
HE staining was conducted to for histopathology observation of the receptor mice. Hydropic degeneration of hepatocytes and apparent lymphocytic infiltration in the portal area were observed in the AHSCT group, with sinus expansion and hyperemia in the hepatic vascular, evident edema in the intestinal submucosa, and intestinal structure destruction accompanied by lymphocyte infiltration in the lamina propria also identified. Moreover, in the AHSCT group, vacuolar degeneration in the epidermal layer and focus dissolution in the basal layer were found, with apparent lymphocytic infiltration observed. Compared with the AHSCT group, improved hydropic degeneration of hepatocytes and lymphocyte infiltration, mitigated intestinal structure destruction and lymphocyte infiltration in the lamina propria, as well as reduced vacuolar degeneration in the epidermal layer and focus dissolution in the basal layer were observed in the AHSCT NK, AHSCT AdIL-2, and AHSCT NK AdIL-2 groups, especially in the AHSCT NK AdIL-2 group (Fig. 4). These data elucidated that donor NK cells and IL-2 gene modification could significantly inhibit the deterioration of tissue pathogenesis in recipient mice.
Donor NK cells and IL-2 gene modification increases expression of peripheral blood factors in recipient mice
To further investigate the effect of donor NK cells and IL-2 gene on peripheral blood cytokines in recipient mice after AHSCT, ELISA was performed to detect the expression patterns of TNF-, IFN-, IL-10, TGF- and IL-35. Compared with the AHSCT group, a significant increase was observed in the expression patterns of TNF-, IFN-, IL-10, TGF- and IL-35 in the peripheral blood from the AHSCT NK, AHSCT AdIL-2, and AHSCT NK AdIL-2 groups (all 0.05). The expression patterns of TNF-, IFN-, IL-10, TGF-, and IL-35 in the AHSCT NK AdIL-2 group were significantly higher than the AHSCT NK and AHSCT AdIL-2 groups (all 0.05) (Fig. 5A–E) whereas no significant difference was evident in the cytokine levels between the AHSCT NK group and the AHSCT AdIL-2 group ( 0.05). The aforementioned data suggested that donor NK cells and IL-2 gene modification could increase the expressions of peripheral blood cytokines in recipient mice after transplantation.
Donor NK cells and IL-2 gene modification could significantly increase the expressions of peripheral blood cytokines in recipient mice after transplantation. Panel A–E, expressions of TNF-, IFN-, IL-10, TGF- and IL-35 in peripheral blood are increased in recipient mice treated with donor NK cells and IL-2 gene detected by ELISA; *, 0.05, vs. the AHSCT group; #, 0.05, vs. the AHSCT NK group; &, 0.05, vs. the AHSCT AdIL-2 group; Differences among groups were analyzed by one-way analysis of variance, followed by Tukey’s post hoc test. AHSCT: allogeneic hematopoietic stem cell transplantation; NK: natural killer; AdIL-2, adenovector-mediated delivery of interleukin-2; IL: interleukin; TNF-: tumor necrosis factor-; IFN-: interferon-; TGF-: transforming growth factor ; ELISA: Enzyme-linked immunosorbent assay.
Donor NK cells and IL-2 gene modification promotes immunogenicity in recipient mice
We also determined the fluorescence intensities of leukemic cell-related genes in order to elucidate the possible effect of donor NK cells and IL-2 gene in immunogenicity of leukemic cells in recipient mice. An increase was observed in the fluorescence intensities of leukemic cell surface (MHCII), co-stimulatory molecule antibody (CD40L, CD28), and adherent molecule (ICAM) in the AHSCT NK, AHSCT AdIL-2, and AHSCT NK AdIL-2 groups (all 0.05) when compared to the AHSCT group. No significant difference was found in these fluorescence levels between the AHSCT NK group and the AHSCT AdIL-2 group ( 0.05). In addition, the fluorescence intensities of MHCII, CD40L, CD28 and ICAM in the AHSCT NK AdIL-2 group were notably higher than those in the AHSCT NK and AHSCT AdIL-2 groups (all 0.05, Fig. 6A–D). These results indicated that donor NK cells and IL-2 gene modification could extensively increase the immunogenicity of leukemic cells in recipient mice after transplantation.
Immunogenicity in recipient mice treated with donor NK cells and IL-2 gene is promoted. Panel A-D, fluorescence intensity of MHCII, costimulatory molecule antibody CD40L, CD28 and adherent molecule ICAM in recipient mice after transplantation detected by flow cytometry is increased by IL-2 recombinant adenovirus infected donor NK cells; *, 0.05, vs. the AHSCT group; #, 0.05, vs. AHSCT NK group; &, 0.05, vs. the AHSCT AdIL-2 group; Differences among groups were analyzed by one-way analysis of variance, followed by Tukey’s post hoc test. AHSCT: allogeneic hematopoietic stem cell transplantation; NK: natural killer; AdIL-2, adenovector-mediated delivery of interleukin-2; ICAM: Intercellular adhesion molecule.
Donor NK cells and IL-2 gene modification decreases the relapse rate and mortality rate of recipient mice
The relapse rate and mortality rate of mice in each group were observed from the beginning to the end of the experiment (four months after surgery). The results indicated that compared to the AHSCT group, the relapse rate and mortality rate were significantly decreased in the AHSCT NK, AHSCT AdIL-2, and AHSCT NK AdIL-2 groups, especially in the AHSCT NK AdIL-2 group (all 0.05) (Table 2). These results revealed the ability of donor NK cells and IL-2 gene modification to significantly decrease the relapse rate and mortality rate of recipient mice after transplantation.
The relapse rate and mortality rate of recipient mice treated with donor NK cells and IL-2 gene is decreased
Group
Relapse rate
Mortality rate
AHSCT
93.33%
14/15
Ref.
AHSCT NK
46.67%
7/15
0.014
AHSCT AdIL-2
53.33%
8/15
0.035
AHSCT NK AdIL-2
33.33%
5/15
0.002
Note: The data was expressed as percentage or enumeration data, which were analyzed by chi-square test. Ref., reference; AHSCT, allogeneic Hematopoietic stem cell transplantation; NK, natural killer; AdIL-2, adenovector-mediated delivery of interleukin-2.
Discussion
In recent decades, AHSCT was widely adopted as a relatively effective cellular immunotherapy in treating numerous malignancies, one of which is AL [23]. However, disease relapse has served as a leading source of treatment failure in an inordinate number of patients who underwent AHSCT prior to advancements achieved in the last 2 to 3 decades [24]. A former study flagged the reliability of AHSCT as one of the best therapeutic options for AL [25]. Therefore, tactical strategies should be presented urgently to reduce the recurrent risk in patients [26], upholding us to investigate from a perspective of molecular levels such as the NK cells and IL-2.
Our research revealed that with donor NK cells and IL-2 gene modification, the expression patterns of IL-2 and peripheral blood cytokines (TNF-, IFN-, IL-10, TGF- and IL-35) in the recipient mice exhibited a notable increase as well as the number of B cells, CD T cells, CD T cells, NK cells and macrophages respectively. TNF-, IFN-, IL-10, TGF- and IL-35 fundamentally are anti-inflammatory cytokines [27, 28]. IL-2 initiates specific signaling pathways in the human NK cells facilitating the production of pro- and anti-inflammatory cytokines [29]. Genetic modification incorporates the direct manipulation of an organism’s genome using biotechnology to alter the genetic foundation of cells to facilitate the production of improved or novel organisms [30]. NK cells function in immunity by regulating antigen presenting cells (APCs) and adaptive responses of T cells which are a major producer of IL-2 [11]. The number of total lymphocyte, CD4 T cell, CD8 T cell, B cell is associated with poor transplantation outcomes, and regulatory T cells (Tregs) hold trivial significance in AHSCT as they may help prevent GVHD, low-dose of IL-2 preferentially expands Tregs in vivo, thereby preventing GVHD [31]. On the basis of the interaction between the co-stimulatory molecules expressed on APCs and counter-receptors expressed on T cells, activated T cells induced the growing expression pattern of IL-2 [32], thus demonstrating an increase in IL-2 expressions. The NK cells initiate their functions against infected cells via a complex network of activating and inhibitory signals, which could potentially highlight cytotoxicity and the release of cytokines in the peripheral blood [33], thus prompting a surge in peripheral blood cytokines. In regard to the elevated numbers of B cells, CD T cells, CD T cells, NK cells and macrophages, IL-2 could potentially induce their proliferation and stimulate their antibody-dependent cell-mediated catalytic activity [34, 35]. The proliferation of NK cells by IL-2 has been acknowledged worldwide and has evolved until now by eliminating mesenchymal stem cells, which may otherwise inhibit NK-cell proliferation [36]. Among the peripheral T cells, Treg cells constitute for a large proportion, which are fundamental to immunological tolerance to self Ags [32]. IL2 is the chief cytokine supporting the survival and function of Tregs, and the low-dose of IL2 safely expands Tregs and improves autoimmune conditions in humans [37]. The Treg cells, whose development, expansion, activity, and survival are dependent on IL-2 and which are attributed by the expressions of CD4, CD25, and transcription factor forkhead box P3 (FOXP3), compound for 5 to 10% of circulating CD T cells [17]. Thus, there is a high possibility that speeding IL-2 transcription leads to facilitated recruitment and function of FoxP3 () CD4 () Treg-cell, and then FoxP3 () Treg cells further isolate the CD T cells from the formation of memory-like autoregulatory CD8 () T-cell [38], thereby increasing the numbers. Kohlhapp et al. [39] concluded in their studies that IL-2 exercised an anti-disease property by rapidly increasing number of NK cells, CD T cells and CD T cells, which further supported our results.
Immune reconstitution following conventional allogeneic transplantation is regarded as a predominant determinant of survival [40]. Delayed reconstitution of cellular immunity following CD34-enriched, T-cell-depleted, allogeneic hematopoietic progenitor cell transplantation (HPCT) is the major cause of morbidity and mortality after haploidentical transplantation in adults [41]. The study demonstrated the ability of donor NK cells and IL-2 gene modification to promote stem cell chimerism of AHSCT and immune reconstitution, rescue cells from histopathological deterioration, and intrinsically reduce leukemia relapse. Poor immune reconstitution in human immunodeficiency virus (HIV)-infected patients is evidently associated with an increased proportion of regulatory CD T cells [42]. In addition, human cytomegalovirus infection is the most frequent viral manifestation in patients undergoing HSCT, the control of which requires the presence of immune reconstitution in specific T cells (such as CD and CD T cells) [43]. After anti-thymocyte globulin-conditioned hematopoietic cell transplantation, immune reconstitution is fast for innate immune cells, intermediate for B cells and CD cells, and very slow for CD T cells and invariant NK T cells [44]. Donor NK cells could facilitate the transplantation rate by inhibiting host alloreactive responses during allogeneic bone marrow transplantation (allo-BMT), and donor NK cells and IL-15 treatment could stimulate the transplantation and development of donor-derived cell subsets in a murine nonmyeloablative allo-BMT model [45], which was similar to ours. In light of donor NK cells, they are lymphocytes that are derived from the bone marrow and are significant for the elimination of virally infected and transformed malignant cells [10]. Upon contact with the target cells in the recipient, they hasten their activation threshold, which is particularly evident upon additional activation of the NK receptor ligands in the recipient cells; and once activated, NK cells can radically eliminate leukemic cells in the recipient [46], thereby contributing to cell transplantation rate and immune reconstitution. As previously verified, donor NK cells can facilitate transplantation, impede GVHD, and reduce infections and relapse of leukemia, which is in consistency with our findings [47]. For IL-2, a pleiotropic cytokine produced by T cell after antigen activation is essential for immune response [48]. It is feasible via a connecting link to multimeric receptors that boast the common -chain to stimulate cellular activation, proliferation, and survival of donor cells and recipient cells, thus preventing subsequent cell deterioration and reducing relapse risk [49]. Chen et al. have reported similar results as us in their study ascertaining the ability of donor activated NK cell infusion and IL-2 treatment to assist immune reconstitution, alleviate GVHD and reduce leukemia relapse [50].
In the context of antiretroviral therapy (ART), treatment with IL-2, granulocyte-macrophage colony-stimulating factor (GM-CSF) and recombinant human growth hormone (rhGH) immunotherapy in conjunction with HIV-1 vaccine has shown to promote CD T-cell reconstitution, numbers and function [51]. Extended IL-2 therapy enables persisted clinical and Treg immune responses, and provides durable clinical improvement in active chronic GVHD [52]. Notably, OX40 is an essential factor enhancing Treg sensitivity to IL-2 and triggers their proliferation and survival [53]. Moreover, IL-2 attenuates the severe disruption in T-cell homeostasis induced by HIV infection, through the interaction with T-cells and cytokine microenvironment [54]. miR-625-3p is found upregulated in CD T cells during early immune reconstitution after allogeneic SCT, whose elevated level is counteracted by IL-2 inhibition [55]. Interestingly, many studies have reported that treatment with IL-2 results in a rapid expansion of NK cells [56, 57]. For instance, IL-2 promotes the early recovery of NK cells after HSCT, up-regulates natural cytotoxicity receptors (NCRs) and lytic potential, and thus improves HSCT outcomes [58]. During reconstitution after HSCT, Ly49G2 () NK cells represent the “first-responder” NK cells (the cells display an activated phenotype and are repopulated and highly cytolytic), which occur independently of NK-cell licensing via Ly49-MHC interactions [56]. Post-HSCT GVHD may stem from the reduced NKG2A NK cells induced by IL-10, which further over-activates T cells [59]. The aforementioned suggest the mechanism by which IL-2/NK improve immune reconstitution may involve their interactions with targeted therapies, certain miRNAs, cytokines, and others, which merits further in-depth investigation.
In conclusion, our findings supported that with the help of donor NK cells and IL-2 gene modification, the risk of leukemia relapse after AHSCT could be decreased remarkably, which could be an excitingly new target for leukemia treatment. Additionally, the results in murine studies in the present research may differ in humans, so future studies are needed to verify our results in a human based study. However, more and more prospective studies of larger sample sizes could be further performed for exploring precise estimates to confirm the effects of NK cells in combination with other treatments, to develop new genetic therapeutic strategies for the treatment of AL, and gradually extending for human trials before concluding promising results.
Footnotes
Acknowledgments
We would like to give our sincere appreciation to the reviewers for their helpful comments on this article. This study was supported by the Outstanding Youth Fund in Hebei Province Department of Education (Y2012002) and Natural Science Foundation of Hebei Province (H2012405016).
Conflict of interest
The authors have declared that no competing interests exist.
Abbreviations
References
1.
EscalanteH.J. et al., Acute leukemia classification by ensemble particle swarm model selection, Artif Intell Med55 (2012), 163–175.
2.
XieY. et al., Trends in leukemia incidence and survival in the United States (1973–1998), Cancer97 (2003), 2229–2235.
3.
FaderlS. et al., Clofarabine and cytarabine combination as induction therapy for acute myeloid leukemia (AML) in patients 50 years of age or older, Blood108 (2006), 45–51.
4.
Rodriguez-GarciaJ.A., F., Ramos High incidence of acute leukemia in the proximity of some industrial facilities in El Bierzo, northwestern Spain, Int J Occup Med Environ Health25 (2012).
5.
MatsuokaK. et al., Altered regulatory T cell homeostasis in patients with CD4+ lymphopenia following allogeneic hematopoietic stem cell transplantation, J Clin Invest120 (2010), 1479–1493.
6.
de LimaM. et al., Maintenance therapy with low-dose azacitidine after allogeneic hematopoietic stem cell transplantation for recurrent acute myelogenous leukemia or myelodysplastic syndrome: a dose and schedule finding study, Cancer116 (2010), 5420–5431.
7.
HsiehM.M. et al., Allogeneic hematopoietic stem-cell transplantation for sickle cell disease, N Engl J Med361 (2009), 2309–2317.
8.
StiehlT.BaranN.HoA.D. et al., Clonal selection and therapy resistance in acute leukaemias: mathematical modelling explains different proliferation patterns at diagnosis and relapse, J R Soc Interface11 (2014), 20140079.
9.
NedvetzkiS. et al., Reciprocal regulation of human natural killer cells and macrophages associated with distinct immune synapses, Blood109 (2007), 3776–3785.
10.
VonarbourgC. et al., Regulated expression of nuclear receptor RORgammat confers distinct functional fates to NK cell receptor-expressing RORgammat (+) innate lymphocytes, Immunity33 (2010), 736–751.
11.
LongE.O. et al., Controlling natural killer cell responses: integration of signals for activation and inhibition, Annu Rev Immunol31 (2013), 227–258.
12.
De SantisD. et al., The detection of NK cell alloreactivity by flow cytometric CD107a assay, Methods Mol Biol882 (2012), 477–489.
13.
MullighanC.G. et al., Genome-wide analysis of genetic alterations in acute lymphoblastic leukaemia, Nature446 (2007), 758–764.
14.
LiaoW.LinJ.X.J.W., Leonard Interleukin-2 at the crossroads of effector responses, tolerance, and immunotherapy, Immunity38 (2013), 13–25.
15.
NelsonB.H., IL-2, regulatory T cells, and tolerance, J Immunol172 (2004), 3983–3988.
16.
MalekT.R., The biology of interleukin-2, Annu Rev Immunol26 (2008), 453–479.
17.
KorethJ. et al., Interleukin-2 and regulatory T cells in graft-versus-host disease, N Engl J Med365 (2011), 2055–2066.
18.
MorettaA. et al., Human NK cells: from HLA class I-specific killer Ig-like receptors to the therapy of acute leukemias, Immunol Rev224 (2008), 58–69.
19.
BjorklundA.T. et al., Naive donor NK cell repertoires associated with less leukemia relapse after allogeneic hematopoietic stem cell transplantation, J Immunol196 (2016), 1400–1411.
20.
ZhangW.G. et al., A phase-I clinical trial of active immunotherapy for acute leukemia using inactivated autologous leukemia cells mixed with IL-2, GM-CSF, and IL-6, Leuk Res29 (2005), 3–9.
21.
OrlansF.B., Ethical decision making about animal experiments, Ethics Behav7 (1997), 163–171.
22.
ChenG. et al., Expanded donor natural killer cell and IL-2, IL-15 treatment efficacy in allogeneic hematopoietic stem cell transplantation, Eur J Haematol81 (2008), 226–235.
23.
WelniakL.A. et al., Immunobiology of allogeneic hematopoietic stem cell transplantation, Annu Rev Immunol25 (2007), 139–170.
24.
CairoM.S.JordanC.T.MaleyC.C. et al., NCI first international workshop on the biology, prevention, and treatment of relapse after allogeneic hematopoietic stem cell transplantation: report from the committee on the biological considerations of hematological relapse following allogeneic stem cell transplantation unrelated to graft-versus-tumor effects: state of the science, Biol Blood Marrow Transplant16 (2010), 709–728.
25.
HuangX.J. et al., Treatment of acute leukemia with unmanipulated HLA-mismatched/haploidentical blood and bone marrow transplantation, Biol Blood Marrow Transplant15 (2009), 257–265.
26.
LutzC.MassenkeilG.NagyM. et al., A pilot study of prophylactic donor lymphocyte infusions to prevent relapse in adult acute lymphoblastic leukemias after allogeneic hematopoietic stem cell transplantation, Bone Marrow Transplant41 (2008), 805–812.
27.
MihailovaS. et al., A study of TNF-alpha, TGF-beta, IL-10, IL-6, and IFN-gamma gene polymorphisms in patients with depression, J Neuroimmunol293 (2016), 123–128.
28.
LiX. et al., IL-35 is a novel responsive anti-inflammatory cytokine – a new system of categorizing anti-inflammatory cytokines, PLoS One7 (2012), e33628.
29.
BessolesS. et al., IL-2 triggers specific signaling pathways in human NKT cells leading to the production of pro- and anti-inflammatory cytokines, J Leukoc Biol84 (2008), 224–233.
30.
KernA. et al., Engineering primary metabolic pathways of industrial micro-organisms, J Biotechnol129 (2007), 6–29.
31.
Kennedy-NasserA.A. et al., Ultra low-dose IL-2 for GVHD prophylaxis after allogeneic hematopoietic stem cell transplantation mediates expansion of regulatory T cells without diminishing antiviral and antileukemic activity, Clin Cancer Res20 (2014), 2215–2225.
32.
ElpekK.G.YolcuE.S.FrankeD.D. et al., Ex vivo expansion of CD4+CD25+FoxP3+ T regulatory cells based on synergy between IL-2 and 4-1BB signaling, J Immunol179 (2007), 7295–7304.
33.
MartinJ.F. et al., An IL-2 paradox: blocking CD25 on T cells induces IL-2-driven activation of CD56 (bright) NK cells, J Immunol185 (2010), 1311–1320.
34.
YuA.L. et al., Anti-GD2 antibody with GM-CSF, interleukin-2, and isotretinoin for neuroblastoma, N Engl J Med363 (2010), 1324–1334.
35.
LiaoW., LinX.LeonardW.J., Interleukin-2 at the crossroads of effector responses, tolerance, and immunotherapy, Immunity38 (2013), 13–25.
36.
SpaggiariG.M.CapobiancoA.BecchettiS. et al., Mesenchymal stem cell-natural killer cell interactions: evidence that activated NK cells are capable of killing MSCs, whereas MSCs can inhibit IL-2-induced NK-cell proliferation, Blood107 (2006), 1484–1490.
37.
ChurlaudG. et al., Sustained stimulation and expansion of Tregs by IL2 control autoimmunity without impairing immune responses to infection, vaccination and cancer, Clin Immunol151 (2014), 114–126.
38.
ShameliA.YamanouchiJ.TsaiS. et al., IL-2 promotes the function of memory-like autoregulatory CD8+ T cells but suppresses their development via FoxP3+ Treg cells, Eur J Immunol43 (2013), 394–403.
39.
KohlhappF.J. et al., NK cells and CD8+ T cells cooperate to improve therapeutic responses in melanoma treated with interleukin-2 (IL-2) and CTLA-4 blockade, J Immunother Cancer3 (2015), 18.
40.
D’SaS. et al., T- and B-cell immune reconstitution and clinical outcome in patients with multiple myeloma receiving T-cell-depleted, reduced-intensity allogeneic stem cell transplantation with an alemtuzumab-containing conditioning regimen followed by escalated donor lymphocyte infusions, Br J Haematol123 (2003), 309–322.
41.
WallerE.K. et al., Facilitating T-cell immune reconstitution after haploidentical transplantation in adults, Blood Cells Mol Dis33 (2004), 233–237.
42.
HortaA. et al., Poor immune reconstitution in HIV-infected patients associates with high percentage of regulatory CD4+ T cells, PLoS One8 (2013), e57336.
43.
LilleriD. et al., Human cytomegalovirus-specific CD4+ and CD8+ T-cell reconstitution in adult allogeneic hematopoietic stem cell transplant recipients and immune control of viral infection, Haematologica93 (2008), 248–256.
44.
BoschM. et al., Immune reconstitution after anti-thymocyte globulin-conditioned hematopoietic cell transplantation, Cytotherapy14 (2012), 1258–1275.
45.
HuB. et al., Donor NK clls and IL-15 promoted engraftment in nonmyeloablative allogeneic bone marrow transplantation, J Immunol189 (2012), 1661–1670.
46.
DecotV. et al., Natural-killer cell amplification for adoptive leukemia relapse immunotherapy: comparison of three cytokines, IL-2, IL-15, or IL-7 and impact on NKG2D, KIR2DL1, and KIR2DL2 expression, Exp Hematol38 (2010), 351–362.
47.
Perez-MartinezA. et al., Blood dendritic cells suppress NK cell function and increase the risk of leukemia relapse after hematopoietic cell transplantation, Biol Blood Marrow Transplant17 (2011), 598–607.
48.
HuX. et al., A novel modulation of structural and functional changes of mouse bone marrow derived dendritic cells (BMDCs) by interleukin-2 (IL-2), Hum Vaccin Immunother11 (2015), 516–521.
49.
GeddesM. et al., Immune reconstitution following hematopoietic stem-cell transplantation, Best Pract Res Clin Haematol20 (2007), 329–348.
50.
ChenG.H. et al., Experimental study on IL-2- and IL-15 application in allogeneic hematopoietic stem cell transplantation, Zhonghua Xue Ye Xue Za Zhi29 (2008), 526–530.
51.
CockerA.T.H. et al., Short communication: therapeutic immunization benefits mucosal-associated invariant t cell recovery in contrast to interleukin-2, granulocyte-macrophage colony-stimulating factor, and recombinant human growth hormone addition in HIV-1+ treated patients: individual case reports from phase i trial, AIDS Res Hum Retroviruses35 (2019), 306–309.
52.
KorethJ. et al., Efficacy, durability, and response predictors of low-dose interleukin-2 therapy for chronic graft-versus-host disease, Blood128 (2016), 130–137.
53.
PiconeseS. et al., A non-redundant role for OX40 in the competitive fitness of Treg in response to IL-2, Eur J Immunol40 (2010), 2902–2913.
54.
TincatiC.d’Arminio MonforteA. and MarchettiG., Immunological mechanisms of interleukin-2 (IL-2) treatment in HIV/AIDS disease, Curr Mol Pharmacol2 (2009), 40–45.
55.
VermaK. et al., miR-625-3p is upregulated in CD8+ T cells during early immune reconstitution after allogeneic stem cell transplantation, PLoS One12 (2017), e0183828.
56.
BaraoI. et al., Mouse Ly49G2+ NK cells dominate early responses during both immune reconstitution and activation independently of MHC, Blood117 (2011), 7032–7041.
57.
TomchuckS.L.LeungW.H. and DallasM.H., Enhanced cytotoxic function of natural killer and CD3+CD56+ cells in cord blood after culture, Biol Blood Marrow Transplant21 (2015), 39–49.
58.
PittariG. et al., Early evaluation of natural killer activity in post-transplant acute myeloid leukemia patients, Bone Marrow Transplant45 (2010), 862–871.
59.
HuL.J. et al., Quantity and quality reconstitution of NKG2A (+) natural killer cells are associated with graft-versus-host disease after allogeneic hematopoietic cell transplantation, Biol Blood Marrow Transplant25 (2019), 1–11.