
Abstract
The soil arthropods are an important component of the soil biota for their species diversity, function, and as tool to evaluate the habitat conservation state. The conservation state of three sectors in an ecosystem of high mountain (Socha, Tasco and Socotá in Boyacá, Colombia) was evaluated measuring alpha and beta diversity of epigean arthropods assemblage in particular Acari and Collembola. Pitfall traps were installed on transects for each sector; the alpha diversity was measured through Berger-Parker index and a gamma-binomial model was used to obtain a parameter α, which describes the fitting to species abundance distribution. The species diversity estimation was established with coverage-based rarefaction and extrapolation curves, and the beta diversity was measured in species replacement and richness difference. The alpha diversity did not show statistical differences between sectors, but with the highest species richness in the sector of Socotá. The a parameter indicated that the sector of Socotá is the better conserved; similar to that obtained with Acari and Collembola. The spatial turnover or species richness difference showed changes with respect to the sector and taxonomic group (Acari or Collembola), explained by vegetation structure, erosion, and aggregate distribution of epigean arthropods. The species replacement did the main contribution to total beta diversity by sectors while in Acari and Collembola the species replacement was highest in Tasco in contrast to the richness difference in Socha and Tasco. The utility of epigean arthropods to evaluate the soil quality and the partition of the beta diversity in their components was discussed.
Introducción
Los artrópodos del suelo son un componente importante de los ecosistemas por su alta diversidad de especies (Giller 1996) y por su papel en la descomposición de materia orgánica, ciclaje de nutrientes y regulación de las comunidades bióticas dentro o fuera del suelo (Brussaard
La alta diversidad de especies ha sido explicada principalmente por la heterogeneidad espacial del hábitat en el suelo que favorece la repartición de recursos y la coexistencia e interacción entre especies (Ettema y Wardle 2002; Wardle 2006). También se puede explicar por la alteración que tienen otros componentes bióticos sobre los artrópodos del suelo, por ejemplo, el tipo y calidad de hojarasca (Wardle
No obstante, los patrones de diversidad de especies de los artrópodos epigeos, en general, son poco conocidos en ecosistemas de alta montaña. Algunos trabajos han sido dirigidos a establecer patrones de diversidad relacionados con gradientes altitudinales (Olson 1994; Leakey y Proctor 1987; Thomas y Proctor 1997), uso de suelo, estado de conservación y estructura de la vegetación (Garay 1981; Morales y Sarmiento 2002; Cerón y García 2009).
Por medio de experimentos de restauración ecológica, se ha demostrado que los artrópodos epigeos, tienden a presentar una mayor abundancia y riqueza de morfoespecies en bosques que actúan como referencia en tratamientos con vegetación regenerante o plantaciones (Cole
En este trabajo, se evaluó el estado de conservación de tres sectores o localidades de colecta, en sistemas de alta montaña por medio de la medición de la diversidad alfa y beta de las comunidades de artrópodos epigeos y con énfasis en Collembola y Acari, justificado por la alta dominancia, en términos de biomasa (Harding y Stuttard 1974), diversidad de especies (Bardgett y Van der Putten 2014) y por su papel como indicadores biológicos de la calidad del suelo (Maraun y Scheu 2000; Ponge
Materiales y métodos
Sitio de estudio
Se escogieron tres sectores de laderas, con base en mapas satelitales y de acuerdo a las coberturas vegetales presentes, ubicados sobre la Cordillera Oriental de Colombia, departamento de Boyacá, con una distancia promedio de 12,5 km entre sí, que corresponden a los municipios de Socotá (6°02′46″N y 72°37′20″O) entre 2.850 y 3.230 msnm; Socha (5°58′56″N y 72°46′37″O) entre 2.800 y 3.200 msnm y Tasco (5°52′59″N y 72°46′37″O) con un intervalo altitudinal comprendido entre 2.900 y 3.195 msnm. Los tres sectores pertenecen a la asociación edáfica Pisba, que presenta circos glaciares, escarpes, afloramientos rocosos y distintos procesos erosivos, con pendientes entre 12-25 % y 25-50%, y forman parte de la unidad geomorfológica glacio-estructural (IGAC 1981; Meneses
Vegetación
En Socha la cobertura vegetal no es continua y se asocia con eventos de perturbación. Predominan los estratos arbustivo y herbáceo y las especies más abundantes son
Colecta de artrópodos
Para cada sector se ubicaron cinco transectos cada 100 m perpendiculares a la pendiente. En cada transecto se instalaron cinco trampas de caída a una distancia de 5 m entre sí (Olson 1994; Ward
Diversidad alfa
Para la medición de la diversidad de especies se utilizó el índice de dominancia de Berger-Parker, que describe la importancia relativa de la especie más dominante en un ensamblaje (Magurran 2004). Dado que ha sido probado bajo distintos usos de suelo, con un aumento de su valor desde suelos no perturbados a perturbados, es sugerido como una herramienta en el monitoreo ambiental de ecosistemas de suelo (Caruso

Aspecto general de los sectores de colecta. Los sectores donde se presenta mayor contraste en la estructura del hábitat son:
Por último, se evaluó el ajuste del ensamblaje de artrópodos a modelos de abundancias de especies por medio del modelo gamma-binomial (Gambin). La forma de la curva de distribución de abundancias es descrita por un parámetro α, adecuado para realizar interpretaciones ecológicas (Ugland
Diversidad beta
Para evaluar el efecto de la distancia geográfica entre puntos de muestreo sobre el ensamblaje de artrópodos, se realizó la prueba de Mantel (Borcard
La diversidad beta se estimó de acuerdo con la aproximación de Baselga (2010) y Carvalho
Resultados
Diversidad alfa
Se recolectaron un total de 2.001 individuos y 455 morfoespecies para los tres sectores, de los cuales Socotá presentó 964 individuos y 166 morfoespecies; Socha, 482 individuos y 142 morfoespecies y Tasco, 555 individuos y 147 morfoespecies. La clase Acari obtuvo la mayor riqueza de especies, y Collembola el grupo con mayor abundancia en los tres sectores (Tabla 1). Otros grupos predominantes fueron Araneae, Isopoda, Blattariae, Hymenoptera (Formicidae), analizados dentro del ensamblaje de artrópodos. El índice de diversidad de Berger-Parker presentó diferencias significativas entre sectores (F = 4,548; df = 2; P < 0,05) que, en promedio, fue mayor en Socotá (0,3052), comparado con Socha (0,1546) y Tasco (0,1306; Tabla 2).
En la figura 2 se presentan las curvas de rarefacción-extrapolación, basadas sobre coberturas de abundancias para los tres sectores. Socotá con 166 morfoespecies presentó una cobertura del 91 %, mientras que, en Socha y Tasco, la cobertura varió entre 83 y 87 %, y al extrapolar la riqueza, varió entre 90 y 92 %, respectivamente. Las curvas para Socha y Tasco no presentaron diferencias en la riqueza de especies, pero sí, con respecto a la curva de Socotá, donde la riqueza de especies es mayor (Fig. 2).
El parámetro α obtenido a partir de modelos Gambin fue de α = 0,458, AIC = 243,2 en Socotá; α = 0,706, AIC=315,6 en Socha, y α=0,74 y AIC=317,2 para Tasco. Los valores altos de α reflejan una distribución log-series, en áreas transformadas o perturbadas. Al considerar Acari y Collembola, los valores de diversidad (Fig. 3A) y parámetro α (Fig. 3B), indican que el gradiente de Socotá es el mejor conservado en contraste con los de Socha y Tasco.
Diversidad beta
El ensamblaje de especies no mostró correlación con la distancia geográfica en Socha (r = -0,0303; P = 0,559), Tasco (r = -0,24; P = 0,812) y Socotá (r = 0,06; P = 0,472), debido al intervalo estrecho altitudinal considerado en este estudio. La diversidad beta total (βtotal) promedio en Socha fue de 0,91, seguido por Socotá (0,88) y Tasco (0,85). A nivel del ensamblaje total de especies, el reemplazamiento de especies realizó la mayor contribución a la βtotal, que osciló entre 0,48 para Socotá, y 0,61 en Tasco; mientras que la diferencia de especies (βdif) realizó la menor contribución a βtotal en los tres sectores (Fig. 4A).
Para el ensamblaje de Acari, el reemplazamiento de especies es el componente más importante en Socotá (0,49) y Tasco (0,53); al contrario de Socha, donde la diferencia de especies es de 0,67. Dentro de Collembola, la diferencia de especies fue representativa para Socotá (0,49) y Socha (0,70), mientras que, en Tasco, la ßtotal fue explicada por el reemplazamiento de especies (0,56; Fig. 4B, C).
Discusión
Los sectores muestreados mostraron alta riqueza de especies (455 morfoespecies), si se comparan con otros estudios y en intervalos altitudinales similares. Morales y Sarmiento (2002) encontraron 74 morfoespecies (con presencia de otros taxones) en un ecosistema de páramo venezolano, entre los 3.300 y 3.800 m de altitud; Smithers y Atkins (2001) colectaron 20,2 especies en promedio por trampa de caída, ubicadas a lo largo de un gradiente entre 3.730 y 4.150 msnm; González
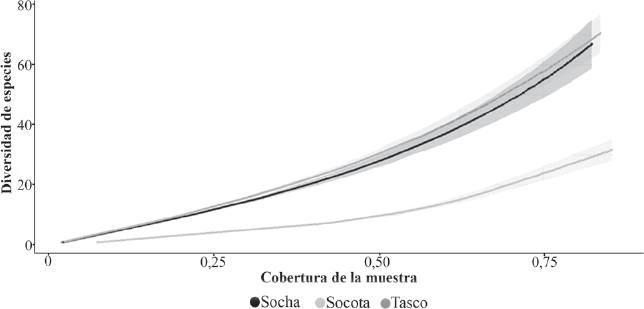
Curvas de rarefacción y extrapolación basadas sobre cobertura de muestra “simple covarege” para los tres sectores muestreados.
Riqueza y abundancia total por orden dentro de cada sector.
Con base en las curvas de rarefacción y extrapolación, se obtuvo un muestreo casi completo en Socha y Tasco. No obstante, la curva de extrapolación y con una muestra de 1,027 individuos indicó una diversidad estimada menor en Socotá en contraste a los demás sectores. Entre los factores que pueden explicar lo anterior se considera, el papel de la estructura del hábitat, por ejemplo, en sectores con vegetación más densa, los métodos de colecta pueden ser menos efectivos, lo cual se refleja en la riqueza observada. Por otra parte, puede ser un efecto debido a la baja intensidad del muestreo que eleva la riqueza extrapolada. Sin embargo, el método empleado al no basarse sobre el tamaño de la muestra (Chao y Jost 2012) puede ser adecuado para realizar comparaciones entre sitios y en estimar la diversidad de grupos taxonómicos en los que se requiere esfuerzos de colecta importantes no obstante, el papel de las especies raras en la estimación de la diversidad de especies.
En bosques secos tropicales, Cabra-García
La dominancia en riqueza de especies de Acari y Collembola, ha sido ampliamente documentada en otros ecosistemas (Giller 1996; Lavelle 1996), lo cual corresponde a lo hallado en este estudio. Pocos trabajos abordan la diversidad de especies en estos taxones, limitando una discusión acerca de los patrones o tendencias en su diversidad en ecosistemas de alta montaña. Se puede mencionar el trabajo de Díaz
Como factores que explican la alta dominancia de Collembola y Acari son: las características del suelo y hojarasca, otros componentes no evaluados puede explicar la riqueza observada, e.g. estructura de la vegetación (Hansen 2000; Wardle
Los criterios empleados para evaluar la calidad del hábitat, el índice Berger-Parker y el parámetro α, reflejaron las condiciones de perturbación locales dentro de cada sector sugieren que los cambios en la cobertura de vegetación y los procesos de erosión presentes, de tipo laminar y que caracterizan los sectores muestreados, son los más importantes en los sitios de estudio. El parámetro α es apropiado para detectar transiciones de áreas perturbadas a no perturbadas, y esto ha sido demostrado para otras comunidades de artrópodos epigeos (Matthews
La diversidad beta total alta, a nivel de todo el ensamblaje de artrópodos, con el reemplazamiento de especies como el componente que realizó el mayor aporte, es reportado bajo otros métodos y escalas espaciales (González
El reemplazamiento de especies, obtenido a una escala espacial relativamente pequeña, es notable en este estudio. Boieiro
Al contrario del ensamblaje de artrópodos, la diferencia de especies realizó la mayor contribución a la diversidad beta total para Acari y Collembola, cuyos valores soportan el enunciado de un patrón no aleatorio en la riqueza de especies para estos grupos taxonómicos (Fig. 4). Además de la disponibilidad espacial de recursos, cabe resaltar el papel de la pérdida y ganancia entre ensamblajes de especies. Como lo mencionan Lindo
Descriptores de la diversidad alfa para cada transecto en los tres sectores.
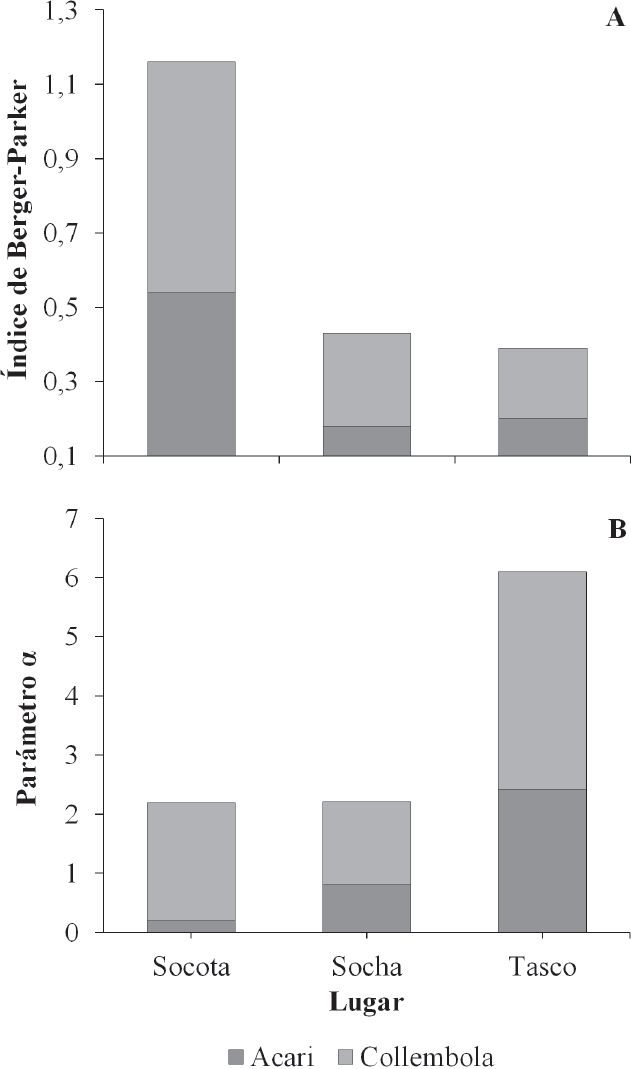
Descriptores de la diversidad alfa y parámetro α (modelo de distribución de abundan cias) para los órdenes Acari y Collembola.
Altos valores de diversidad beta han sido informados especialmente para Acari (Caruso
En este estudio se sugiere que procesos erosivos y la geomorfología, hacen que la contribución del reemplazamiento o diferencia de especies difiera localmente, ya sea a nivel de taxones o en todo el ensamblaje. El proceso erosivo presente en los sectores muestreados corresponde principalmente al tipo laminar. Las capas superficiales del suelo al ser removidas por escorrentía, ocasionan que la capa orgánica se deposite en láminas a lo largo de la pendiente junto al establecimiento de la vegetación, lo cual puede modular la distribución espacial de los artrópodos del suelo, sin embargo, la evaluación de su efecto real puede presentar dificultades dado que involucra la separación de otros factores relacionados, por ejemplo, propiedades del suelo (Fig. 1).
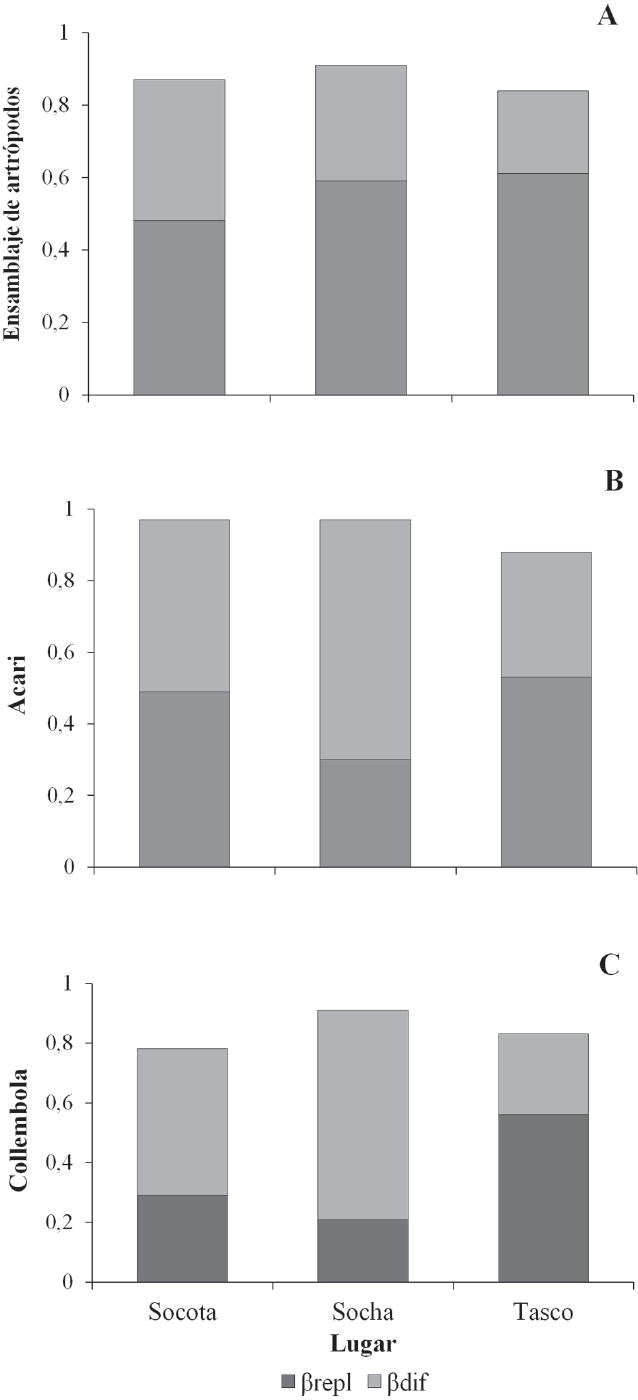
Contribución del reemplazamiento de especies (βrepl) y diferencia de especies (βdif) a la diversidad beta total para (A) el ensamblaje de artrópodos, (B) Acari y (C) Collembola.
Según Legendre (2014) la diferencia de especies (βdif) refleja la riqueza que puede contener una comunidad. De acuerdo con lo anterior, una condición fragmentada de la vegetación, al limitar la capacidad de dispersión de las especies, puede promover este componente de la diversidad beta total (Lindo y Winchester 2007; Calderón-Patrón
Conclusiones
En este estudio, el contraste de métodos para la medición de la diversidad alfa y beta, permite realizar evaluaciones rápidas del estado de conservación del hábitat, más aun, cuando son aplicados a grupos taxonómicos sensibles a factores estresantes como son los Acari y Collembola. El parámetro α es una herramienta práctica en planes de conservación, aunque puede ser afectado por el tamaño de la muestra y puede ser trabajado junto a otros índices, ya probados experimentalmente, en este caso, el índice de Berger-Parker. Los sectores muestreados se caracterizan por presentar poca cobertura vegetal y suelos erosionados, no obstante, la diversidad de especies es alta, si se contrasta con trabajos en hábitats con suelos mejor establecidos. Sin embargo, trabajos de diversidad en sistema de alta montaña, son relativamente escasos o realizados bajo otros métodos no comparables, y no menos, el conocimiento de los procesos detrás de esta diversidad de especies. El método empleado para medir la diversidad beta, destaca cómo el ensamblaje de artrópodos epigeos es afectado por factores que actúan a una escala pequeña, y otros poco considerados como la erosión o la geomorfología que modulan la distribución de la riqueza de especies. En términos de conservación, la diversidad beta, sugiere cómo la riqueza de especies responde a cambios en el hábitat, con valores más bajos en sistemas intervenidos, y que se reflejan en el reemplazamiento y diferencia en la riqueza de especies. Se sugiere contrastar experimentalmente distintos tipos de uso de suelo o a lo largo de un gradiente de perturbación con el fin de poder reconocer las causas que pueden explicar la contribución de la diferencia en la riqueza de especies y como ello modula las comunidades de artrópodos epigeos.
Footnotes
Agradecimientos
A Carolina Camargo B. y Laura Ortega por sus comentarios al manuscrito. A Carolina Acevedo por su colaboración en el trabajo de campo y habitantes de los Municipios de Socotá, Socha y Tasco. A la Dirección de Investigación DIN-UPTC. El presente artículo es resultado del convenio de cooperación número 13-12.092-080CE entre el Instituto de Investigación de Recursos Biológicos “Alexander von Humboldt” y la Universidad Pedagógica y Tecnológica de Colombia, proyecto “Estudio biótico para la delimitación del complejo de páramos Pisba-Boyacá”. Un agradecimiento especial a los evaluadores anónimos por las sugerencias hechas al manuscrito.