
Abstract
Preliminary observations were conducted on the nonparasitic phase of the life cicle of
The PAL did not change withing different grasses and it varied from 27 to 32 days according the month of exposure. The grass plot method was the best.
The SL varied according the grass and month of exposure, being on dry and hot months shorter in
The LT was short enough in the dry months to lead to recomendations on the use of pasture spelling methods for tick control, but precise information is needed. More research is necesary on quantitative methods before their use on field work.
Introduccion
Por su amplia distribución en las regiones tropicales y subtropicales, la garrapata del ganado
Los expertos en control de garrapatas enfatizan la necesidad de los estudios sobre el ciclo no parasítico de la garrapata, y la influencia de los factores ambientales en él, con el fin de poder conocer y preveer las fluctuaciones en cuanto a abundancia de garrapatas en un ecotipo particular, así como incorporar en los estudios de la epidemiología de enfermedades transmitidas por garrapatas para usar esta información en el establecimiento de planes racionales de control. El ciclo no parasítico de
Las observaciones descritas a continuación fueron realizadas en el Centro Nacional de Investigaciones Agropecuarias "Carimagua", localizado en el extremo nororiental del Departamento del Meta en los Llanos Orientales de Colombia. El Centro es representativo de cuatro millones de hectáreas de la altillanura plana colombiana.
Revision de Literatura
La importancia del estudio del ciclo no parasítico de la garrapata ha sido ampliamente discutida por numerosos autores. Hitchcock (1955) anota que la gran longevidad potencial de la larva indica la extrema importancia de la consideración de este estado en el establecimiento de medidas prácticas de control; pero Wilkinson (1961) expone cómo la amplia distribución de las larvas hace impracticable, económicamente, controlar infestaciones mediante el tratamiento de las praderas con acaricidas.
Harley (1966) sostiene que los datos de supervivencia en el campo son esenciales para la aplicación más efectiva de métodos de control como el descanso de potreros y los baños estratégicos; asimismo expone que el período crítico a considerar en la planeación de la rotación de potreros es el transcurrido desde la caída de la teleogina a la muerte de su progenie, período conocido como longevidad máxima. En este sentido cita los experimentos de Harley y Wilkinson en 1964, con descanso de potreros un mes menor que la longevidad máxima total obtenida para garrapatas expuestas en períodos precedentes.
Las fases no parasíticas de
El mismo autor demostró que el período de preoviposición varió de 19-30 días a 15°C a 2-3 días a 36°C; que la duración de la oviposición no fue influenciada por la humedad relativa pero varió de un máximo de 44 días a 15°C a un mínimo de 14 días a 36°C.
Harley (1966) encontró que el período preeclosión (preoviposición + oviposición + incubación) está correlacionado con la temperatura del suelo durante el período de exposición; también encontró que éste período es considerablemente más largo en invierno que en verano; la baja temperatura retarda la oviposición y eclosión de los huevos.
La humedad relativa, la temperatura, el período adulto-larva o preeclosión y la factibilidad de acceso al rocío parecen influir en la supervivencia larvaria, la cual se ha encontrado es más corta para la progenie de garrapatas expuestas en la estación seca y más larga hacia el fin de la estación húmeda. La disminución de la humedad relativa durante los meses secos parece ser la causante de este efecto (Harley, 1966; Hitchcock, 1955).
Hay evidencias de que la supervivencia larvaria depende en alguna extensión de la temperatura y humedad relativa a las que fue sometido el estado de huevo. Hay una aparente relación inversa entre la duración del período preeclosión y la supervivencia larvaria, relación no aparente en las épocas secas del año (Harley, 1966). Se ha obtenido un máximo de supervivencia de 240 días bajo condiciones de laboratorio a 22°C y 90% de humedad relativa (Hitchcock, 1955). Las larvas pueden recuperar pérdidas de agua producidas por bajas humedades relativas, mediante absorción de la atmósfera durante subsecuentes períodos de alta humedad relativa. Asimismo, la larva puede beber agua cuando la encuentra libre, principalmente en forma de rocío (Wilkingson y Wilson, 1959; Hitchcock, 1955; Harley, 1966). De otro lado, se ha encontrado alguna evidencia de que la atmósfera saturada no es buena para la larva porque pronto se vuelve letárgica y sobrevive períodos más cortos que a humedades relativas ligeramente menores. Las larvas son más vulnerables que los huevos a bajas temperaturas. (Hitchcock, 1955).
La longevidad larvaria es influenciada por el grado de actividad. Hitchcock (1955) encontró que cultivos frecuentemente estimulados viven períodos más cortos que los controles en similares condiciones. Los vientos y lluvias fuertes pueden desplazar las larvas de su posición de reposo en el pasto y, aunque no las destruyan realmente, pueden acortar sus vidas por crearles un gasto de energía al tratar éstas de volver a ganar posiciones favorables en la pradera (Wilkinson y Wilson, 1959).
Una circunstancia del patrón de supervivencia larvaria en el campo es la tendencia de la mayoría de las larvas de persistir por un período mucho más corto que un pequeño porcentaje de sus congéneres, existiendo poco conocimiento sobre aspectos cuantitativos de la supervivencia larvaria (Snowball, 1957; Wilkinson y Wilson, 1959).
Materiales Y Metodos
Colonia De Laboratorio
Las teologinas utilizadas durante todas las observaciones fueron obtenidas de la colonia de laboratorio establecida en "Carimagua" y mantenida sobre dos terneros de seis y ocho meses de edad, los que permanecían en aislamiento en una caseta especialmente diseñada y eran infestados regularmente con cultivos de larvas mantenidas en el laboratorio.
Praderas
Se trabajó con las siguientes gramíneas:
Para cada pradera se sembraron dos materas de 2 m2 de área, limitada por un círculo de zinc.
Observaciones En Tubos De Malla
Estas observaciones se iniciaron en mayo de 1981, realizando dos observaciones por semana, y finalizaron en marzo de 1982. Se utilizaron tubos de malla de cobre (15 agujeros por cm2) de un tamaño aproximado de 5 cms. por 1,5 de diámetro, los cuales eran sellados debidamente para evitar el escape de la garrapata o de su progenie.
Inicialmente, en el período comprendido entre mayo y octubre, se colocaron dos teleoginas por tubo y dos tubos por gramínea, en una matera de 2 m2 de área. Las teleoginas de uno de los tubos se pesaron en una balanza de precisión (d= 0,01).
En estos tubos se determinó:
Período adulto-larva (PAL) que es el tiempo entre la caída de la garrapata del animal y la aparición de su progenie (larvas).
Indice de potencial reproductivo (IPR), medida cuantitativa desarrollada en este trabajo con el ánimo de conocer las variaciones en cantidad de progenie producida por una teleogina de peso conocido en distintos hábitats. El IPR se determinó recolectando los tubos cuyas teleoginas se habían pesado, el día 38 post-exposición y contando el número de larvas producido, previa inmovilización por refrigeración; el número de larvas encontradas se dividió por la suma del peso de las teleoginas (IPR = No. de larvas en tubo/peso tel A + tel B en mg).
Total de tubos produciendo progenie sobre el total expuesto, lo que arbitrariamente se llamó natalidad y se expresó en fracciones (natalidad = tubos con progenie/total tubos); la natalidad está influenciada por la fecundidad o capacidad de la hembra para ovipositar y por la eclosionabilidad de los huevos (Thompson, 1976).
En las observaciones de noviembre a diciembre de 1981, se aumentó a seis el número de tubos colocados por matera en cada exposición, pesando las teleoginas en dos de estos tubos, así se determinó:
PAL (6 tubos)
IPR (2 tubos)
Natalidad (6 tubos).
Supervivencia larvaria (SL) que es el tiempo máximo que sobreviven las larvas sin estar adheridas a ningún hospedante, determinándose en la última observación donde se hallaron larvas con movimiento (4 tubos) (SL No. días entre eclosión y última observación con movimiento).
Para las observaciones de enero y febrero, se colocaron dos tubos adicionales pesando las teleoginas con el fin de determinar el Indice de Eficiencia de la Conversión (IEC), parámetro introducido por Drummond y Whestone en 1970 (Davey et al, 1980), para determinar la conversión del peso de una hembra ingurgitada en huevos; para ello el día 150 de colocados, se colocaron dos tubos y se pesaron los huevos producidos (IEC = Peso huevos producidos/Peso tel A + tel B).
Observaciones En Materas
En las materas de 2 m2 para cada gramínea, se inicó una observación en julio 1981, sembrando en cada matera 30 teleoginas obtenidas de la colonia; las materas eran podadas cada ocho días para simular pastoreo, las hojas con larvas adheridas eran repuestas a la matera. En ellas se determinó el PAL hasta el día de aparición de larvas libres en las hojas y la SL hasta la desaparición de las mismas (dos observaciones por semana). En noviembre 81 y enero 1982 se mejoró la tecnología de muestreo adaptándola a la usada por Harley (1966), sembrando 30 teleoginas por matera, que al momento de la siembra había sido podada a una altura de 10 cms. y posteriormente no fue tocada. El PAL se determinó tan pronto se observaron larvas libres en las hojas. Para determinar la SL, se continuó la observación dos veces por semana hasta no encontrar larvas visualmente; entonces se realizó un barrido de la matera con un palo de 30 cms. de largo por 5 cms. de diámetro, recubierto con bayetilla blanca. Las larvas así recuperadas se regresaban a la pradera por medio de un pincel. La inspección continuó hasta que en seis barridas en el día de la inspección no se recuperaron larvas luego de cuatro observaciones consecutivas. La supervivencia se consideró hasta la primera de estas observaciones en que no se recuperaron larvas en el barrido.
Registros Meteorologicos
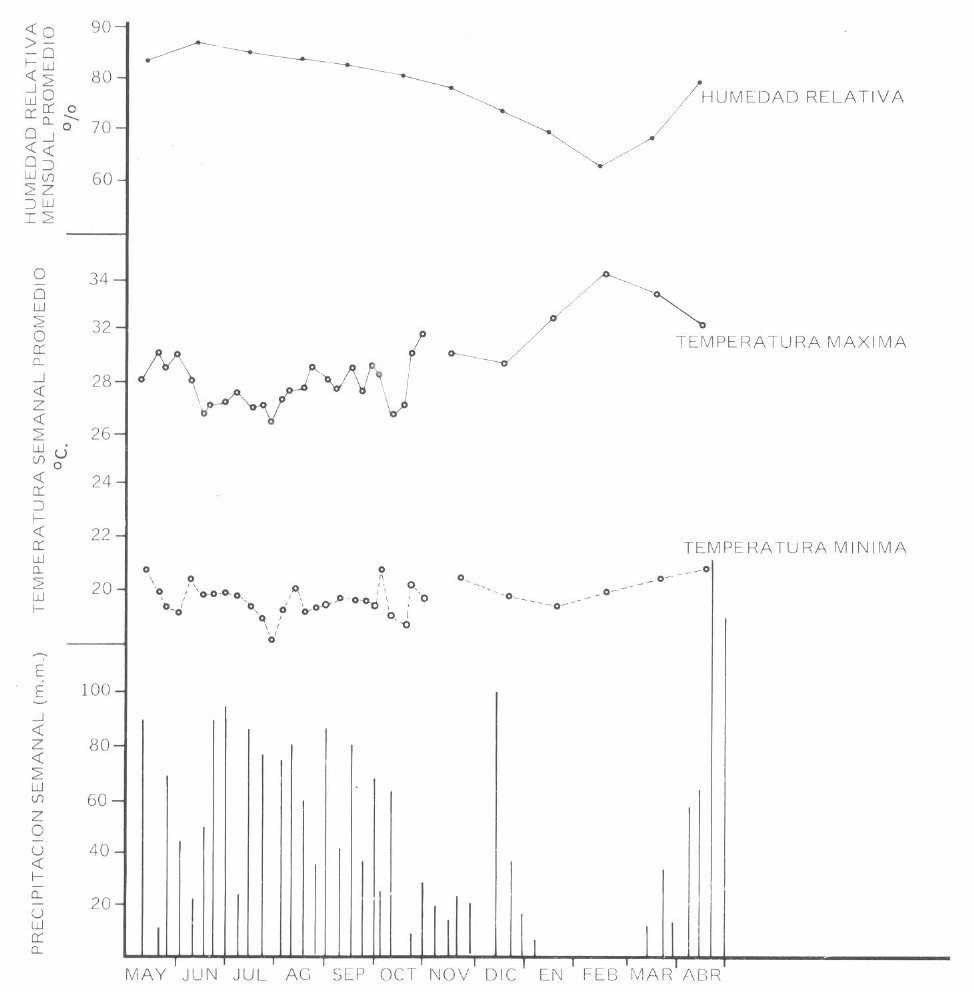
Los registros de precipitación pluvial se obtuvieron de la estación del HIMAT del Centro, durante todo el período de observaciones. Los registros de temperatura máxima y mínima fueron tomados de la misma fuente hasta octubre de 1981 agrupándose por semanas. Por carecer de datos, las gráficas de noviembre a abril se calcularon con base en los promedios mensuales obtenidos en el período 1971-1980. La humedad relativa fue calculada de la misma manera.
Analisis Estadisticos
Los valores de PAL y SL se registraron como los valores mínimos y máximos respectivamente encontrados en cada exposición. Para el análisis de IPR se realizó el análisis de varianza usando un modelo de bloques al azar considerando cada fecha de exposición como bloque y cada gramínea utilizada como tratamiento. El mismo modelo fue usado para el análisis de IEC.
Resultados Y Discusion
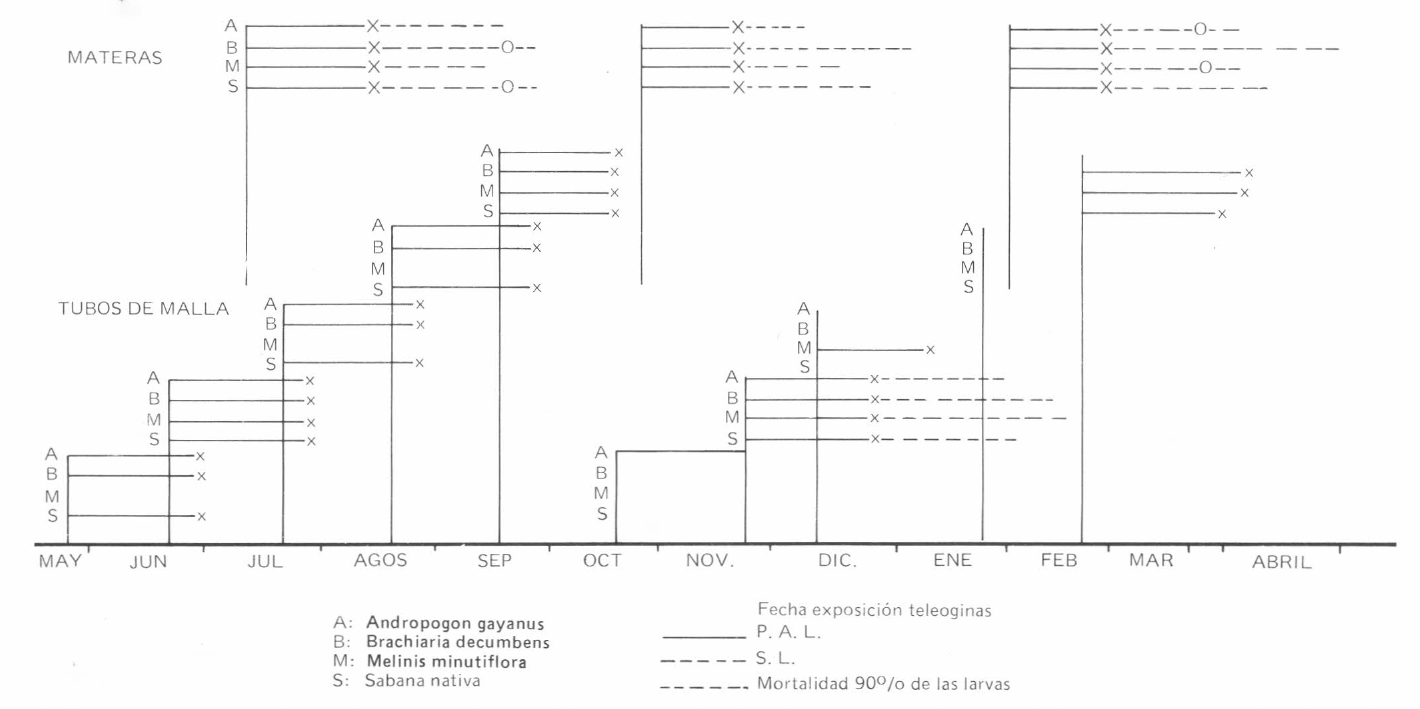
En la Tabla 1 se presentan los resultados obtenidos para los distintos parámetros obtenidos en las exposiciones realizadas en tubos de malla. Los resultados de las observaciones en materas en resumen en la Tabla 2; el PAL determinado por la obervación de larvas libres en las hojas no demostró variaciones entre las gramíneas, pero sí ligeras diferencias en épocas siendo 32 días en julio, 27 días en octubre y 29 en febrero.
Duración del ciclo no parasítico y resultados de parámetros cuantitativos de
P.A.L. Período Adulto Larva;
S.L. Supervivencia larvaria;
I.E.C. Peso huevo/peso teleogina;
I.P.R. No. larvas/peso teleogina.
A
B
M
S Sabana nativa
N.D. No determinado.
Duración del ciclo no parasítico de
Días en los que se notó una disminución del 90% de larvas dificultando su observación.
P.A.L. Período Adulto Larva;
S.L. Supervivencia Larvaria;
L.T. Longevidad Total;
A.
B
M
S Sabana.
En la gráfica 1 se observan los resultados de meteorología durante el período de observación. La precipitación expresada como totales por semana demuestra un período seco con ausencia total de lluvias abarcando los meses de enero y febrero, que coincide con la máxima temperatura promedio (34°C) y una baja humedad relativa (62%). Adicionalmente se realizó la medición de la humedad relativa directamente en las materas a 30 cms. del suelo utilizando un higrómetro MASON, en la primera semana de marzo cuando aún no habían iniciado las lluvias, en la segunda semana del mes luego de una lluvia de 15 mm., dos días con anterioridad a la evaluación y la cuarta semana del mes luego de varias lluvias que sumaron 64,7 mm. de precipitación. Los resultados de cada matera se expresan en la Tabla 3.
Evaluación del ciclo no parasítico de
Evaluación ciclo no parasítico de
Período adulto larva (P.A.L.) y supervivencia larvaria (S. L.) en
Discusion
Periodo Adulto Larva (Pal)
Los resultados sobre el PAL obtenidos en estas observaciones (28-41 días en tubos de malla, 27-32 días en materas) concuerdan con los obtenidos por Mateus (Mateus V., G. Estudio sobre el ciclo de vida del
Al aumentar la temperatura disminuye el período de desarrollo (Hitchcock, 1955; Snowball, 1957). Las temperaturas promedias para "Carimagua" son: julio 24,5°C y febrero 28,5°C. El PAL más prolongado en la exposición de febrero puede ser debido a un período de preadaptación larvaria más prolongado por condiciones adversas de humedad más que a un retardo en la eclosión de los huevos.
Un PAL de 41 días fue observado en los tubos expuestos en verano en
El PAL observado en cilindros fue mayor que el encontrado en las exposiciones en el mismo mes en la matera; esto coincide con las observaciones de Snowball (1957) y es causado por la desventaja del confinamiento en los tubos, pero esto no se considera mayor problema en este tipo de estudios.
Estudios sobre biología de la oviposición de
La Supervivencia Larvaria
La SL en tubos de malla sólo fue posible determinarla en una ocasión, presentando períodos relativamente cortos para todas las gramíneas observadas, efecto causado por las condiciones ambientales adversas en el período de vida larvaria (enero-febrero) con humedades relativas promedio de 74 y 70% respectivamente (Gráficas 1 y 2).
Para la exposición realizada en julio, la SL en materas presentó una duración de 41 días en
Para la evaluación de SL en materas en las exposiciones de octubre y febrero, se siguió una metodología similar a la descrita por Harley (1966); Snowball (1957) y Wilkinson y Wilson (1959).
La corta SL observada en la exposición de octubre se explica por las condiciones ambientales cálidas y secas que encontraban las larvas desde su nacimiento (Harley, 1966; Wilkinson y Wilson, 1959), principalmente en
Las larvas resultantes de las teleoginas expuestas en febrero presentan SL ligeramente mayores, explicadas por el aumento de la humedad relativa luego de las primeras lluvias caídas en marzo. La Tabla 3, explica claramente este efecto. En la primera semana del mes, la humedad relativa en las materas promediaba 32% mientras que en la última semana promedió 91,5%. Cómo las larvas sobreviven a humedades ambientales tan bajas, es explicado por los hallazgos de Wilkinson y Wilson (1959) que indican que la larva puede recuperar pérdidas de humedad absorbiéndola de la atmósfera o bebiendo rocío en las horas de la noche cuando la humedad relativa aumenta.
Longevidad Total
La LT en observaciones en tubos de malla y materas se presenta en la Gráfica 2. Los valores obtenidos para la exposición en materas del mes de julio no se consideran representativos. Los resultados de LT en la observación en tubos de malla para la exposición de noviembre (
De Jesús (1934) encontró que el efecto repelente del pasto gordura (
La corta LT obtenida en las observaciones de octubre y febrero, en todas las gramíneas, indica la factibilidad del uso del descanso de potreros como medida de control de garrapatas en la zona, en la época seca. Snowball (1957) indica que los grupos de larvas disminuyeron rápidamente en las hojas hasta quedar muy pocas que permanecen largos períodos, Harley (1966) cita experimentos que presentan períodos de descanso un mes menores que la longevidad máxima obtenida para garrapatas expuestas el mes precedente. El autor hace énfasis en que períodos de descenso más largos darían mejor control pero podrían causar problemas por fiebre de garraptas al alterar la estabilidad enzoótica. De esta manera, los períodos de descanso a utilizar en las diferentes praderas serían 15-20 días menores a la longevidad total (LT) obtenida en estas observaciones.
Metodos Cuantitativos
Los métodos cuantitativos como el IPR y el IEC se han desarrollado con el fin de conocer el potencial de fertilidad de la garrapata y poder usar estos conocimientos integrados con planes de control.
El IEC promedio de todas las observaciones (0,327) es menor al reportado en la región en condiciones de medio ambiente y que fue de 0,485 (Benavides, 1982). Con el fin de tener un patrón de comparación se realizó conjuntamente con la exposición de enero la determinación del IEC en teleoginas de la colonia en condiciones ambientales lo que promedió 0,474 (Tabla 4).
Indice de eficiencia de la conversión "IEC". Teleoginas. Colonia de laboratorio de
Promedios de 29 observaciones.
Este menor IEC se explica por la manipulación a la que se someten las teleoginas en los tubos de malla (Davey, 1980) y/o a las condiciones adversas en el sitio de exposición en la pradera.
El uso del IEC en este tipo de observaciones no aporta directamente a la comprensión de la capacidad reproductiva de
El uso conjunto del IEC y el porcentaje de eclosión en un ambiente determinado daría un mayor acercamiento a la comprensión de este fenómeno.
El IPR se ha desarrollado tratando de obviar este problema para conocer el número de larvas producido por un peso conocido de una hembra. El IPR encontrado en estas observaciones presenta gran variación por no tenerse experiencia en el manejo de este parámetro. Se recomienda realizar ensayos bajo condiciones más controladas para poder indicar su uso bajo condiciones de campo como medida de la cantidad de larvas que es capaz de producir una teleogina de determinado peso.
Conclusiones
El período adulto larva (PAL) no presentó variaciones al exponer las garrapatas en diferentes gramíneas en materas; sólo se encontraron pequeñas variaciones de acuerdo a la época de exposición aparentemente relacionada con la temperatura del medio, variando entre 27 y 32 días.
Las exposiciones en tubos de malla son valederas cuando se requiere observar de cerca el proceso de desarrollo pero pueden causar un ligero alargamiento de este período.
De acuerdo con estos resultados la progenie de una garrapata que cae al suelo aparecerá en aproximadamente cuatro semanas, por lo que simples tratamientos acaricidas aplicados a un intervalo mayor en el ganado no causarán reducciones substanciales en la población de larvas en el potrero.
La supervivencia larvaria (SL) fue determinada con mayor precisión mediante la exposición en materas, las que no se podan durante el período de observación. La SL encontrada durante los meses secos (noviembre-marzo) es resultado de las adversas condiciones ambientales para las larvas, siendo menor en
La longevidad total (LT) es lo bastante corta para poder recomendar el uso de la práctica de descanso de potreros como estrategia en el control no químico de
Las larvas sobreviven por períodos más cortos en
El uso de métodos para cuantificar progenie en este tipo de estudios está poco desarrollado. La introducción del parámetro IPR debe ser ajustada a experimentación previa en condiciones controladas. Se debe ajustar el uso del IEC a condiciones de campo y buscar la manera de extrapolar los valores de número de huevos producidos con el porcentaje de eclosión de los huevos con el fin de conocer la capacidad reproductiva de este parástio en ambientes cambiantes.
La importancia que está adquiriendo
Las peculiares condiciones de manejo de las sabanas naturales existentes en la altillanura, incluyendo manejo extensivo, quemas secuenciales, uso de bajos y esteros (Mullenax, 1982) requieren el profundizar conocimientos en estas áreas para poder recomendar métodos alternos de control que no afecten la estabilidad enzoótica de
Footnotes
Agradecimientos
A los doctores Eduardo Aycardi y Obed García de la Sección Salud Animal del Programa de Pastos Tropicales del CIAT y al doctor Guillermo Mateus del Programa de Parasitología del ICA, por sus comentarios y colaboración para la realización de este trabajo, como al Auxiliar del Programa señor Adonay Rojas.
Al doctor Antonio Betancourt, Director del Programa de Parasitología por sus comentarios críticos sobre el manuscrito de este trabajo.