
Abstract
Lima bean (
Introduccion
El lorito verde, Empoasca kraemeri Ross & Moore es una de las plagas más importantes del fríjol en América Latina; en ciertas zonas y bajo circunstancias favorables puede llegar a causar la pérdida total en la producción (Schwartz et al, 1978). Este insecto está ampliamente distribuido en América Central y del Sur, México y el Caribe. Ocurre en una amplia gama de hospedantes, entre los cuales los más importantes son las especies de Phaseolus, P. vulgaris, P. acutifolius, P. coccineus, P. lunatus y P. filiformis. Además, se le encuentra en algodonero, papa, batata, cebada, maiz, alfalfa, maní y tabaco (Schoonhoven y Cardona, 1980; Langlitz, 1966).
El daño de E. kraemerei es más severo durante épocas secas y cálidas y durante la floración y la formación de vainas (Schoohoven et al, 1978). Los síntomas del daño son amarillamiento, secamiento y doblamiento hacia abajo de las hojas, seguidos por necrosis en la punta y márgenes de las hojas. La planta se atrasa y se achaparra, produce menos vainas y éstas se deforman y no llenan completamente (Bonnefil 1965; Schwartz et. al, 1978). El daño es una consecuencia de la extracción de savia del floema por el insecto, lo cual resulta en desorganización y taponamiento de los vasos (Schoonhoven y Cardona, 1980). Ni la calificación visual del daño, ni los recuentos de insectos predicen adecuadamente las pérdidas de producción en fríjol común (Avalos 1978; Schoonhoven y Cardona, 1980).
La obtención de variedades de fríjol común resistentes al lorito verde ha progresado con relativa lentitud, porque no se conocen a fondo ni los mecanismos de resistencia ni las bases genéticas de la misma.
En fríjol común (P. vulgaris), la resistencia parece ser recesiva y cuantitativa (CIAT, 1976). La heredabilidad es poca (CIAT, 1977) y la tolerancia parece ser el mecanismo responsable (Schoohoven y Cardona, 1980). En fríjol lima (P. lunatus) no hay información relacionada con la resistencia a E. kraemeri. Sin embargo, Eckenrode y Ditman (1963), McFarlane y Riedman (1943) y Wolfenbarger y Sleesman (1961) hallaron grados diversos de resistencia del fríjol lima a la especie de lorito E. fabae Harris, la cual es muy similar al E. kraemeri.
La presencia de pilosidad en el envés de las hojas ha sido relacionada con frecuencia con la resistencia al salta hojas, pero este es un aspecto controversial. Variedades de fríjol común y de soya con tricomas erectos fueron menos atacadas que variedades glabras o con tricomas postrados (Pillemer y Tingey, 1976, 1978; Poos y Smith, 1931; Wolfenbarger y Sleesman, 1963). Al estudiar la, captura de ninfas por los tricomas en forma de gancho se encontró que era mucho mayor en variedades con densidades de 2000/cmβ que en variedades con 400 tricomas por cmβ (Pillemer y Tingey, 1976, 1978). En estudios similares, Schoonhoven y Cardona (1980) observaron baja mortalidad de ninfas de E. kraemeri por captura de tricomas en fríjol común y no obtuvieron correlación significativa entre la densidad de tricomas y la calificación visual del daño. No se hallaron registros en la literatura sobre la herencia de la densidad de tricomas ni en fríjol lima ni en fríjol común.
Los objetivos de este trabajo fueron evaluar por su resistencia a Empoasca todas las líneas de fríjol lima disponibles, identificar las más resistentes y las más susceptibles, cruzar una resistente y una susceptible, evaluar las progenies F2 por su respuesta al lorito verde, relacionar el daño con la pilosidad de las variedades y estimar la forma de herencia de estas características en fríjol lima.
Materiales Y Metodos
Evaluación de la resistencia
El germoplasma de fríjol lima fue evaluado en 1979 en la Estación principal del Centro Internacional de Agricultura Tropical (CIAT) localizado en Palmira, Valle del Cauca, Colombia. Inicialmente se estudiaron 185 cultivares, de los cuales 48 fueron reevaluados en el semestre siguiente; los promedios de temperatura y humedad relativa fueron de 28°C y 80% respectivamente. Las parcelas fueron de un surco de 3 m de longitud sembrados a 1 m y con distancia de 10 cm entre plantas. Se utilizaron testigos de fríjol común susceptible y resistentes. Se utilizó un diseño de bloques completos al azar con dos replicaciones. No se aplicaron insecticidas ni fungicidas y las prácticas culturales y la frecuencia de riegos fueron las usuales en el cultivo.
Los síntomas de daño por el ataque del lorito fueron evaluados de acuerdo con la siguiente escala:
0= no hay daño
1= daño leve con ligero amarillamiento de los bordes de las hojas.
2 = daño moderado con encocamiento de las hojas
3=daño moderado acompañado por amarillamiento de los bordes de las hojas y achaparramiento de la planta
4= daño severo, con más encocamiento de las hojas, con más amarillamiento y achaparramiento mayor
5= daño muy severo, con fuerte encocamiento y amarillamiento de las hojas, achaparramiento severo, reducción sustancialdel número de flores y vainas/planta; frecuentemente, muerte de la planta después de la floración.
Las evaluaciones de daño se hicieron a los 36 45 y 60 días después de la siembra y los valores obtenidos se interpretaron de acuerdo con la siguiente escala:
0,0-2,0 resistente
2,1-3,0 intermedio
3,1-5,0 susceptible
Los promedios de daño fueron ordenados por mérito y se calculó el coeficiente de correlación de rangos (Spearman) para los materiales que se evaluaron en ambos semestres. Las poblaciones de ninfas se estimaron a los 36, 45 y 60 días después de la siembra mediante recuentos en 10 hojas trifoliadas tomadas al azar por replicación. Los promedios de ninfas/trifolio se correlacionaron entonces con los promedios de calificación visual de daño.
Se contó el número de tricomas en forma de gancho en el envés de las hojas de 20 variedades escogidas por su suceptibilidad o resistencia. Para esto se tomaron 4 muestras por variedad por replicación, cortando secciones de aproximadamente 3 cmβ de lámina foliar por muestra. Con la ayuda de un microscopio estereoscópico se contó el número de tricomas por cmβ. Con base en observaciones preliminares se establecieron las siguientes categorías: densidad alta para recuentos mayores de 500 tricomas/cmβ y densidad baja para recuentos inferiores a 500 tricomas/cmβ.
Hibridación y evaluación de progenies.
Para adelantar los estudios de herencia se escogieron dos líneas: una resistente (G25172) y una susceptible (G 25031). Se hicieron los cruzamientos recíprocos entre ellas. La semilla F1 se multiplicó por autopolinización; la F2 se puso a germinar en una incubadora y las plántulas de ocho días de edad fueron transplantadas al campo.
Las evaluaciones para daño del Empoasca se hicieron tanto en las líneas parentales como en cada una de las plantas F2 a los 40, 47 y 54 días después de la siembra. Se hicieron también recuentos de tricomas en los padres, en la F1 y en la F2.
Los análisis estadísticos se hicieron con transformación de los datos a log (x + 1). Las medias ponderadas de daño en los padres y progenitores se compararon por la prueba de rangos de Student cuando las varianzaseran iguales o por la prueba de aproximación de t de Cochran cuando las varianzas eran disímiles (Snedecor y Cochran, 1967). Para calcular el tamaño de población requerido para encontrar por lo menos una planta con alta densidad de tricomas en la F2, se utilizó la fórmula de Muller (1923). Se hizo regresión lineal para daño en función de tricomas, tanto en la evaluación de germoplasma como en la evaluación de los padres y de la población F2.
Resultados Y Discusion
Evaluación de la resistencia.
La alta infestación de lorito verde en estos experimentos (más de 10 ninfas/trifolio en promedio) permitió clasificar como susceptibles un poco más del 60°/o de las variedades de fríjol lima evaluadas. Sólo 4 líneas (2°/o) fueron clasificadas como resistentes (Tabla 1). Estas líneas mostraron un nivel de resistencia mejor que los encontrados hasta ahora en P. vulgaris, especie de fríjol para la cual sólo niveles moderados de resistencia han sido detectados (CIAT, 1975, 1977; Schoonhoven y Cardona, 1980).
Evaluación del daño causado por Empoasca kraemeri en variedades de fríjol lima.
En escala visual de 0 a 5 (0= no hay daño; 5 = daño muy severo).
Reevaluación de 48 variedades.
Las poblaciones de ninfas variaron entre 5 y 41 por hoja trifoliada con un promedio de 14,8/trifolio. Se obtuvo una correlación altamente significativa (r = 0,45**) entre los recuentos de ninfas y las calificaciones visuales de daño, lo cual indica que la expresión de resistencia sí fue afectada por las poblaciones. Esta clase de relación nunca ha sido hallada para el fríjol común (Schoonhoven y Cardona, 1980).
Los recuentos de tricomas variaron entre 21 y 1074/cmβ. La correlación entre el número de tricomas y las calificaciones de daño fue negativa y altamente significativa (r=-0,87**) (Figura 1), lo cual sugirió que la pilosidad tiene un efecto importante como factor de resistencia. En efecto, las variedades con mayor densidad de tricomas fueron las menos dañadas.
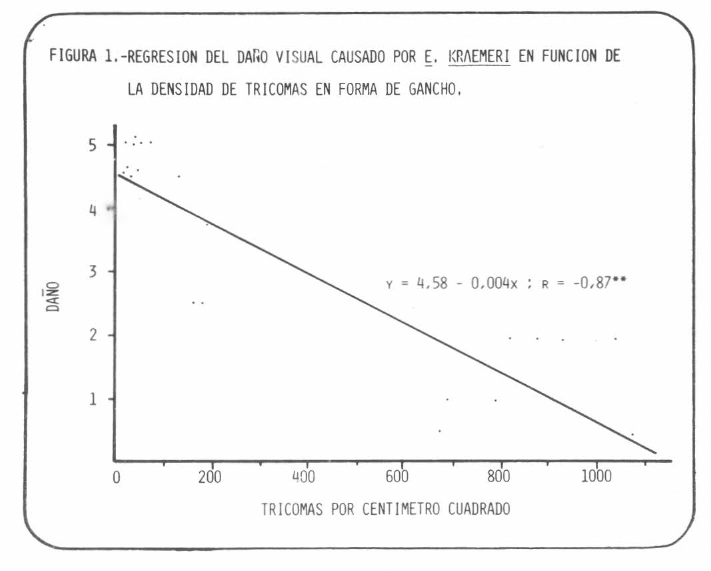
REGRESION DEL DAÑO VISUAL CAUSADO POR E. KRAEMERI EN FUNCION DE LA DENSIDAD DE TRICOMAS EN FORMA DE GANCHO. Y = 4,58 - 0,004x R = -0,87**
Cuando se reevaluaron 48 materiales, más del 60°/o fueron clasificados como de resistencia intermedia (Tabla 1). La correlación de rangos entre semestres fue altamente significativa (rs= 0,69**), lo cual confirmó la estabilidad de la reacción en diferentes semestres.
Las relaciones entre las cuentas de ninfas, daño y densidad de tricomas encontradas en fríjol lima en estos estudios fueron consistentes con resultados obtenidos con E. fabae en fríjol común (Pillemer and Tingey, 1976, 1978). Sin embargo, difieren de resultados previos con E. kraemeri en fríjol común (CIAT, 1976, 1977).
Las cuatro líneas de fríjol lima catalogadas como más resistentes fueron colectadas en el Perú; todas tienen follaje gris plateado, con hojas grandes y pubescentes y son de crecimiento indeterminado. Todas son de semilla blanca. Las variedades susceptibles son más heterogéneas, pero la mayoría tienen follaje verde oscuro, hojas pequeñas y mucho menos pilosas. Eckenrode y Ditman (1963) señalaron que las variedades de fríjol lima de semilla grande tenían menor daño por E. fabae que las de semilla pequeña, pero Wolfenbarger y Sleesman (1961) concluyeron que el tamaño de semilla no guardaba relación con el nivel de resistencia a E. fabae. En nuestros estudios tampoco el tamaño de semilla fue un factor útil para clasificar la resistencia de las variedades.
Hibridación y evaluación de progenies
La hibridación se verificó por la segregación de las progenies F2 tanto para resistencia al insecto, como para densidad de tricomas. Las calificaciones visuales de daño para F2 fueron intermedias en comparación con las que recibieron los padres y significativamente diferentes de ellas (Tabla 2). La naturaleza continúa de la segregación para la reacción al insecto observada en la F2 (Figura 2) sugiere herencia cuantitativa para esta característica en fríjol lima. Esta observación concuerda con la conclusión de que la reacción a Empoasca también se hereda cuantitativamente en fríjol común (CIAT, 1976).
Evaluación del daño causado por Empoasca Kraemeri, en líneas parentales y en plantas F2 de un cruce de fríjol lima.
Las cifras seguidas por la misma letra no son significativamente diferentes al nivel del 5°/0.
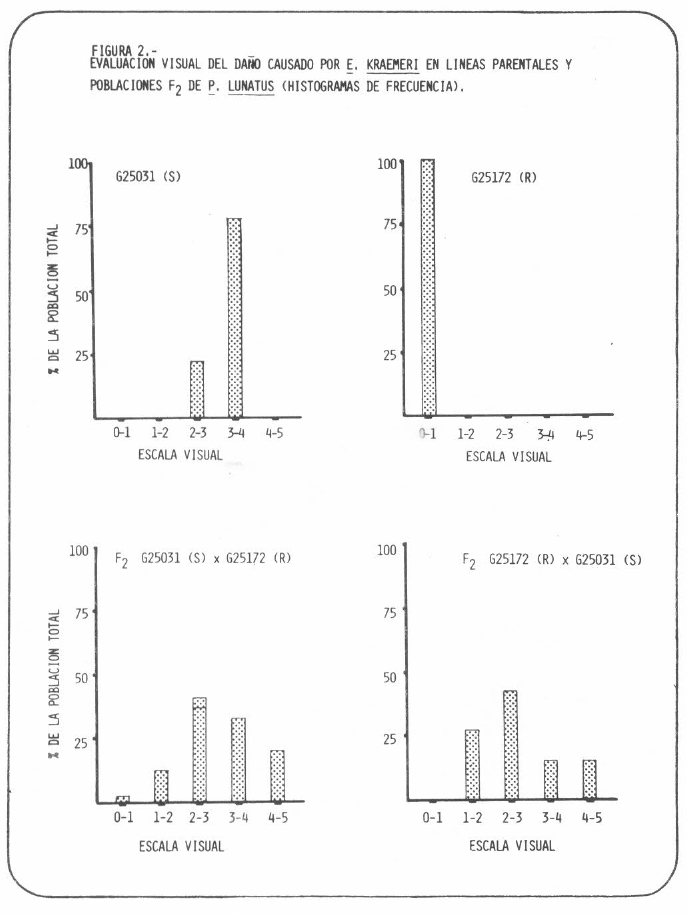
EVALUACION VISUAL DEL DAÑO CAUSADO POR E. KRAEMERI EN LINEAS PARENTALES Y POBLACIONES F2 DE P. LUNATUS (HISTOGRAMAS DE FRECUENCIA).
El daño promedio de la progenie fue significativamente mayor que el del promedio de los padres. Esto indica dominancia de la reacción susceptible, porque la diferencia entre daños promedios para las progenies recíprocas no fue significativa. Estos resultados coinciden con registros de dominancia de la reacción susceptible en cruces interespecíficos entre líneas resistentes y susceptibles de fríjol común, fríjol lima y Phaseolus coccineus (CIAT, 1976; Wolfenbarger y Sleesman, 1961).
Las densidades de tricomas en los padres, la F1 y la F2 aparecen en la Tabla 3. El promedio de tricomas/cmβ en la F1 no fue significativamente diferente del promedio del padre con baja densidad; en la F2 el promedio fue intermedio entre los promedios de los padres y significativamente diferente de ellos. El promedio general de todas las progenies fue significativamente menor que el de los padres, tanto en la F1 como en la F2; estos resultados sugieren dominancia de la baja densidad de tricomas. La segregación para esta característica fue contínua en la F2, lo cual sugiere herencia cuantitativa. La correlación entre el daño y la densidad qe tricomas en la F2 fue negativa pero no significativa (r=-0,11 N.S.). Esto difiere ampliamente del valor altamente significativo (r=-0,96**) encontrado entre el daño de los padres y sus densidades de tricomas; por lo cual se infiere que al iniciarse el mejoramiento de P. lunatus por resistencia a Empoasca esta clase de análisis debe también hacerse en la F3 y en generaciones sucesivas, con el fin de confirmar la verdadera importancia de esta característica como factor de resistencia al insecto.
Densidad de tricomas/cmβ de hoja en líneas parentales, plantas F1 y plantas F2 de un cruce de fríjol lima.
En cada evaluación, las cifras seguidas por la misma letra no son significativamente diferentes al nivel del 5°/0.
Los resultados de estos estudios sientan las bases para iniciar el mejoramiento del fríjol lima por resistencia a E. kraemeri. Dado que la herencia de la resistencia al insecto es cuantitativa, hay dominancia de la reacción susceptible y no existen muchas fuentes de resistencia, lo más conveniente sería escoger un esquema de selección recurrente con prueba de progenies. Este es un sistema flexible (Allard, 1960) que permite la incorporación de padres resistentes en cualquiera de los ciclos de selección.
Conclusiones
Se encontraron niveles de resistencia a Empoasca kraemeri Ross & Moore superiores a los que ocurren en fríjol común (Phaseolus vulgaris L.).
Contrario a lo que ocurre en el fríjol común, se encontró una correlación positiva y altamente significativa entre el número de ninfas presentes y la calificación visual del daño.
El número de tricomas presentes en la hoja, tuvo un efecto importante como factor de resistencia, o sea, materiales con mayor número de ellos, presentaron menores calificaciones de daño.
Cuando se realizaron cruces entre materiales resistentes y suceptibles, se encontró dominancia de la reacción susceptible y de la baja densidad de tricomas.