Besides the crucial role of hyperinsulinemia in the development of breast cancer with Type 2 diabetes mellitus (T2DM), it has been shown that hyperglycemia could contribute to promote cancer progression. A remarkable association within hyperglycemia, PKC and Ubiquitin-proteasome system (UPS) has been reported, suggesting that PKC may mediate high glucose-induced UPS activation in breast cancer cells. Although the independent effects of PKC or UPS on breast cancer and T2DM are increasingly supported by experimental evidence, the complex interactional link between PKC and UPS is still unclear. Hence, we focus on the relationship between PKC and UPS in breast cancer with T2DM. We hypothesize that PKC may have the function to regulate the activity of UPS. Further, we speculate that PKC combine with proteasome 2 promoter, that indicate PKC regulate the function of UPS by change the composition of proteasome. Therefore, we surmise that PKC mediated high glucose-induced UPS activation in breast cancer cells, and specific PKC inhibitor rottlerin significantly suppressed elevated glucose induced the activity of UPS. We hope that our paper will stimulate further studies the relationship between PKC and UPS, and a new targeted therapy and early medical intervention for PKC could be a useful option for breast cancer cases complicated with T2DM or hyperglycemia.
Breast cancer is the most commonly cancer for women worldwide and it is the underlying cause of cancer-related death in women [1]. Type 2 diabetes mellitus (T2DM) is characterized by a chronic hyperglycemic state concurrent with insulin resistance and/or impaired insulin secretion [2]. It now demonstrated that T2DM is related to increase breast cancer incidence and mortality [3, 4, 5, 6, 7]. Indeed, prediabetes may also increase the risk of breast cancer [5, 8, 9, 10, 11]. The detailed mechanisms remain unknown; however, hyperglycemia in T2DM patients may play important roles in the process of breast cancer.
In this paper, we hypothesize that PKC may have the function to regulate the activity of UPS. Further, PKC combine with proteasome 2 promoter, that indicate PKC regulate the function of UPS by change the composition of proteasome. Therefore, PKC regulated high glucose-induced UPS activation in breast cancer cells, and rottlerin, specific PKC inhibitor, dramatically suppressed high glucose-induced the activation of UPS.
The Protein kinase C family
The Protein kinase C (PKC) pathways represent one of the most familiar mechanisms by a signal relay of extracellular response as a phospholipid or calcium-dependent protein kinase [12]. Presence of a cell and tissue changes in the distribution and abundance, which is divided into three sub-categories represent 10 different mammalian PKC isoforms: the “classical” (PKC, I, II, ), which may be calcium and diacylglycerol stimulation (DAG) or phorbol esters; “novel” (PKC, , , ), which can be activated by DAG, but independent of calcium or phorbol ester; and the ‘atypical’ (protein kinase C and /), which is in response to calcium, DAG and phorbol esters. PKC isoforms are crucial for differentiation, proliferation and apoptosis, but more limited portion of the key [13, 14]. Because of its contribution to intracellular signal transduction, it makes sense to suppose that the alterative expression or activity levels of these kinases may result in diseases, including breast cancer. One of the PKCs identified to exhibit equivocal properties is PKC [15]. Most researches reached consensus on the PKC as a tumor suppressing activity having an anti-proliferative in different cells [15]. While several reports indicated PKC was identified as the pro-proliferative role and a cancer-promoting gene [16].
Role of PKC in breast cancer
The effects of PKC in breast cancer are still vague, and the expression levels of PKC in primary breast cancer are inadequate. Though it was not a prerequisite for the cancer progression, some researches have confirmed that overexpressing PKC is able to be a tumor promotion in breast cancer cells [17, 18]. PKC is the most important partner in Ras/Raf/MEK/ERK pathway, that has been proved to accelerate cellular proliferation and tumorigenesis [19]. Insulin growth factor-I (IGF-I) signaling is another primary signaling in regulating growth. Interplays between PKC and mTOR have demonstrated for modulating stress and IGF-I targeted the Insulin receptor substrate 1 (IRS-1) phosphorylation in MCF-7 breast cancer cells [20]. On the other hand, some literatures have confirmed that PKC was response to anti-proliferative effect. For instance, the inositol hexaphosphate possessed the antimitogenic effect in MCF-7 by PKC-induced phosphorylation inhibition of ERK, AKT and Rb [21].
PKC also plays an ambiguous role in cell cycle modulation and apoptosis. It has revealed that PKC plays a proapoptotic role in MCF-7 for UV damage by a caspase dependent mechanism [22]. This PKC-activated mechanism results in the phosphorylation of aSMase, leading to apoptosis ceramide generation and signal enhancement [23]. In contrast, PKC has anti-apoptotic effect in MDA-MB-231and MCF-7 for ionizing radiation [24] and tumor necrosis factor-related apoptosis-inducing ligand (TRAIL)-induced apoptosis [25]. In addition, Grossini et al. [26] has shown that PKC promotes proliferation in murine mammary cells. PKC regulates the G1/S phase transition to influence cell cycle. PKC mediates p21-dependent pathway in SKBR-3 and p27-dependent pathway in MCF-7 resulting in G1 arrest [21, 27, 28]. Both the p21 and p27 can combine with cyclin E/Cdk2 complex and inhibit its activity.
To meet invasion and metastasis of tumor, they must be capable of progression. PKC has been demonstrated to inhibit the MCF-7migration [29]. This reduction may be partly due to the regulation of PKC. However, for breast cancer cells PKC has been shown to activate MMP-9 [30, 31], again increasing the ambiguity of PKC effects on breast cancer progress.
Numerous studies show that the crosstalk between PKC and Estrogen receptor (ER). Inhibition of PKC blocks most of the 17-B-estriadiol-regulated but not affects TNF- activated ERK1/2 in MCF-7 [32]. Markov et al. [33] have reported that the methylation of PKC gene in ER+ breast cancer is correlated with a favorable response to anti-estrogenic therapy. However, another study [34] found that raised PKC protein is related to the sensitivity of endocrine therapy. In addition, tamoxifen-resistant breast cancer cells usually express higher PKC protein than their parental tamoxifen-sensitive cells [34, 35]. Regarding a potential role for PKC as a target for breast cancer therapy, AD198, a new doxorubicin analogues lacking the ability to bind and inhibit DNA topoisomerase II, induces apoptosis by activating PKC [36].
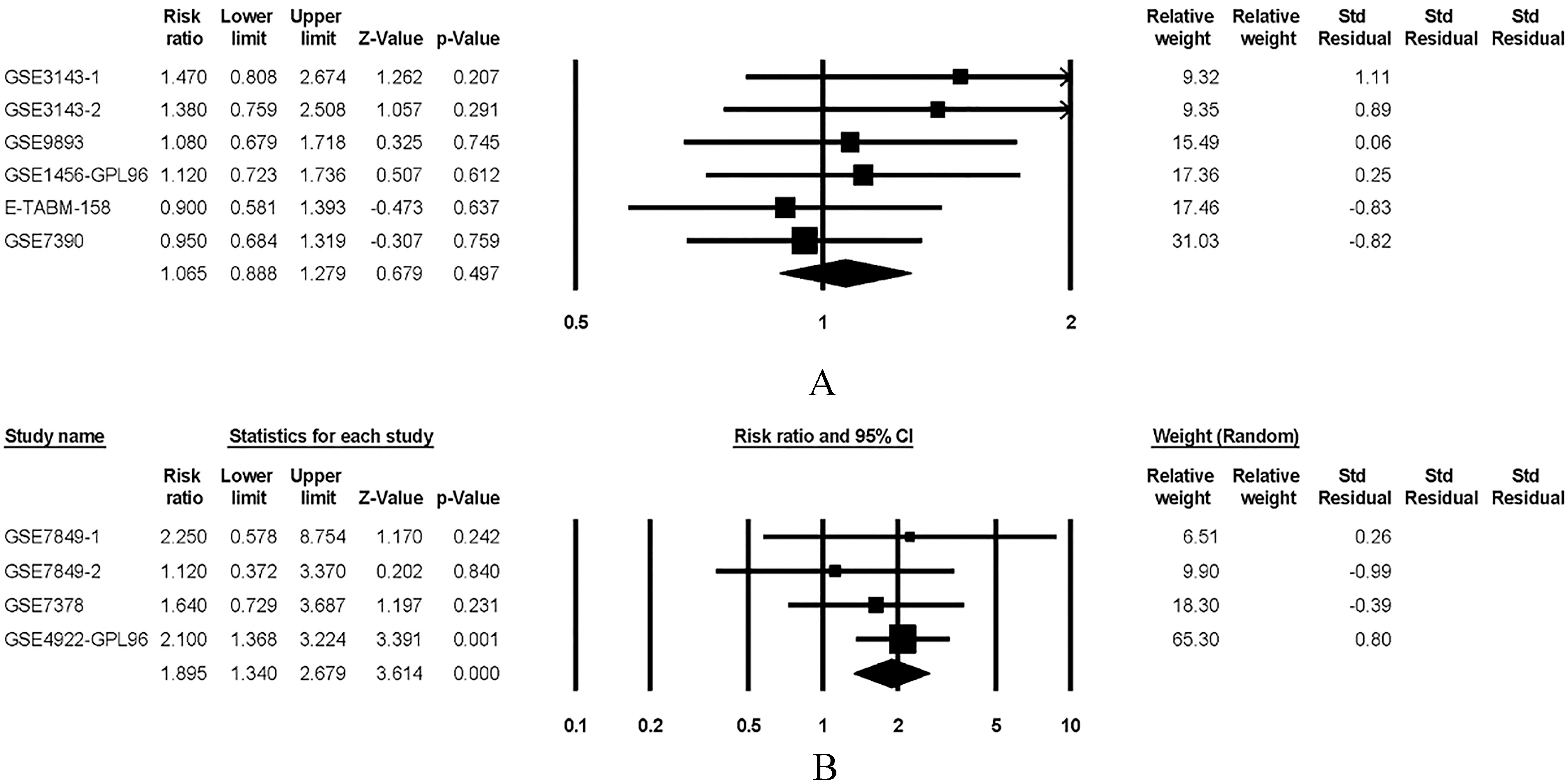
PrognoScan is a meta-analysis database served as the prognostic value of genes [37]. PrognoScan is publicly acquired at http://gibk21.bse.kyutech.ac.jp/PrognoScan/index.html. The meta-analysis was carried out using the Comprehensive meta-analysis system version 2 (Comprehensive Meta-Analysis; Biostat Inc, Englewood, NJ). The results clearly indicate that PKC expression was non-significant trend toward worse OS, but PKC overexpression was significantly associated with a lower rate of DFS, as shown in Fig. 1 (OS, RR 1.065, P 0.497; DFS, RR 1.895, P 0.001). These finding further confirm that PKC was potential biomarkers and associated with poor prognosis of breast cancer.
The expressions of PKC correlate with overall survival (OS) and disease free survival (DFS)in breast cancer patients. (A). The forest plot curves showed correlation between OS and PKC expression levels in breast cancer patients. (B). The forest plot curves showed correlation between DFS and PKC expression levels in breast cancer patients.
Role of PKC in diabetes
Type 2 diabetes mellitus (T2DM) and obesity are main global health problem, and the incidence is expected to increase further in the coming two decades. Genome-wide scanning found out the difference in the development of diabetes by several Quantitative trait locus (QTLs) [38, 39]. The chromosome 14 is the strongest QTL and is related to the occurrence of hyperinsulinemia and insulin resistance. Gene screening associated with insulin resistance in a region is PRKCD encoding PKC. PKC played an important effect in insulin resistance by fatty acid and lipogenesis in liver [40, 41], PKC has also primary modulation effect of insulin resistance [42]. PKC regulates muscle glucose uptake [43], lipogenesis [41], endoplasmic reticulum (ER) stress [44], and angiopoiesis [45]. PKC also affects insulin sensitivity by regulating inflammation. For instance, PKC acts as the regulator in response to TNF--induced the activation of NF-B and ER stress in liver [44]. PKC has also resulted in ROS accumulation of adipocytes in High fat diet (HFD)-treated animals, it could aggravate insulin resistance and increase risk of T2DM [46, 47]. PKC is also resulted in inflammation of adipose tissue, such as PKC activation has leaded to the secretion and production of IL-6 in mesenteric adipose [48]. Therefore, PKC represents a strongly potential therapeutic targets for insulin resistance, T2DM as well as related complications.
The ubiquitin proteasome system and breast cancer
The Ubiquitin-proteasome system (UPS) facilitates the degradation of proteins and regulation of growth and stress response and is required for many biological processes such as efficient cell cycle control, stress response, DNA repair, and differentiation [49]. The UPS consists of a variety of crucial enzymes including Ebiquitin-activating enzyme (E1), Ebiquitin-conjugating enzyme (E2), Ebiquitin ligase (E3), the 26S proteasome and the Deubiquitinating enzymes (DUBs) [50, 51]. Normally, E1 activates ubiquitin in an ATP-dependent manner. The activated ubiquitin is then transferred from an E1 enzyme to an E2 enzyme. E3 transfers the activated ubiquitin to lysine residues of targeted proteins, forming a polyubiquitin chain that earmarks the targeted proteins. The specificity of the E3 enzymes determines the specific recognition of target proteins, providing selectivity in which proteins are targeted to the proteasome for degradation [52]. DUBs are a cluster of enzymes that oppose the action of the E3 ligases by cleaving the isopeptide bonds between lysine residues of targeted proteins and the C-terminal glycine of ubiquitin [53, 54].
Tumorigenesis is due to the malfunction in cell growth, proliferation and apoptosis [55]. UPS regulates protein degradation that is involved in cell proliferation, apoptosis, migration, and invasion. Disruption of the ubiquitination and deubiquitination by mutations or modified expression of specific components within the cascade plays an important role in tumorigenesis. Recently, various studies have been conducted to clarify and highlight the roles of UPS in breast cancer.
E2 enzymes and breast cancer
Various E2 enzymes such as Ubiquitin-conjugating enzyme 9 (UBC 9), UBC13, UBE2C, E2D3, E2Q1, and E2S have been shown to be involved in the development and progression of breast cancer. UBC9 has been frequently reported to be over-expressed in breast cancer which is positively related with poor breast cancer specific survival [56, 57]. Chen et al. also demonstrated that high Ubc9 expression was associated with poor response to chemotherapy and poor clinical prognosis [58]. UBC13 expression is elevated in primary and metastatic breast cancer and could promote metastatic spread of breast cancer [59]. UBE2C contributes to ubiquitin-mediated proteasome degradation of cell cycle progression in breast cancer [60]. Knockdown or inhibition of UBE2C could reduce proliferation and sensitizes breast cancer cells to radiation, doxorubicin, tamoxifen, and letrozole [61, 62]. UBE2D3 participates in the ubiquitination and degradation of cyclin D1 which is stabilized after UBE2D3 suppression and promotes proliferation of breast cancer cells in vitro [63]. UBE2Q1 could mediate ubiquitination and subsequent proteasomal degradation of p53 [64]. UBE2S is associated with malignant characteristics of breast cancer cells such as migration and invasion [65]. These findings strongly suggest that E2 enzymes are implicated in the development and metastasis of breast cancer.
E3 enzymes and breast cancer
There are various E3 enzymes participating in the development of breast cancer, some of which are involved in cancer progression and metastasis. RNF31 highly expressed in breast cancer facilitates p53 polyubiquitination and degradation and promotes cancer progression [66, 67]. The Carboxyl terminus of the Hsc70-interacting protein (CHIP) could result in downregulation of profilin1 and enhanced cell migration [68]. However, another study focusing on ER-positive breast cancer patients has demonstrated that the overexpression of CHIP is a potent prognostic factor of a good prognosis [69]. Further study is needed to clarify this issue. RNF115 (also known as BCA2) is also reported to be overexpressed in ER-positive breast tumors and to promote breast cell proliferation through downregulating the expression of p21 [70]. Besides acting as cancer promoters, some E3 enzymes act as tumor suppressors in breast cancer. HRD1 could suppress the growth and metastasis of breast cancer cells by promoting IGF-1R degradation and attenuated IL-6-induced epithelial-mesenchymal transition [71]. Smurf2 could regulate cell polarity, migration, division, differentiation, and death, by targeting diverse substrates that are critical for receptor signaling, cytoskeleton, chromatin remodeling, and transcription. Recently, Liu et al. found that Smurf2 might serve as a tumor suppressor in triple negative breast cancer [72]. E6AP, acting both as an E3 ubiquitin-protein ligase and as a coactivator of steroid hormone receptors could suppress breast cancer invasiveness, colonization, and metastasis by regulating actin cytoskeletal remodeling via regulation of Rho GTPases [73]. There are still many E3 enzymes playing important roles in breast cancer development, progression and metastasis. We thus summarized several recently reported E3 enzymes in Table 1.
DUBs play important roles in ubiquitin processing, reversal of ubiquitin signaling and recycling of ubiquitin. There are about 100 DUBs in human body categorized into six classes: Ubiquitin-specific proteases (USPs), ubiquitin carboxy-terminal hydrolases (UCHs), Ovarian-tumor proteases (OTUs), Machado-Joseph disease protein domain proteases, JAMM/MPN domain-associated metallopeptidases (JAMMs) and Monocyte chemotactic protein-induced protein (MCPIP) family [83, 84]. These DUBs also play crucial roles in breast cancer. ATXN3L, a member of Machado-Joseph disease protein domain proteases, could promote breast cancer proliferation by deubiquitinating Krüppel-like factor 5 [85]. USP2 which belongs to USPs was overexpressed in triple negative breast cancer and associated with increased cell migration and invasiveness [86]. However, some DUBs play a controversial role in breast cancer. For example, Li et al. found that the USP4 could inhibit breast cancer cell growth while other studies demonstrated USP4 could stimulate the TGF-mediated EMT, invasion and metastasis [87].
PKC and the Ubiquitin-proteasome system
The remaining question is whether and how does PKC regulate the UPS? Some studies have suggested that PKC and PKC activation regulates transcriptional activity and degradation of progesterone receptor (PR) by the 26S proteasome [88]. As previously mentioned, our study suggests that the expressions of PKC and its phosphorylation are dramatically increased in breast cancer tissues with diabetes when compared with breast cancer alone. While proteasome chymotrypsin-like activity is also markedly increased, and there is a high degree of correlation between the expression of PKC and chymotrypsin-like protease activities. As a consequence, we speculate that PKC may have the function to regulate the activity of UPS. Further, we have demonstrated that PKC mediates high glucose-induced UPS activation, and specific PKC inhibitor rottlerin significantly suppresses high glucose induced the activity of UPS. This presents a very interesting experimental model through which the interaction of PKC and UPS could be investigated by ChIP assay [17]. In MCF-7 cells, PKC combines with proteasome 2 promoter, which indicate that PKC regulates the function of UPS by alter the composition of proteasome.
Cytoscape is an open source software project for integrating bimolecular interaction networks with high-throughput expression data and other molecular states into a unified conceptual framework [89]. The results distinctly indicate that c-Myc interacts with PKC and proteasome 2 promoter for glycometabolism pathways. The results present that PKC connected with c-Myc translation and proteasome 2 promoter has an affinity with c-Myc. c-Myc is a well-known oncogene frequently up-regulated in different malignancies, that modulated glucose metabolism in various carcinoma [90, 91, 92]. Consequently, we guess that the combination of PKC with proteasome 2 promoter may affect the activation of c-Myc, further modulated glucose metabolism to promote tumor progression.
PKC is thought to be an ambiguous role in carcinogenesis. Furthermore, numerous in vitro and in vivo studies have suggested that PKC have been linked to increased invasion, proliferation, drug resistance and genetic instability. UPS regulates degradation of proteins that is involved in cell proliferation, apoptosis, migration, and invasion. Disruption of UPS by mutations or modified expression of specific components within the cascade plays an important role in tumorigenesis. The notion supports a possibility of that PKC regulates the function of UPS by changing the composition of proteasome in the aggressiveness of a cancer. Human high-grade cancers very frequently display mutations in genes such as c-Myc, which lead to increased proliferation rate, metastasis, anti-apoptosis, and resistance to therapy. Altered PKC and imbalanced UPS may be a mediator of these pro-cancer events. Taken together, PKC regulates the function of UPS during breast cancer progression, and targeted therapies against PKC could be effective in the treatment cancers.
In summary, our hypothesis points to the role of PKC in the regulation of UPS in a subset of breast cancer patients with T2DM or hyperglycemia. The presence of PKC overexpression in breast cancer cases complicated with T2DM or hyperglycemia could worsen survival and contribute to cancer progression.
Footnotes
Acknowledgments
This work was partially supported by a National Natural Science Foundation of China (NSFC) grant to Dr. Shengrong Sun (Grant NO: 81471781) and the Fundamental Research Funds for the Central Universities grant to Dr. Si Sun (Grant NO: 2042015kf0111).
Conflict of interest
No other conflict of interests to declare.
References
1.
SiegelR.L.MillerK.D. and JemalA., Cancer statistics, 2016, CA Cancer J Clin66 (2016), 7–30.
2.
American DiabetesA., Diagnosis and classification of diabetes mellitus, Diabetes Care37(Supple 1) (2014), S81–90.
3.
WolfI.SadetzkiS.CataneR.KarasikA. and KaufmanB., Diabetes mellitus and breast cancer, Lancet Oncol6 (2005), 103–111.
4.
PanB.RenH.HeY.LvX.MaY.LiJ.HuangL.YuB.KongJ.NiuC.ZhangY.SunW.B. and ZhengL., HDL of patients with type 2 diabetes mellitus elevates the capability of promoting breast cancer metastasis, Clin Cancer Res18 (2012), 1246–1256.
5.
PeairsK.S.BaroneB.B.SnyderC.F.YehH.C.SteinK.B.DerrR.L.BrancatiF.L. and WolffA.C., Diabetes mellitus and breast cancer outcomes: a systematic review and meta-analysis, J Clin Oncol29 (2011), 40–46.
6.
EricksonK.PattersonR.E.FlattS.W.NatarajanL.ParkerB.A.HeathD.D.LaughlinG.A.SaquibN.RockC.L. and PierceJ.P., Clinically defined type 2 diabetes mellitus and prognosis in early-stage breast cancer, J Clin Oncol29 (2011), 54–60.
7.
LipscombeL.L.GoodwinP.J.ZinmanB.McLaughlinJ.R. and HuxJ.E., Diabetes mellitus and breast cancer: a retrospective population-based cohort study, Breast Cancer Res Treat98 (2006), 349–356.
8.
De BruijnK.M.ArendsL.R.HansenB.E.LeeflangS.RuiterR. and van EijckC.H., Systematic review and meta-analysis of the association between diabetes mellitus and incidence and mortality in breast and colorectal cancer, Br J Surg100 (2013), 1421–1429.
9.
KaplanM.A.PekkolayZ.KucukonerM.InalA.UrakciZ.ErtugrulH.AkdoganR.FiratU.YildizI. and IsikdoganA., Type 2 diabetes mellitus and prognosis in early stage breast cancer women, Med Oncol29 (2012), 1576–1580.
10.
Salinas-MartinezA.M.Flores-CortesL.I.Cardona-ChavarriaJ.M.Hernandez-GutierrezB.AbundisA.Vazquez-LaraJ. and Gonzalez-GuajardoE.E., Prediabetes, diabetes, and risk of breast cancer: a case-control study, Arch Med Res45 (2014), 432–438.
11.
OnitiloA.A.StankowskiR.V.BergR.L.EngelJ.M.GlurichI.WilliamsG.M. and DoiS.A., Breast cancer incidence before and after diagnosis of type 2 diabetes mellitus in women: increased risk in the prediabetes phase, Eur J Cancer Prev23 (2014), 76–83.
12.
TakaiY.KishimotoA.IwasaY.KawaharaY.MoriT. and NishizukaY., Calcium-dependent activation of a multifunctional protein kinase by membrane phospholipids, J Biol Chem254 (1979), 3692–3695.
13.
MusashiM.OtaS. and ShiroshitaN., The role of protein kinase C isoforms in cell proliferation and apoptosis, Int J Hematol72 (2000), 12–19.
14.
LiD.LiC.WuM.ChenQ.WangQ.RenJ. and ZhangY., PKCdelta stabilizes TAp63 to promote cell apoptosis, FEBS Lett590 (2016), 1672.
15.
JacksonD.N. and FosterD.A., The enigmatic protein kinase Cdelta: complex roles in cell proliferation and survival, FASEB J18 (2004), 627–636.
16.
KileyS.C.ClarkK.J.DuddyS.K.WelchD.R. and JakenS., Increased protein kinase C delta in mammary tumor cells: relationship to transformtion and metastatic progression, Oncogene18 (1999), 6748–6757.
17.
ZhuS.YaoF.LiW.-H.WanJ.-N.ZhangY.-M.TangZ.KhanS.WangC.-H. and SunS.-R., PKCδ-dependent Activation of the Ubiquitin Proteasome System is Responsible for High Glucose-induced Human Breast Cancer MCF-7 Cell Proliferation, Migration and Invasion, Asian Pacific Journal of Cancer Prevention14 (2013), 5687–5692.
18.
SunS.LiuY.LuJ.OmarA.SunS.BiY. and WangC., The inhibitory effects of PKCtheta on adiponectin expression is mediated by ERK in 3T3-L1 adipocytes, J Endocrinol Invest34 (2011), 8–15.
19.
UedaY.HiraiS.OsadaS.SuzukiA.MizunoK. and OhnoS., Protein kinase C activates the MEK-ERK pathway in a manner independent of Ras and dependent on Raf, J Biol Chem271 (1996), 23512–23519.
20.
Mingo-SionA.M.FergusonH.A.KollerE.ReylandM.E. and Van Den BergC.L., PKCdelta and mTOR interact to regulate stress and IGF-I induced IRS-1 Ser312 phosphorylation in breast cancer cells, Breast Cancer Res Treat91 (2005), 259–269.
21.
VucenikI.RamakrishnaG.TantivejkulK.AndersonL.M. and RamljakD., Inositol hexaphosphate (IP6) blocks proliferation of human breast cancer cells through a PKCdelta-dependent increase in p27Kip1 and decrease in retinoblastoma protein (pRb) phosphorylation, Breast Cancer Res Treat91 (2005), 35–45.
22.
DenningM.F.WangY.NickoloffB.J. and Wrone-SmithT., Protein kinase Cdelta is activated by caspase-dependent proteolysis during ultraviolet radiation-induced apoptosis of human keratinocytes, J Biol Chem273 (1998), 29995–30002.
23.
ZeidanY.H.WuB.X.JenkinsR.W.ObeidL.M. and HannunY.A., A novel role for protein kinase Cdelta-mediated phosphorylation of acid sphingomyelinase in UV light-induced mitochondrial injury, FASEB J22 (2008), 183–193.
24.
McCrackenM.A.MiragliaL.J.McKayR.A. and StroblJ.S., Protein kinase C delta is a prosurvival factor in human breast tumor cell lines, Mol Cancer Ther2 (2003), 273–281.
25.
YinS.SethiS. and ReddyK.B., Protein kinase Cdelta and caspase-3 modulate TRAIL-induced apoptosis in breast tumor cells, J Cell Biochem111 (2010), 979–987.
26.
GrossoniV.C.FalboK.B.KazanietzM.G.de Kier JoffeE.D. and UrtregerA.J., Protein kinase C delta enhances proliferation and survival of murine mammary cells, Mol Carcinog46 (2007), 381–390.
27.
YokoyamaG.FujiiT.TayamaK.YamanaH.KuwanoM. and ShirouzuK., PKCdelta and MAPK mediate G(1) arrest induced by PMA in SKBR-3 breast cancer cells, Biochem Biophys Res Commun327 (2005), 720–726.
28.
ShanmugamM.KrettN.L.MaizelsE.T.MuradF.M.RosenS.T. and Hunzicker-DunnM., A role for protein kinase C delta in the differential sensitivity of MCF-7 and MDA-MB 231 human breast cancer cells to phorbol ester-induced growth arrest and p21(WAFI/CIP1) induction, Cancer Lett172 (2001), 43–53.
29.
JacksonD.ZhengY.LyoD.ShenY.NakayamaK.NakayamaK.I.HumphriesM.J.ReylandM.E. and FosterD.A., Suppression of cell migration by protein kinase Cdelta, Oncogene24 (2005), 3067–3072.
30.
LinC.W.HouW.C.ShenS.C.JuanS.H.KoC.H.WangL.M. and ChenY.C., Quercetin inhibition of tumor invasion via suppressing PKC delta/ERK/AP-1-dependent matrix metalloproteinase-9 activation in breast carcinoma cells, Carcinogenesis29 (2008), 1807–1815.
31.
Alonso-EscolanoD.MedinaC.CieslikK.RadomskiA.JuraszP.Santos-MartinezM.J.JiffarT.RuvoloP. and RadomskiM.W., Protein kinase C delta mediates platelet-induced breast cancer cell invasion, J Pharmacol Exp Ther318 (2006), 373–380.
32.
KeshamouniV.G.MattinglyR.R. and ReddyK.B., Mechanism of 17-beta-estradiol-induced Erk1/2 activation in breast cancer cells. A role for HER2 AND PKC-delta, J Biol Chem277 (2002), 22558–22565.
33.
MartensJ.W.NimmrichI.KoenigT.LookM.P.HarbeckN.ModelF.KluthA.Bolt-de VriesJ.SieuwertsA.M.PortengenH.Meijer-Van GelderM.E.PiepenbrockC.OlekA.HoflerH.KiechleM.KlijnJ.G.SchmittM.MaierS. and FoekensJ.A., Association of DNA methylation of phosphoserine aminotransferase with response to endocrine therapy in patients with recurrent breast cancer, Cancer Res65 (2005), 4101–4117.
34.
AssenderJ.W.GeeJ.M.LewisI.EllisI.O.RobertsonJ.F. and NicholsonR.I., Protein kinase C isoform expression as a predictor of disease outcome on endocrine therapy in breast cancer, J Clin Pathol60 (2007), 1216–1221.
35.
NabhaS.M.GlarosS.HongM.LykkesfeldtA.E.SchiffR.OsborneK. and ReddyK.B., Upregulation of PKC-delta contributes to antiestrogen resistance in mammary tumor cells, Oncogene24 (2005), 3166–3176.
36.
Diaz BessoneM.I.BerardiD.E.CampodonicoP.B.TodaroL.B.LothsteinL.Bal de Kier JoffeE.D. and UrtregerA.J., Involvement of PKC delta (PKCdelta) in the resistance against different doxorubicin analogs, Breast Cancer Res Treat126 (2011), 577–587.
37.
MizunoH.KitadaK.NakaiK. and SaraiA., PrognoScan: a new database for meta-analysis of the prognostic value of genes, BMC Med Genomics2 (2009), 18.
38.
AlmindK. and KahnC.R., Genetic determinants of energy expenditure and insulin resistance in diet-induced obesity in mice, Diabetes53 (2004), 3274–3285.
39.
AlmindK.KulkarniR.N.LannonS.M. and KahnC.R., Identification of interactive loci linked to insulin and leptin in mice with genetic insulin resistance, Diabetes52 (2003), 1535–1543.
40.
LamT.K.YoshiiH.HaberC.A.BogdanovicE.LamL.FantusI.G. and GiaccaA., Free fatty acid-induced hepatic insulin resistance: a potential role for protein kinase C-delta, Am J Physiol Endocrinol Metab283 (2002), E682–91.
41.
FrangioudakisG.BurchfieldJ.G.NarasimhanS.CooneyG.J.LeitgesM.BidenT.J. and Schmitz-PeifferC., Diverse roles for protein kinase C delta and protein kinase C epsilon in the generation of high-fat-diet-induced glucose intolerance in mice: regulation of lipogenesis by protein kinase C delta, Diabetologia52 (2009), 2616–2620.
42.
BezyO.TranT.T.PihlajamakiJ.SuzukiR.EmanuelliB.WinnayJ.MoriM.A.HaasJ.BiddingerS.B.LeitgesM.GoldfineA.B.PattiM.E.KingG.L. and KahnC.R., PKCdelta regulates hepatic insulin sensitivity and hepatosteatosis in mice and humans, J Clin Invest121 (2011), 2504–2517.
43.
JacobA.I.Horovitz-FriedM.Aga-MizrachiS.Brutman-BarazaniT.OkhrimenkoH.ZickY.BrodieC. and SampsonS.R., The regulatory domain of protein kinase C delta positively regulates insulin receptor signaling, J Mol Endocrinol44 (2010), 155–169.
44.
GreeneM.W.RuhoffM.S.BurringtonC.M.GarofaloR.S. and OrenaS.J., TNFalpha activation of PKCdelta, mediated by NFkappaB and ER stress, cross-talks with the insulin signaling cascade, Cell Signal22 (2010), 274–284.
45.
GeraldesP.Hiraoka-YamamotoJ.MatsumotoM.ClermontA.LeitgesM.MaretteA.AielloL.P.KernT.S. and KingG.L., Activation of PKC-delta and SHP-1 by hyperglycemia causes vascular cell apoptosis and diabetic retinopathy, Nat Med15 (2009), 1298–1306.
46.
TaliorI.YarkoniM.BashanN. and Eldar-FinkelmanH., Increased glucose uptake promotes oxidative stress and PKC-delta activation in adipocytes of obese, insulin-resistant mice, Am J Physiol Endocrinol Metab285 (2003), E295–302.
47.
TaliorI.TennenbaumT.KurokiT. and Eldar-FinkelmanH., PKC-delta-dependent activation of oxidative stress in adipocytes of obese and insulin-resistant mice: role for NADPH oxidase, Am J Physiol Endocrinol Metab288 (2005), E405–11.
48.
WallerstedtE.SmithU. and AnderssonC.X., Protein kinase C-delta is involved in the inflammatory effect of IL-6 in mouse adipose cells, Diabetologia53 (2010), 946–954.
49.
DeshaiesR.J., Proteotoxic crisis, the ubiquitin-proteasome system, and cancer therapy, BMC Biol12 (2014), 94.
50.
HershkoA.HellerH.EliasS. and CiechanoverA., Components of ubiquitin-protein ligase system. Resolution, affinity purification, and role in protein breakdown, J Biol Chem258 (1983), 8206–8214.
51.
HochstrasserM., Ubiquitin-dependent protein degradation, Annu Rev Genet30 (1996), 405–439.
52.
KonstantinovaI.M.TsimokhaA.S. and MittenbergA.G., Role of proteasomes in cellular regulation, Int Rev Cell Mol Biol267 (2008), 59–124.
53.
FraileJ.M.QuesadaV.RodriguezD.FreijeJ.M. and Lopez-OtinC., Deubiquitinases in cancer: new functions and therapeutic options, Oncogene31 (2012), 2373–2388.
54.
TsouW.L.SheedloM.J.MorrowM.E.BlountJ.R.McGregorK.M.DasC. and TodiS.V., Systematic analysis of the physiological importance of deubiquitinating enzymes, PLoS One7 (2012), e43112.
55.
HanahanD. and WeinbergR.A., The hallmarks of cancer, Cell100 (2000), 57–70.
56.
AgboolaA.O.MusaA.A.AyoadeB.A.BanjoA.A.AnunobiC.C.Deji-AgboolaA.M.RakhaE.A.NolanC.EllisI.O. and GreenA.R., Clinicopathological and molecular significance of Sumolyation marker (ubiquitin conjugating enzyme 9 (UBC9)) expression in breast cancer of black women, Pathol Res Pract210 (2014), 10–17.
57.
WozniakK.KrupaR.SynowiecE. and MorawiecZ., Polymorphism of UBC9 gene encoding the SUMO-E2-conjugating enzyme and breast cancer risk, Pathol Oncol Res20 (2014), 67–72.
58.
ChenS.F.GongC.LuoM.YaoH.R.ZengY.J. and SuF.X., Ubc9 expression predicts chemoresistance in breast cancer, Chin J Cancer30 (2011), 638–644.
59.
WuX.ZhangW.Font-BurgadaJ.PalmerT.HamilA.S.BiswasS.K.PoidingerM.BorcherdingN.XieQ.ElliesL.G.LytleN.K.WuL.W.FoxR.G.YangJ.DowdyS.F.ReyaT. and KarinM., Ubiquitin-conjugating enzyme Ubc13 controls breast cancer metastasis through a TAK1-p38 MAP kinase cascade, Proc Natl Acad Sci U S A111 (2014), 13870–13875.
60.
ChouC.P.HuangN.C.JhuangS.J.PanH.B.PengN.J.ChengJ.T.ChenC.F.ChenJ.J. and ChangT.H., Ubiquitin-conjugating enzyme UBE2C is highly expressed in breast microcalcification lesions, PLoS One9 (2014), e93934.
61.
RawatA.GopalG.SelvaluxmyG. and RajkumarT., Inhibition of ubiquitin conjugating enzyme UBE2C reduces proliferation and sensitizes breast cancer cells to radiation, doxorubicin, tamoxifen and letrozole, Cell Oncol (Dordr)36 (2013), 459–467.
62.
WangC.PanY.H.ShanM.XuM.BaoJ.L. and ZhaoL.M., Knockdown of UbcH10 enhances the chemosensitivity of dual drug resistant breast cancer cells to epirubicin and docetaxel, Int J Mol Sci16 (2015), 4698–4712.
63.
WangW.YangL.HuL.LiF.RenL.YuH.LiuY.XiaL.LeiH.LiaoZ.ZhouF.XieC. and ZhouY., Inhibition of UBE2D3 expression attenuates radiosensitivity of MCF-7 human breast cancer cells by increasing hTERT expression and activity, PLoS One8 (2013), e64660.
64.
ShafieeS.M.RastiM.SeghatoleslamA.AzimiT. and OwjiA.A., UBE2Q1 in a Human Breast Carcinoma Cell Line: Overexpression and Interaction with p53, Asian Pac J Cancer Prev16 (2015), 3723–3727.
65.
AkterK.A.HyodoT.AsanoE.SatoN.MansourM.A.ItoS.HamaguchiM. and SengaT., Erratum to: UBE2S is associated with malignant characteristics of breast cancer cells, Tumour Biol37 (2016), 6999.
66.
ZhuJ.ZhaoC.ZhuangT.JonssonP.SinhaI.WilliamsC.StrombladS. and Dahlman-WrightK., RING finger protein 31 promotes p53 degradation in breast cancer cells, Oncogene35 (2016), 1955–1964.
67.
ZhuJ.ZhuangT.YangH.LiX.LiuH. and WangH., Atypical ubiquitin ligase RNF31: the nuclear factor modulator in breast cancer progression, BMC Cancer16 (2016), 538.
68.
ChoiY.N.LeeS.K.SeoT.W.LeeJ.S. and YooS.J., C-Terminus of Hsc70-interacting protein regulates profilin1 and breast cancer cell migration, Biochem Biophys Res Commun446 (2014), 1060–1066.
69.
KurozumiS.YamaguchiY.HayashiS.HiyoshiH.SudaT.GohnoT.MatsumotoH.TakeiH.HoriguchiJ.TakeyoshiI.OyamaT. and KurosumiM., Prognostic value of the ubiquitin ligase carboxyl terminus of the Hsc70-interacting protein in postmenopausal breast cancer, Cancer Med5 (2016), 1873–1882.
70.
WangZ.NieZ.ChenW.ZhouZ.KongQ.SethA.K.LiuR. and ChenC., RNF115/BCA2 E3 ubiquitin ligase promotes breast cancer cell proliferation through targeting p21Waf1/Cip1 for ubiquitin-mediated degradation, Neoplasia15 (2013), 1028–1035.
71.
XuY.M.WangH.J.ChenF.GuoW.H.WangY.Y.LiH.Y.TangJ.H.DingY.ShenY.C.LiM.XuanW.Y.LiuL.H.WangJ.WangX.R.GaoZ.J.LiangX.B. and SuD.M., HRD1 suppresses the growth and metastasis of breast cancer cells by promoting IGF-1R degradation, Oncotarget6 (2015), 42854–42867.
72.
LiuX.GuX.SunL.FlowersA.B.RademakerA.W.ZhouY. and KiyokawaH., Downregulation of Smurf2, a tumor-suppressive ubiquitin ligase, in triple-negative breast cancers: involvement of the RB-microRNA axis, BMC Cancer14 (2014), 57.
73.
MansourM.HauptS.ChanA.L.GoddeN.RizzitelliA.LoiS.CaramiaF.DebS.TakanoE.A.BishtonM.JohnstoneC.MonahanB.Levav-CohenY.JiangY.H.YapA.S.FoxS.BernardO.AndersonR. and HauptY., The E3-ligase E6AP Represses Breast Cancer Metastasis via Regulation of ECT2-Rho Signaling, Cancer Res (2016).
74.
ZhangL.TengY.FanY.WangY.LiW.ShiJ.MaY.LiC.ShiX.QuX. and LiuY., The E3 ubiquitin ligase Cbl-b improves the prognosis of RANK positive breast cancer patients by inhibiting RANKL-induced cell migration and metastasis, Oncotarget6 (2015), 22918–22933.
75.
ChenA.WongC.S.LiuM.C.HouseC.M.SceneayJ.BowtellD.D.ThompsonE.W. and MollerA., The ubiquitin ligase Siah is a novel regulator of Zeb1 in breast cancer, Oncotarget6 (2015), 862–873.
76.
DuanY.HuoD.GaoJ.WuH.YeZ.LiuZ.ZhangK.ShanL.ZhouX.WangY.SuD.DingX.ShiL.WangY.ShangY. and XuanC., Ubiquitin ligase RNF20/40 facilitates spindle assembly and promotes breast carcinogenesis through stabilizing motor protein Eg5, Nat Commun7 (2016), 12648.
77.
ZhouX.DengS.LiuH.LiuY.YangZ.XingT.JingB. and ZhangX., Knockdown of ubiquitin protein ligase E3A affects proliferation and invasion, and induces apoptosis of breast cancer cells through regulation of annexin A2, Mol Med Rep12 (2015), 1107–1113.
78.
YangS.ChenY.SunF.NiQ.WangH.HuangY.ZhangC.LiuK.WangS.QiuJ.XuZ.HuaL. and HeZ., Downregulated PIRH2 Can Decrease the Proliferation of Breast Cancer Cells, Arch Med Res47 (2016), 186–195.
79.
HakemA.BohgakiM.LemmersB.TaiE.SalmenaL.Matysiak-ZablockiE.JungY.S.KaraskovaJ.KaustovL.DuanS.MadoreJ.BoutrosP.ShengY.ChesiM.BergsagelP.L.Perez-OrdonezB.Mes-MassonA.M.PennL.SquireJ.ChenX.JurisicaI.ArrowsmithC.SanchezO.BenchimolS. and HakemR., Role of Pirh2 in mediating the regulation of p53 and c-Myc, PLoS Genet7 (2011), e1002360.
80.
GuoH.WuF.WangY.YanC. and SuW., Overexpressed ubiquitin ligase Cullin7 in breast cancer promotes cell proliferation and invasion via down-regulating p53, Biochem Biophys Res Commun450 (2014), 1370–1376.
81.
McDonaldW.J.ThomasL.N.KoiralaS. and TooC.K., Progestin-inducible EDD E3 ubiquitin ligase binds to alpha4 phosphoprotein to regulate ubiquitination and degradation of protein phosphatase PP2Ac, Mol Cell Endocrinol382 (2014), 254–261.
82.
GokaE.T. and LippmanM.E., Loss of the E3 ubiquitin ligase HACE1 results in enhanced Rac1 signaling contributing to breast cancer progression, Oncogene34 (2015), 5395–5405.
83.
PalA. and DonatoN.J., Ubiquitin-specific proteases as therapeutic targets for the treatment of breast cancer, Breast Cancer Res16 (2014), 461.
84.
NijmanS.M.Luna-VargasM.P.VeldsA.BrummelkampT.R.DiracA.M.SixmaT.K. and BernardsR., A genomic and functional inventory of deubiquitinating enzymes, Cell123 (2005), 773–786.
85.
GeF.ChenW.QinJ.ZhouZ.LiuR.LiuL.TanJ.ZouT.LiH.RenG. and ChenC., Ataxin-3 like (ATXN3L), a member of the Josephin family of deubiquitinating enzymes, promotes breast cancer proliferation by deubiquitinating Kruppel-like factor 5 (KLF5), Oncotarget6 (2015), 21369–21378.
86.
QuQ.MaoY.XiaoG.FeiX.WangJ.ZhangY.LiuJ.ChengG.ChenX.WangJ. and ShenK., USP2 promotes cell migration and invasion in triple negative breast cancer cell lines, Tumour Biol36 (2015), 5415–5423.
87.
LiY.JiangD.ZhangQ.LiuX. and CaiZ., Ubiquitin-specific protease 4 inhibits breast cancer cell growth through the upregulation of PDCD4, Int J Mol Med38 (2016), 803–811.
88.
Gonzalez-ArenasA.Pena-OrtizM.A.Hansberg-PastorV.Marquina-SanchezB.Baranda-AvilaN.Nava-CastroK.Cabrera-WroomanA.Gonzalez-JorgeJ. and Camacho-ArroyoI., PKCalpha and PKCdelta activation regulates transcriptional activity and degradation of progesterone receptor in human astrocytoma cells, Endocrinology156 (2015), 1010–1022.
89.
ShannonP.MarkielA.OzierO.BaligaN.S.WangJ.T.RamageD.AminN.SchwikowskiB. and IdekerT., Cytoscape: a software environment for integrated models of biomolecular interaction networks, Genome Res13 (2003), 2498–2504.
90.
HuangJ.KongW.ZhangJ.ChenY.XueW.LiuD. and HuangY., c-Myc modulates glucose metabolism via regulation of miR-184/PKM2 pathway in clear-cell renal cell carcinoma, Int J Oncol (2016).
91.
DangC.V., MYC on the path to cancer, Cell149 (2012), 22–35.
92.
WahlstromT. and HenrikssonM.A., Impact of MYC in regulation of tumor cell metabolism, Biochim Biophys Acta1849 (2015), 563–569.