
Abstract
Abstract
Hornworm (Erinnyis ello) and lace bug (Leptopharsa heveae) are considered the most important pests of rubber tree (Hevea brasiliensis). This study aimed to evaluate incidence and abundance of E. ello and L. heveae in three rubber tree clones (two promising clones and control IAN 873) in different planting systems with copoasu trees (Theobroma grandiflorum) and banana trees (Musa AAB) in two localities of Caquetá (Colombian Amazonia). A completely randomized design with three replications and split-plot arrangement, analyzed with a general linear mixed model, was used in each locality. Very low means of incidence and abundance in both of E. ello (<5% and < 1 insect per 100 leaflets per plant) and L. heveae (<7% and < 2 insects per plant per leaflet) were found for all factors studied, after 23 months of monitoring (six rainfall seasons). It was concluded that the incidence and abundance of E. ello and L. heveae in H. brasiliensis, are influenced not only by spatial and temporal variation, but also by implemented planting system, given by the fact the insect pests are present in the same rainfall season. Similarly, a season effect was observed in clonal reaction to L. heveae, where the clone FDR 5788 was the most tolerant for the four planting systems evaluated in this research.
Introducción
El caucho natural [Hevea brasiliensis (Willd. Ex Adr. de Juss.) Muell.-Arg.] es una especie de origen suramericano productora de látex destinado principalmente a la industria llantera (Compagnon 1998). A nivel mundial, el sudeste asiá- tico produce el 91 % del caucho natural mientras que América Central y del Sur representan el 3 % de la producción (IRSG 2013).
En Colombia, a diciembre de 2014, se reportaron 49.500 ha cultivadas de caucho, constituidas principalmente por clones introducidos de origen asiático, africano y americano (Becerra 2014). Caquetá es el departamento con mayor tradición cauchera de Colombia y el tercero en área sembrada con 6.017 ha, de las cuales 1.581 ha corresponden a plantaciones en etapa productiva con 1.894,4 toneladas de caucho seco por año (Asoheca 2014).
Dada la importancia del caucho natural y su creciente demanda en el mercado internacional, esta situación impone al sector heveícola colombiano el reto no sólo de incrementar las áreas de siembra sino además identificar, para las diferentes regiones del país, los clones apropiados con mejor desempeño y tolerancia a las principales enfermedades y plagas del cultivo (Castellanos et al. 2009; Sterling et al. 2009; Sterling y Rodríguez 2011, 2012; GTP 2012).
Entre las plagas del cultivo de caucho, las más importantes son el gusano cachón Erinnyis ello Linnaeus, 1758 (Lepidoptera: Sphingidae) y la chinche de encaje Leptopharsa heveae Drake & Poor, 1935 (Hemiptera: Tingidae) (Vendramin 1992; Garzón 2000; Tanzini 2006; Sterling y Rodríguez 2011; Rincón 2012).
Además del cultivo de caucho, E. ello se ha reportado en otras especies de importancia económica como yuca (Manihot esculenta Crantz) (CIAT 1989; Bellotti 2000; Fazolin et al. 2007), papaya (Carica papaya L.) (Acosta y León 2003), tomate (Solanum lycopersicum L.), tabaco (Nicotiana tabacum L.) y algodón (Gossypium hirsutum L.) (CIAT 1989). Según CIAT (1989), la etapa desde el huevo (2 mm de diámetro) hasta adulto (mariposa de 5 a 7 cm de longitud) tiene una duración aproximada de 30 días. Las larvas presentan policromías y se caracterizan por tener un cuerno caudal erecto (Belloti 2000).
La voracidad de la larva en el cultivo del caucho es tal que llega a consumir hasta 1.000 cmβ de superficie foliar, 75 % de los cuales son consumidos durante el quinto y último estadio (Garzón 2000). Los ataques de E. ello casi siempre ocurren en hojas jóvenes y avanza de arriba hacia abajo dejando sólo la nervadura central o en algunos casos únicamente el peciolo (Vendramin 1992; Garzón 2000). Según CTS (2010) una infestación severa de E. ello puede ocasionar una defoliación en el 70 % de las plantas, reduciendo drásticamente la productividad del cultivo de caucho.
Dentro del manejo de E. ello, revisten gran importancia las prácticas que integran el control cultural, físico, biológico y químico (Garzón 2000; CTS 2010; León et al. 2010). El control de E. ello depende en gran medida del estado del cultivo y del nivel de infestación (CTS 2010; Sterling et al. 2012).
Respecto a la chinche de encaje L. heveae, es originaria de la región amazónica, donde vive endémicamente en cauchales nativos y es considerada la principal plaga del cultivo de caucho en Brasil (Tanzini y Lara 1998; CTS 2010), mientras que en Colombia constituye una amenaza latente para las nuevas plantaciones de caucho natural (Garzón 2000; GTP 2012; Santos et al. 2012).
Según Tanzini y Lara (1998), la duración del ciclo de vida desde el huevo, pasando por cinco estadios ninfales (coloración amarillo-verdosa) hasta el adulto (individuo alado blanquecino de 1,35 a 1,50 mm de longitud), es afectada por la temperatura, siendo de 36 días a 20 °C y de 18 días a 30 °C. Las ninfas y adultos de L. heveae se distribuyen de manera uniforme en los diferentes estratos de la planta y en el envés de las hojas, succionan la savia y destruyen el parénquima (Tanzini y Lara 1998). Las plantas jóvenes infestadas pueden presentar una reducción del 27,7 % en altura y 43,5 % en diámetro del fuste (Moreira 1986). La producción puede ser reducida en un 30 % debido a la pérdida fotosintética y debilitamiento de los árboles infestados (Tanzini y Lara 1998).
Las principales prácticas de manejo de L. heveae involucran el uso de clones tolerantes, control químico y control biológico (CTS 2010). Tanzini y Lara (1998) concluyeron que la tolerancia clonal a L. heveae está influenciada por la no preferencia en la alimentación y la ovoposición del insecto. Asimismo, la implementación de otras prácticas culturales como el uso de sistemas agroforestales puede contribuir con la disminución de insectos plagas en el cultivo de caucho (Alvin et al. 1989). Según Pereira et al. (1997), han resultado promisorios para diversas regiones heveícolas de Brasil y del mundo, los consorcios de caucho con cultivos perennes como: cacao (Theobroma cacao L.), café (Coffea arabica L.), plátano (Musa spp.), copoazú (Theobroma grandiflorum (Willd. ex Spreng.) K. Schum) y cítricos, entre otros.
Aunque son muchas las bondades que se atribuyen al uso de los sistemas agroforestales (Pereira et al. 1997), no se tienen registros convincentes que demuestren el efecto de estos sistemas en la aparición y proliferación de insectos plagas, como E. ello y L. heveae, en el cultivo de caucho.
En general, los distintos estudios sobre E. ello y L. heveae en el cultivo del caucho, se han enfocado principalmente en la dinámica poblacional, la influencia de las condiciones ambientales y el manejo biológico (Vendramin 1992; Tanzini y Lara 1998; Garzón 2000; Cividanes et al. 2004; CTS 2010). Sin embargo, son pocos los estudios que han abordado la reacción clonal y su interacción con otros factores como el sistema de siembra frente al ataque de E. ello (Sterling et al. 2009, 2012) y de L. heveae (Tanzini y Lara 1998; Junqueira et al. 1999; Sterling et al. 2009, 2012). Al respecto se presume que la probabilidad de aparición y la proliferación de E. ello y L. heveae deben estar influenciadas no sólo por las condiciones ambientales, sino también por el clon de caucho y por el sistema de siembra implementado.
De acuerdo con lo anterior, este estudio tuvo como objetivo evaluar la incidencia y abundancia de E. ello y de L. heveae en tres clones de caucho (dos promisorios y el testigo IAN 873), en diferentes sistemas de siembra con copoazú (T. grandiflorum) y plátano hartón (Musa AAB), en dos zonas del departamento del Caquetá (Amazonia colombiana).
Materiales y métodos
Área de estudio
Se trabajó en dos zonas del departamento del Caquetá, la primera localizada en el municipio de Albania, vereda Florida (01°14′41,9″N 75°53′22,9"O, 277 msnm) con clima cálido húmedo según la clasificación de Caldas Lang (IGAC 2010). La segunda zona en el municipio de San Vicente del Caguán, vereda Buenos Aires #2 (02°01′33,6"N 74°55′07,4"O, 344 msnm) con clima cálido semihúmedo a húmedo (IGAC 2010). Se abarcaron 23 meses representados en seis épocas pluviométricas (Tabla 1), acordes con la distribución y el nivel de precipitaciones (IDEAM 2014).
Épocas pluviométricas monitoreadas en los municipios de Albania y San Vicente del Caguán (Caquetá, Colombia) en el periodo comprendido entre julio de 2012 y junio de 2014.
Medias con error estándar.
Fuente: IDEAM (2014).
Los suelos del Caquetá, presentan textura principalmente arcillosa con diferentes grados de drenaje interno. Son suelos ácidos a muy ácidos (rango de pH: 4,5 a 5,8), con un alto contenido de aluminio (> 50 %) que los hace tóxicos y de baja fertilidad (IGAC 2010).
Material vegetal
Correspondió a tres clones de caucho (H. brasiliensis) los cuales se establecieron en sistema agroforestal con copoazú (T. grandiflorum) y plátano hartón (Musa AAB). Se compararon dos clones de caucho promisorios para la región (FDR 5788 y FX 4098) con un testigo local, el cultivar IAN 873. Los clones utilizados son de origen brasilero y son producto del cruzamiento intraespecífico de progenitores de H. brasiliensis de la siguiente forma: IAN 873 (FA 1717 x PB 86), FX 4098 (PB 86 x B 110) y FDR 5788 (HARB 8 x MDF 180) (Rivano et al. 2010). Respecto a los cultivos asociados, se utilizó un material promisorio de copoazú, denominado "ecotipo sobresaliente No. 4" introducido al departamento del Caquetá en 2005 (Melgarejo et al. 2006) y como sombrío transitorio se emplearon rizomas de plátano hartón adquiridos comercialmente en la región.
Diseño experimental
En cada zona se estableció un experimento de 1,4 ha en un diseño de bloques completos al azar con tres repeticiones y un arreglo en parcelas divididas, donde los sistemas de siembra (Tabla 2) al interior de cada bloque correspondieron a las parcelas principales y los clones de caucho asociados a los sistemas de siembra constituyeron las subparcelas. La unidad experimental correspondió a cada una las plantas de caucho y se emplearon 20 plantas al interior de cada subparcela.
Sistemas de siembra de caucho (Hevea brasiliensis) en asocio con copoazú (Theobroma grandiflorum) y plátano hartón (Musa AAB) implementados en los municipios de Albania y San Vicente del Caguán (Caquetá, Colombia).
Fase de campo
Con una periodicidad mensual se realizó la inspección visual de larvas de E. ello (estadios 1 a 5) y ninfas y adultos de L. heveae en las copas de los materiales de caucho para determinar la incidencia y abundancia de ambos insectos en las dos zonas. Además, se recolectaron ejemplares de los insectos para corroborar su identificación taxonómica en laboratorio.
La incidencia correspondió al porcentaje de plantas infestadas por cada insecto. Para estimar la abundancia de E. ello, se muestrearon al azar 100 foliolos por planta y se contabilizaron las larvas. La abundancia de E. ello se clasificó según la escala de Fonseca (2009), donde: < 5 larvas = abundancia media; 5 =100 larvas = alta; 100-300 larvas = muy alta; > 300 larvas = demasiado alta. Para la abundancia de L. heveae, se muestreó el tercio medio de las copas de las plantas donde se escogió visualmente el foliolo más infestado y se realizó el conteo de insectos (ninfas y adultos), a partir del cual se empleó la escala de Fonseca (2001), donde: 1-2 insectos por foliolo = baja; 3-4 insectos por foliolo = media; ≥ 5 insectos por foliolo = alta.
Fase de laboratorio
Los ejemplares recolectados se conservaron rotulados en viales con alcohol al 70 %, y se llevaron al Laboratorio de Biología de la Universidad de la Amazonia (Florencia, Caquetá), donde se realizó su identificación partir de descripciones originales basadas en claves taxonómicas (Borror y White 1970; Coto 1997), ilustraciones y revisiones literarias (Arias y Bellotti 1984; Abreu 1996; Junqueira et al. 1999; Garzón 2000; Fonseca 2001).
Análisis estadístico
Se ajustó un modelo lineal general mixto donde los factores fijos fueron la zona, el sistema de siembra, la época de evaluación, el clon de caucho, y sus interacciones. El efecto de bloques anidados en las localidades, parcelas asociadas a los sistemas de siembra dentro de bloques y subparcelas asociadas a los clones dentro de los sistemas en cada bloque se incluyeron como efectos aleatorios.
La varianza residual se modeló para contemplar varianzas diferentes según la heterocedasticidad observada en los efectos fijos del modelo, mientras que la correlación residual para las observaciones sucesivas realizadas sobre un mismo clon (incidencia) o sobre una misma planta (abundancia) se contempló mediante un modelo de simetría compuesta. Los criterios de Akaike (AIC), Bayesiano (BIC) y LogLik se utilizaron para la selección de la estructura de varianzas y correlaciones residuales (Di Rienzo et al. 2011). El ajuste se realizó mediante la función Ime de la librería nlme (Pinheiro et al. 2013) de R (R Core Team 2013), bajo la interfaz implementada en InfoStat (Di Rienzo et al. 2013).
El análisis de las interacciones, mediante la comparación de medias para las combinaciones de los niveles de todos los factores, se realizó mediante la prueba de comparaciones múltiples LSD de Fisher (a = 0,05). Se realizó un análisis de correlación múltiple de Pearson entre las variables estudiadas con un nivel del significancia del 5 %. Todas las pruebas estadísticas se corrieron en el programa InfoStat versión 2013 (Di Rienzo et al. 2013).
Resultados
Efectos de los factores estudiados
Se observó un baja incidencia de E. ello (< 5 %) y de L. heveae (< 7 %) en las dos zonas, en los tres clones de caucho y en los cuatro sistemas de siembra (Tabla 3). La época pluviométrica fue el factor más influyente, especialmente en L. heveae, que estuvo ausente en las tres primeras épocas y alcanzó la mayor incidencia (25,63 %) en la segunda época lluviosa (Tabla 3). Altas correlaciones significativas se encontraron entre la incidencia y la abundancia de E. ello y de L. heveae (ambas, r ≥ 0,90; gl = 431; P < 0,01).
Medias de la incidencia (%) y abundancia de gusano cachón (Erinnyis ello) y chinche de encaje (Leptopharsa heveae) en tres clones de caucho (Hevea brasiliensis) establecidos en diferentes sistemas de siembra y evaluados en diferentes épocas en dos zonas de Caquetá (Colombia).
Error estándar.
Valores en cada columna seguidos por la misma letra no difieren estadísticamente (Prueba LSD, P < 0,05).
L. heveae, expresada en número de insectos por foliolo por planta. E. ello, expresada en número de insectos por 100 foliolos por planta.
Se encontraron diferencias significativas entre Albania y San Vicente del Caguán en las incidencias de E. ello (F = 31,69; gl = 1; P < 0,01) y de L. heveae (F = 17,93; gl = 1; P < 0,05) (Tabla 3). Asimismo, hubo un efecto muy significativo de la época en la incidencia de E. ello (F = 8,89; gl = 1; P < 0,01) y de L. heveae (F = 355,51; gl = 5; P < 0,01). No se observaron diferencias significativas entre clones ni entre sistemas de siembras en la incidencia de ambos insectos (P > 0,05).
La prueba de hipótesis para la interacción zona x época x sistema resultó significativa para la incidencia de E. ello (F = 2,05; gl = 15; P < 0,05) (Fig. 1). Esto indica que para una zona dada, las diferencias observadas en la incidencia de E. ello entre los cuatro sistemas de siembra, fueron a su vez influenciadas por la época pluviométrica en la que se realizaron las evaluaciones. Del mismo modo, fueron significativas las interacciones zona x época (F = 40,75; gl = 5; P < 0,01) y época x sistema (F = 6,86; gl = 15; P < 0,01) para la incidencia de L. heveae (Figs. 2A y B). Esto confirma la influencia de la época pluviométrica sobre las diferencias observadas en la incidencia del chinche de encaje entre zonas y entre sistemas de siembra.
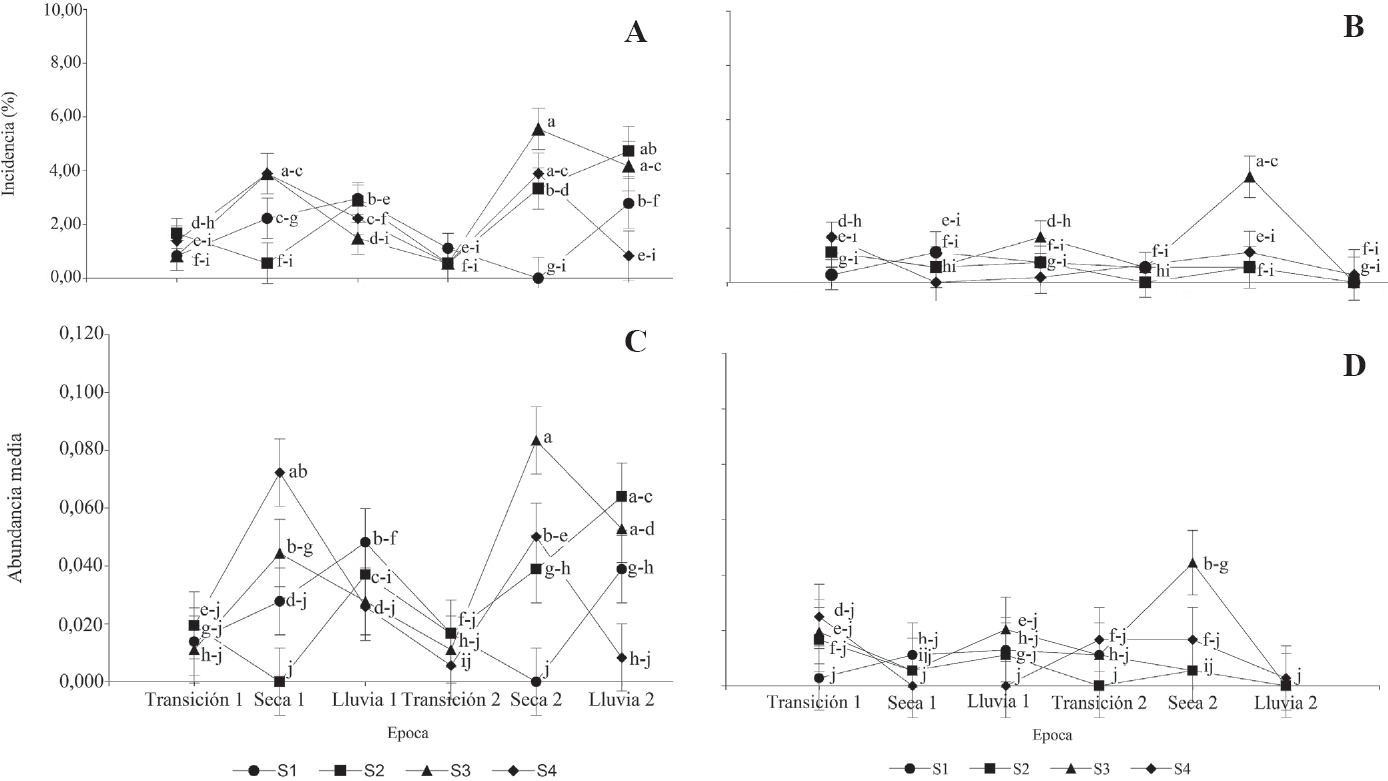
Medias de la incidencia (%) y abundancia (número de insectos por 100 foliolos por planta) de gusano cachón (Erinnyis ello) en diferentes sistemas de siembra (S): [Hevea brasiliensis (S1: 2,5 x 3,0 x 14,0 m; S2: 2,5 x 3,0 x 13,0 m; S3: 2,8 x 4,0 x 10,0 m; S4: 2,8 x 7,0 m); Theobroma grandiflorum y Musa AAB: 4,0 x 5,0 m para los sistemas S1, S2 y S3] para seis épocas pluviométricas en dos zonas de Caquetá (Colombia). A y C. Albania, B y D. San Vicente del Caguán. Medias seguidas por la misma letra en cada época no difieren estadísticamente (Prueba LSD; P < 0,05). Las barras representan el error estándar de la media.
En relación con la abundancia, se encontraron diferencias significativas entre las dos zonas para E. ello (F = 36,86; gl = 1; P < 0,01) y L. heveae (F = 9,07; gl = 1; P < 0,05) (Tabla 3). Sólo se observaron diferencias significativas entre clones de caucho en la abundancia media de L. heveae (F = 5,94; gl = 2; P < 0,01). Se observó además un efecto muy significativo de la época en la abundancia de los dos insectos (ambos, P < 0,01). No se observaron diferencias significativas entre sistemas de siembras en la abundancia de las dos plagas (ambas, P > 0,05).
El análisis de la interacción zona x época x sistema mostró significancia para la abundancia media de E. ello (F = 1,83; gl = 15; P < 0,05) (Figs. 1C y D) y de L. heveae (F = 2,47; gl = 15; P < 0,01) (Figs. 2C y D). Esto significa que la abundancia de ambos insectos fue influenciada en cada sistema de siembra por la zona en la que se establecieron los clones de caucho y por las condiciones pluviométricas (época) en las que se realizaron las evaluaciones. Para el L. heveae se observó además una interacción significativa entre épocas y clones de caucho (F = 5,31; gl = 10; P < 0,01). En este sentido, las diferencias observadas en la abundancia de este insecto entre clones no fue la misma entre las seis épocas pluviométricas estudiadas.
Incidencia y abundancia de E. ello
Se observó una incidencia media muy baja de E. ello, la cual no fue superior al 2 % en ninguno de los clones estudiados (Tabla 3). Del mismo modo, la abundancia de E. ello presentó un valor medio muy bajo (0,021 insectos por cada 100 foliolos por planta), donde el clon más tolerante fue el FDR 5788 (Tabla 3).
En relación con las zonas, la menor incidencia y abundancia de E. ello se observó en San Vicente del Caguán (Tabla 3 y Fig. 1). Sin embargo, la mayores medias de incidencia (< 6 %) y abundancia (< 1 insecto por cada 100 foliolos por planta) en ambas zonas, se presentaron en el sistema de siembra S3, especialmente durante la segunda época seca (Figs. 1A y B).
De acuerdo con la Figura 1, en Albania, durante las dos transiciones hacia la época seca se observaron las menores medias de incidencia (≤ 1,7 %) y abundancia (≤ 0,08 insectos por cada 100 foliolos por planta) de E. ello en los cuatro sistemas de siembra. Por su parte, en San Vicente del Caguán los menores registros de incidencia (≤ 1,7 %) y abundancia (≤ 0,04 insectos por cada 100 foliolos por planta) se encontraron durante la segunda época lluviosa (Figs. 1B y D). En general, para ambas zonas y en las diferentes épocas, el sistema de siembra S1 fue el más favorable para H. brasiliensis frente al ataque de E. ello.
Incidencia y abundancia de L. heveae
Se registró una baja incidencia de L. heveae en los tres clones de caucho, la cual no fue superior al 5 % en los tres clones estudiados (Tabla 3). Asimismo, se observó una abundancia media muy baja (< 1 insecto por foliolo por planta), donde el clon FDR 5788 fue el más tolerante al ataque del insecto (Tabla 3).
En Albania se registraron las menores medias de incidencia y abundancia de L. heveae, las cuales fueron significativamente diferentes de lo observado en San Vicente del Caguán (Tabla 3). Esto se debió principalmente a la mayor incidencia (34,17 %) presentada por L. heveae en la segunda época lluviosa de San Vicente del Caguán, ya que en las demás épocas pluviométricas no hubo diferencias significativas en la incidencia de L. heveae entre ambas zonas (Fig. 2A).
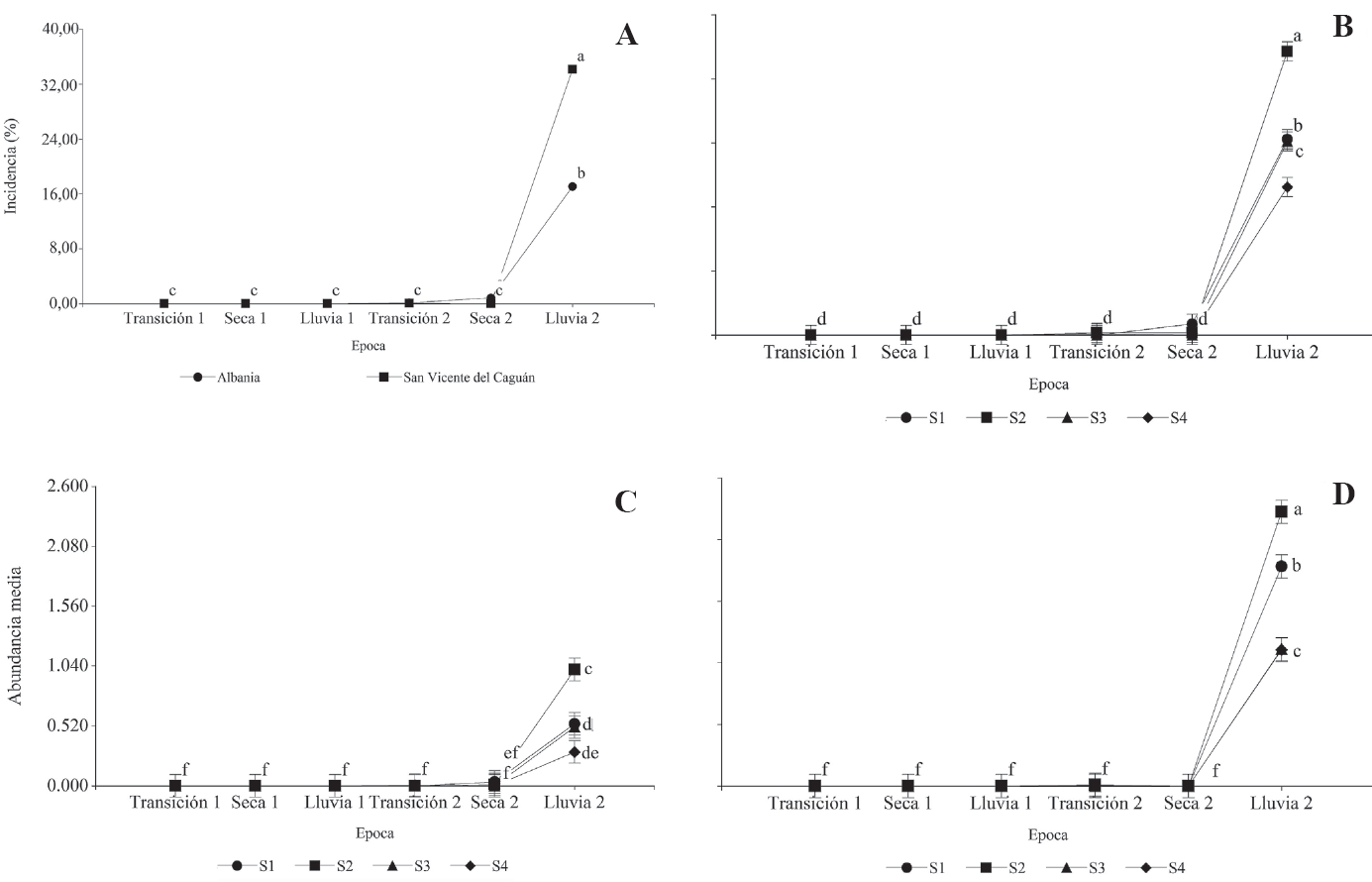
Medias de la incidencia (%) y abundancia (número de insectos por foliolo por planta) de chinche de encaje (Leptopharsa heveae) en caucho (Hevea brasiliensis) en Caquetá (Colombia). A. Interacción localidad x época. B. Interacción sistema x época. C y D. Interacción época x sistema x zona: (C, Albania) y (D, San Vicente del Caguán). Clave: Sistemas de siembra (S): [Hevea brasiliensis (S1: 2,5 x 3,0 x 14,0 m; S2: 2,5 x 3,0 x 13,0 m; S3: 2,8 x 4,0 x 10,0 m; S4: 2,8 x 7,0 m); Theobroma grandiflorum y Musa AAB: 4,0 x 5,0 m para los sistemas S1, S2 y S3]. Medias seguidas por la misma letra en cada época no difieren estadísticamente (Prueba LSD; P < 0,05). Las barras representan el error estándar de la media.
En general, durante la segunda época lluviosa se presentó la mayor incidencia media de L. heveae en ambas zonas y en los cuatro sistemas se siembra (ambos, < 36 %) (Figs. 2A y B). El sistema de siembra S4 fue el que presentó menor incidencia del insecto. Asimismo, en Albania como en San Vicente del Caguán, hubo ausencia de L. heveae en cuatro de las seis épocas evaluadas.
Las mayores medias de abundancia de L. heveae entre clones (< 2 insectos por foliolo por planta) y entre sistemas de siembra (< 3 insectos por foliolo por planta) se registraron también en la segunda época lluviosa en ambas zonas de estudio (Figs. 2C y D). El clon FDR 5788 fue el más tolerante (< 1 insecto por foliolo por planta), y los sistemas de siembra S4 y S3, los más favorables para H. brasiliensis frente al ataque de L. heveae (< 2 insectos por foliolo por planta).
Discusión
El uso de clones de caucho tolerantes a las principales enfermedades y plagas del cultivo (Gonçalves et al. 1997) y la implementación de prácticas agroforestales (Pereira et al. 1997) representan dos de las estrategias más prometedoras para el manejo fitosanitario de H. brasiliensis en regiones con alta incidencia de patógenos bióticos y artrópodos plagas (Garzón 2000).
Los resultados de este estudio confirman lo enunciado anteriormente, dado que se evidenciaron abundancias medias muy bajas de E. ello y de L. heveae (ambos, < 2 insectos por foliolo) en los clones promisorios de caucho y en los nuevos sistemas de siembra probados, contrario a lo informado en otras investigaciones donde el nivel de infestación ha sido de 14,4 insectos por foliolo en clones comerciales de caucho en países como Brasil (Da Silva 2002). En Colombia, por ejemplo se han reportado infestaciones por E. ello superiores al 15 % (Sterling et al. 2009) y al 50 % (Garzón 2000), valores que difieren en gran medida con lo informado en el presente estudio (< 5 %).
Aunque en esta investigación no se observaron diferencias significativas en las incidencias de E. ello y L. heveae entre clones, autores como Lara y Tanzini (1997) encontraron diferentes niveles de susceptibilidad clonal a L. heveae, donde el clon IAN 873 fue uno de los más susceptibles. No obstante, acorde con el presente estudio, los resultados de otras investigaciones realizadas sobre plantaciones en monocultivo de caucho en condiciones de la Amazonia colombiana, confirman la ausencia de una reacción clonal diferencial frente a la probabilidad de ataque en campo de ambos insectos plaga sobre el cultivo del caucho (Sterling et al. 2012).
En el caso de E. ello, se ha informado también sobre la reacción clonal diferencial del caucho al ataque de éste insecto. En este sentido, Sterling et al. (2011) en un campo clonal a pequeña escala de genotipos élite de caucho en Caquetá, encontraron diferencias significativas en la incidencia de E. ello entre éstos materiales y el testigo IAN 873, contrario a lo informado en el presente estudio, donde no se observaron diferencias estadísticamente significativas entre los tres clones de caucho analizados.
De acuerdo con la escala de Fonseca (2001), el intervalo de abundancia observado en este estudio para L. heveae, fue bajo, debido a que no superó los dos insectos por foliolo. En el mismo sentido y con la escala de Fonseca (2009), la abundancia de E. ello se clasificó como media, dado que no superó los cincos insectos por cada 100 foliolos. Esto evidencia una reacción favorable de los clones evaluados en este estudio ante la infestación de estos dos insectos plagas, dada la tolerancia intrínseca atribuida a cada clon (Lara y Tanzini 1997) y en gran manera a los sistemas de siembra en los que fueron establecidos. En este sentido, Gol dab et al. (1990) afirman que E. ello es menos abundante en cultivos asociados que en monocultivo, similar a lo observado en éste estudio especialmente en el sistema de siembra S1.
Aunque no se observaron diferencias significativas en las incidencias de E. ello y L. heveae entre los cuatro sistemas de siembra, en general, se encontraron bajos niveles de infestación en los sistemas agroforestales S1 para E. ello, y S3 para L. heveae. Al respecto, Santos et al. (2012) señalan que las plantaciones de caucho establecidas masivamente en monocultivo son más susceptibles el ataque de las plagas, por lo que la alternativa de siembra en arreglos agroforestales resulta promisoria (Alvin et al. 1989; Pereira et al. 1997). Asimismo, Junqueira et al. (1999) afirman además, que la arquitectura de los árboles dentro de los sistemas de cultivo, propician condiciones de humedad y luminosidad favorables para la sobrevivencia y diseminación de los enemigos naturales, y por tanto, contribuyen con la biorregulación de plagas como L. heveae al interior del cultivo.
Por otro lado, es importante comprender que las características ambientales propias de cada zona son preponderantes en la probabilidad de aparición y ataque de los insectos hacia sus hospederos (Campbell et al. (1974). En la presente investigación la variación espacial no afectó la incidencia de E. ello, pero si hubo un efecto significativo sobre L. heveae, contrario a lo informado por Sterling et al. (2012) quienes no encontraron diferencias significativas en tres campos clonales de caucho en condiciones de la Amazonia colombiana. Las diferencias entre zonas, informadas en el presente estudio para L. heveae podrían explicarse a partir de las características térmicas propias de cada región. Al respecto, Campbell et al. (1974) y Higley et al. (1986) afirman que la temperatura es considerada el factor más importante en el desarrollo y la reproducción de los insectos.
Cividanes et al. (2004) afirman que en regiones más cálidas con el aumento de la temperatura (20 a 30 °C), el tiempo de incubación de las posturas es menor, la supervivencia de los huevos es mayor y los estadios ninfales se adaptan más rápido, lo que favorece la mayor probabilidad de aparición y diseminación del insecto. Al respecto, en la presente investigación, la mayor incidencia (5,71 %) de L. heveae se registró en la zona más cálida (25,4 °C) que en éste caso correspondió a San Vicente del Caguán. No obstante, lo anterior difiere para E. ello, debido a que la mayor incidencia y abundancia se presentó en la zona menos cálida, es decir, en Albania. Esto coincide con Sterling et al. (2012) quienes registraron las menores incidencias (< 1%) de E. ello en las zonas con menores valores de temperatura.
Aunado a la variación espacial, las variaciones microclimáticas propias de cada zona juegan un rol muy importante en la probabilidad de aparición y diseminación de los insectos plagas (Higley et al. 1986). En el presente estudio, las condiciones microclimáticas registradas en cada época pluviométrica tuvieron un efecto significativo sobre las incidencias de E. ello y L. heveae similar a lo informado por Sterling et al. (2011) en un campo clonal a pequeña escala con materiales regionales de caucho en condiciones de la Amazonia colombiana. No obstante, las mayores incidencias de L. heveae se registraron en épocas contrastantes, comparativamente entre ambos estudios. De acuerdo con lo anterior, en el presente estudio, la mayor incidencia de L. heveae se observó en la época lluviosa de 2014 (lluvia 2) tanto en Albania como en San Vicente del Caguán, contrario a lo informado por dichos autores, quienes registraron los índices poblacionales más altos en la primera época seca de 2010.
No obstante, Santos y Freitas (2008) en el estado de Mato Grosso en Brasil, soportan también la idea de que el período de lluvias favorece la aparición de L. heveae, debido a que sus resultados permitieron concluir que el comienzo de la temporada de lluvias se correlaciona positivamente con el aumento en la población L. heveae, similar a lo informado en el presente estudio. Asimismo, Santos et al. (2012) afirman que en el inicio de la temporada de lluvias se produce un aumento en el número de huevos de L. heveae así como un incremento de nitrógeno soluble en los tejidos de las hojas de caucho, lo que aumenta su palatabilidad y por tanto, se incrementa la probabilidad de ataque de éste insecto, lo que conlleva a su vez a la formación de picos poblacionales durante la época lluviosa.
Respecto a E. ello, la mayor incidencia informada en este estudio coincide con Sterling et al. (2011) para la época seca, seguida de la época lluviosa y en menor grado durante los periodos de transición hacia la seca. Sin embargo, los valores medios (< 23 %) entre épocas encontrados por estos autores, son superiores a lo informado en la presente investigación (< 3 %). Bellotti et al. (1999) y Martínez y García (2007) afirman que los daños producidos por E. ello en la época seca son mayores que los observados en la lluviosa. Asimismo, León et al. (2010) afirman que los daños causados por esta especie pueden llegar al 100 % de defoliación, especialmente al final de la época seca, después del ciclo natural de defoliación - refoliación del caucho, debido a la disponibilidad de foliolos inmaduros altamente palatables para los estadios larvales de éste insecto.
Conclusiones
Los resultados del estudio permiten concluir que las incidencias y abundancias de E. ello y L. heveae en H. brasiliensis, están influenciadas tanto por la variación espacial (zona) y temporal (época pluviométrica) como por el sistema de siembra implementado, en este último caso, según la época pluviométrica en la que hayan aparecido estos insectos. En el mismo sentido, existe una variación temporal en la reacción clonal a L. heveae, en donde el clon FDR 5788 en esta investigación fue el más tolerante en los cuatro sistemas de siembra evaluados, especialmente en las épocas con menores niveles de precipitación.
Este estudio es el primero en el que se analiza desde un enfoque agroforestal, los cambios en la infestación producida por E. ello y L. heveae en clones promisorios de H. brasiliensis en condiciones de la Amazonia colombiana.
Agradecimientos
Los autores agradecen: a) al proyecto "Evaluación de clones promisorios de caucho natural (Hevea brasiliensis) para la Amazonia colombiana bajo sistema agroforestal con copoazú (Theobroma grandiflorum) y plátano hartón (Musa AAB) en el departamento de Caquetá" – Contrato RC No. 558 -2011- Fiduciara Bogotá – Colciencias – Sinchi –Asoheca y al convenio marco No. 342/2006 de cooperación interinstitucional de investigación en la cadena productiva del caucho natural entre la Universidad de la Amazonia y Asoheca, por el desarrollo de la presente investigación; b) a Jean Alexander Gamboa, Clemencia Serrato y Jaime Enrique Velásquez por sus importantes comentarios en la realización de este estudio.