Spodoptera cosmioides (Lepidoptera: Noctuidae) is a polyphagous pest that causes damage to various crops including those used in the production of biodiesel, such as castor bean (Ricinus communis), Barbados nut (Jatropha curcas), and Tung-oil tree (Aleurites fordii). One of the main biological control agents of lepidopteran pests are egg parasitoids of the genus Trichogramma. The influence of temperature on the performance of three strains (TRS13, TRS65 and TRS81) of Trichogramma pretiosum (Hymenoptera: Trichogrammatidae) on eggs of S. cosmioides and thermal requirements were studied. The experiments were carried out at constant temperatures (18, 20, 22, 25 and 28) and 30 °C, 70 ± 10% relative humidity and photoperiod of 14:10 (light: dark) hours. The development time of the three strains of T. pretiosum was inversely related to temperature rise. The temperatures affected the parasitoid emergency, being higher than 74% for each of the strains at 25 °C. However, the temperatures tested did not affect the number of adults per egg and sex ratio. The lower temperature threshold (Tb) and thermal constant (K) were 11.21 °C and 131.41 DD (degree-days) for strain TRS13, 10.73 °C and 137.70 DD for TRS65 and 10.68 °C and 138.70 DD for TRS81.
Cultivos con potencial agro-energético como el ricino (Ricinus communis L.) (Euphorbiaceae), piñón (Jatropha curcas L.) (Euphorbiaceae) y tung (Aleurites fordii Hemsl) (Euphorbiaceae), han despertado el interés del gobierno brasilero para viabilizar la producción de aceites vegetales y biodiesel (Suarez et al. 2006; Leite y Leal 2007; Pousa et al. 2007; Oliveira et al. 2009). Sin embargo, faltan informaciones sobre el manejo de insectos-plaga y de enfermedades en estos cultivos (Shanker y Dhyani 2006; Franco y Gabriel 2008).
Entre los insectos, Spodoptera cosmioides (Walker, 1858) (Lepidoptera: Noctuidae) es considerada una importante plaga del ricino (Carvalho 2005), siendo también encontrada en cultivos de piñón y tung (D. E. Nava, com. pers. 2012). Su presencia se restringe a América del Sur, con excepción del sur de Argentina, Chile y de regiones de Perú situadas al oeste de Los Andes (Silvain y Lalanne-Cassou 1997). Los registros sobre este insecto son escasos; considerado sinonimia de S. latifascia (Walker, 1856) (Lepidoptera: Noctuidae), hasta que Silvain y Lalanne-Cassou (1997) describieran las diferencias moleculares, morfológicas, fisiológicas y de comportamiento que las separan. Así, los estudios realizados en América del Sur hasta 1997 con S. latifascia, especie presente en América Central, se refieren a S. cosmioides (Araújo 2009). Esta especie es polífaga y ataca varias plantas cultivadas y malezas. En Brasil, más de 24 cultivos de interés económico son citados como sus hospederos (Habib et al. 1983; Bavaresco et al. 2001, 2002, 2003, 2004; Pastrana 2004; Specht et al. 2004). Sin embargo, a pesar de su amplia gama de hospederos, su importancia como plaga es poco conocida en algunos cultivos, probablemente debido al uso excesivo de insecticidas de amplio espectro (Araújo 2009). Entre los agentes de control biológico de S. cosmioides se destacan los parasitoides de huevos del género Trichogramma, que constituyen uno de los grupos de enemigos naturales más estudiados en el mundo (van Lenteren 2003). Estos microhimenópteros parasitan exclusivamente huevos de lepidópteros (Hassan 1997; Zucchi et al. 2010) e impiden que el insecto llegue a la fase larval y que, consecuentemente, cause daños. El empleo de Trichogramma en programas de control biológico de plagas es realizado en cerca de 16 millones de hectáreas entre cultivos anuales y perennes en todo el mundo (Parra y Zucchi 2004).
El desarrollo, calidad y desempeño de Trichogramma son influenciados por factores bióticos y abióticos (Hassan 1997). La temperatura es el factor abiótico de mayor importancia, interfiere en el ciclo de desarrollo, proporción sexual, supervivencia, desempeño y fecundidad (Stein y Parra 1987; Cônsoli y Parra 1996; Pratissoli y Parra 2000; Molina et al. 2005).
El objetivo de este trabajo fue evaluar la influencia de la temperatura en el desempeño de T. pretiosum Riley, 1879 (Hymenoptera: Trichogrammatidae) sobre huevos de S. cosmioides, así como determinar el límite térmico inferior de desarrollo y la constante térmica.
Materiales y métodos
El trabajo se realizó en el Laboratorio de Entomología del Centro de Pesquisa Agropecuária de Clima Temperado (CPACT) de la EMBRAPA (Empresa Brasileira de Pesquisa Agropecuária), Pelotas, Rio Grande do Sul, Brasil.
Colonia de mantenimiento de S. cosmioides
La colonia de mantenimiento se inició con colectas de huevos en cultivos de ricino localizados en el área experimental del CPACT de EMBRAPA. Las larvas neonatas fueron colocadas en tubos de vidrios de fondo plano (8,5 cm de altura x 2,7 cm de diámetro) con dieta artificial a base de germen de trigo, frijol y levadura de cerveza (Greene et al. 1976), y criadas según el método propuesto por Parra (2009). Después de la pupación, los insectos se separaron por sexo y se colocaron en cajas Gerbox ® (11,5 cm x 11,0 cm x 3,5 cm) con vermiculita húmeda, hasta la emergencia de los adultos. Parejas de S. cosmioides, de hasta 24 horas, se colocaron en jaulas formadas por tubos de PVC (13,0 cm de altura x 10,0 cm diámetro), internamente revestidas con papel, que sirvió como substrato de oviposición. Los adultos se alimentaron con una solución de miel al 10%, ofrecida por capilaridad en pedazos de algodón colocados en recipientes plásticos y cambiados cada dos días. En las extremidades de los tubos de PVC se colocaron placas de acrílico para evitar la fuga de los adultos. El papel con las posturas se retiró y cambió diariamente. Los insectos se mantuvieron en salas climatizadas a 25 ± 1 °C de temperatura, humedad relativa de 70 ± 10% y fotoperiodo de 14:10 h L:O.
Cría de las cepas de T. pretiosum
Se utilizaron tres cepas de T. pretiosum (TRS13, TRS65 e TRS81), mantenidas en laboratorio, cuyas colectas se realizaron en varios hospederos de diferentes localidades del estado de Río Grande do Sul. Los parasitoides se criaron en huevos del hospedero alternativo Anagasta kuehniella (Zeller, 1879) (Lepidoptera: Pyralidae), mantenido en dieta artificial a base de harina de trigo integral (97%) y levadura de cerveza (3%), según la metodología descrita por Parra (1997). Las cepas de T. pretiosum se mantuvieron en tubos de vidrio (8,5 cm de altura x 2,5 cm de diámetro) tapados con una película plástica de PVC Magipack®, siendo los adultos alimentados con una pequeña gota de miel, colocada en la parte interna del tubo. Los huevos de A. kuehniella se adhirieron a un rectángulo de cartulina azul (8,0 cm de largo x 2,0 cm de ancho), con goma arábiga diluida en agua al 50% y se esterilizaron con luz germicida durante 45 minutos, a una distancia de 15 cm de la fuente de luz (Stein y Parra 1987). En las extremidades de la cartulina se anotó la fecha de parasitismo y el código de identificación de las cepas, para facilitar el control. Después de inviabilizados, los huevos se expusieron al parasitismo durante 24 horas, siendo posteriormente retirados y colocados dentro de tubos con las mismas dimensiones descritas anteriormente y almacenados en cámara climatizada regulada a 25 ± 1 °C de temperatura, humedad relativa de 70 ± 10% y fotoperiodo de 12:12 h L:O.
Efecto de la temperatura sobre el parasitismo
Quince hembras de cada cepa con 24 horas de edad se individualizaron en tubos de vidrio (7,3 cm de altura x 1,0 cm de diámetro) tapados con Magipack® y se alimentaron con miel pura. Para cada hembra se colocaron 30 huevos de S. cosmioides con una edad máxima de 24 horas de desarrollo embrionario. Estos huevos se expusieron a parasitismo por 24 horas, en cámaras climatizadas a 25 ± 1 °C de temperatura, humedad relativa de 70 ± 10% y fotoperiodo de 12:12 h L:O. Después de este período, las hembras se retiraron de los tubos y éstos, conteniendo los huevos, se mantuvieron en cámaras climatizadas con temperaturas de 18, 20, 22, 25, 28 y 30 ± 1 °C, humedad relativa de 70 ± 10% y fotoperiodo de 12:12 h L:O. Se evaluaron los siguientes parámetros biológicos: duración del período huevo-adulto, porcentaje de emergencia, número de adultos emergidos por huevo y proporción sexual (hembras/(hembras + machos)). Con los datos de duración del período huevo-adulto se determinaron las exigencias térmicas, utilizando el método de la hipérbole (Haddad et al. 1999).
Análisis estadístico
Se utilizó un diseño estadístico totalmente al azar con arreglo factorial (3 cepas x 6 temperaturas). Los resultados de los parámetros biológicos observados se sometieron a análisis de varianza, usando el procedimiento PROC GLM del SAS (SAS Institute 2002) y las medias de los tratamientos se compararon por la prueba t de Student (P < 0,05).
Resultados y discusión
Para el período huevo-adulto no se observaron diferencias significativas entre las cepas (F = 0,19; df = 2, 234; P = 0,83), pero sí para temperatura (F = 3140,37; gl = 5, 234; P < 0,01) e interacción de cepas x temperatura (F = 14,10; gl = 10, 234; P < 0,01), variando de 6,29 días a 30 °C a 18,83 días a 18 °C. Entre las cepas, el período medio varió de 11,62 a 11,83 días (Tabla 1). La disminución de la duración del período de desarrollo huevo-adulto de las cepas de T. pretiosum, con el aumento de la temperatura, puede ser consecuencia del incremento de la actividad metabólica de los parasitoides (Bueno et al. 2009). Comportamientos similares para T. pretiosum en diferentes hospederos fueron registrados por Pratissoli y Parra (2000), Molina et al. (2005), Pratissoli et al. (2005b) y Bueno et al. (2009).
El porcentaje de emergencia tambien fue altamente afectado por la temperatura en las condiciones de laboratorio estudiadas, observando un efecto significativo entre las cepas (F = 4,70; gl = 2, 234; P = 0,01), temperatura (F = 8,84; gl = 5, 234; P < 0,01) e interacción de cepas x temperatura (F = 2,28; gl = 10, 234; P = 0,02) (Tabla 1). Para cada una de las cepas evaluadas, los mayores porcentajes de emergencia se obtuvieron a 25 °C, variando de 74,95 a 77,45%.
Parámetros biológicos de cepas de Trichogramma pretiosum criadas en huevos de Spodoptera cosmioides a diferentes temperaturas. HR: 70 ± 10%; fotoperiodo: 12:12 h L:O.
Medias seguidas de la misma letra minúscula en la línea y letra mayúscula en la columna no difieren entre sí por la prueba t (P < 0,05).
Con relación al número de adultos por huevo, no se observaron diferencias significativas entre las cepas (F = 0,05; gl = 2, 146; P = 0,95), temperatura (F = 0,83; gl = 5,146; P = 0,53) e interación de cepas x temperatura (F =1,36; gl = 10,146; P = 0,51) (Tabla 1). De igual forma, la temperatura no afectó la proporción sexual entre cepas (F = 1,76; gl = 2, 146; P = 0,18), temperatura (F = 1,92; gl = 5,146; P = 0,10) e interacción de cepas x temperatura (F =1,23; gl = 10,146; P = 0,28) (Tabla 2). Molina et al. (2005) y Pratissoli et al. (2005b) no encontraron diferencias significativas del efecto de la temperatura sobre estos mismos parámetros en T. pretiosum. Sin embargo, Pratissoli y Parra (2000) observaron diferencias en la proporción sexual de T. pretiosum en función de la temperatura, en huevos de Phthorimaea operculella (Zeller, 1873) (Lepidoptera: Gelechiidae). Por otro lado, Bueno et al. (2009) observaron que la proporción sexual de T. pretiosum en huevos de Anticarsia gemmatalis (Hübner, 1818) (Lepidoptera: Noctuidae) y de Pseudoplusia includens (Walker) es afectada por la temperatura y también por el hospedero.
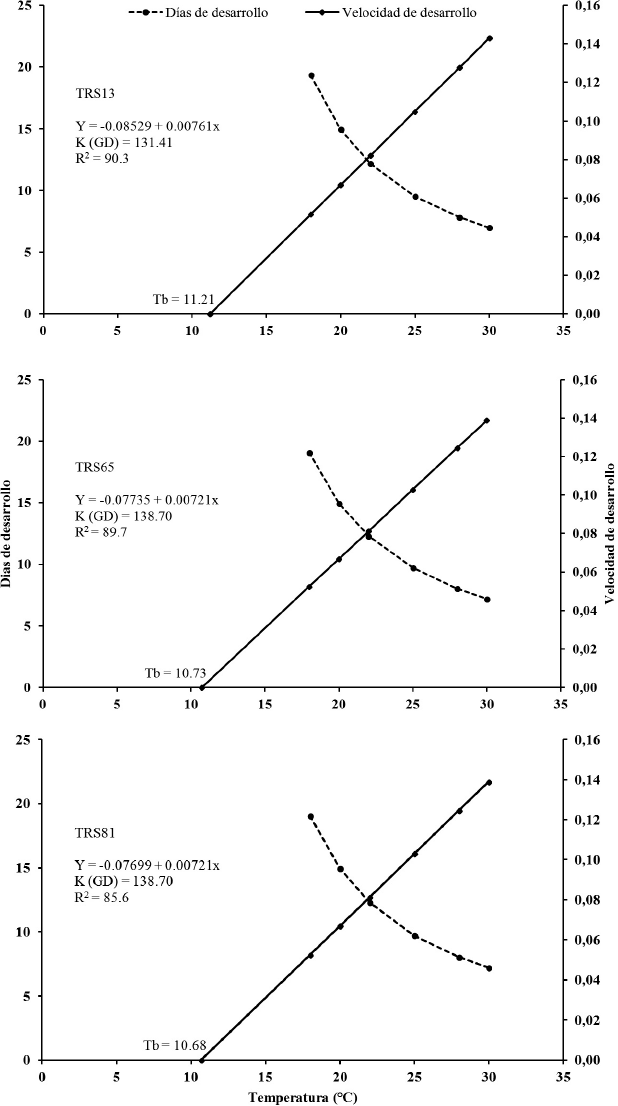
Los valores del límite térmico inferior de desarrollo (Tb) para las cepas de T. pretiosum TRS13 (11,21 °C), TRS65 (10,73 °C) y TRS81 (10,68 °C) (Fig. 1) son similares a los encontrados con otros hospederos (Pratissoli et al. 2005a; Bueno et al. 2009). La constante térmica (K) observada en este estudio para las tres cepas de T. pretiosum (131,41 para TRS13 y 138,70 GD para TRS65 y TRS81) (Fig. 1) en huevos de S. cosmioides son superiores a las registradas para esta misma especie, cuando criada en huevos de Cydia pomonella (Linnaeus, 1758) (Lepidoptera: Tortricidae) (130,7 GD), Anagasta kuehniella (Zeller, 1879) (Lepidoptera: Pyralidae), (128,5 GD), P. operculella (120,9 GD), Tuta absoluta (Lepidoptera: Gelechiidae) (131,3 GD), Ecdytolopha aurantiana (Lima, 1927) (Lepidoptera: Tortricidae) (107,8 GD) y A. gemmatalis (127,6 GD) (Yu et al. 1984; Pratissoli y Parra 2000; Molina et al. 2005; Bueno et al. 2009). No obstante, los grados-días de las cepas evaluadas en este estudio son inferiores a los valores reportados para T. pretiosum en huevos de Heliothis virences (164,6 GD) y Galleria mellonella (183,3 GD) (Goodenough et al. 1983), A. kuehniella (151,8 GD), Sitotroga cerealella (160,0 GD) (Pratissoli et al. 2005a) y P. includens (151,3 GD) (Bueno et al. 2009). Las variaciones entre los valores de temperatura base y constante térmica observados en el presente trabajo y los registrados por los autores anteriormente citados, indican la influencia de la cepa y el hospedero sobre la biología de T. pretiosum. Estas variaciones también pueden estar asociadas al comportamiento de búsqueda, preferencia por el hospedero y respuestas a las condiciones ambientales (Hassan 1997). Pratissoli et al. (2005a) sostienen que la constante térmica (K) con base en la duración del período de desarrollo del parasitoide en diferentes temperaturas, a partir de un límite inferior de temperatura (Tb), está relacionada con las necesidades térmicas del insecto. Los resultados presentados muestran que la temperatura es uno de los factores abióticos que afecta el desarrollo de las cepas de T. pretiosum.
Proporción sexual de cepas de Trichogramma pretiosum criadas en huevos de Spodoptera cosmioides a diferentes temperaturas. HR: 70 ± 10%; fotoperiodo: 12:12 h L:O.
Medias seguidas de la misma letra minúscula en la línea y letra mayúscula en la columna no difieren entre sí por la prueba t (P < 0,05).
Relación entre la velocidad de desarrollo y la temperatura, para el período huevo-adulto de las cepas TRS13, TRS65 y TRS81 de Trichogramma pretiosum, criadas en huevos de Spodoptera cosmioides, en diferentes temperaturas. HR: 70 ± 10%; fotoperiodo: 12:12 h L:O; Tb = Límite térmico inferior de desarrollo, K =Constante térmica y GD = Grados día.
Conclusiones
Los parámetros biológicos y los requerimientos térmicos de T. pretiosum observados sobre huevos de S. cosmioides aportan informaciones importantes para el desarrollo de estrategias de manejo integrado de este insecto-plaga en los cultivos de ricino, tung y piñon. Sin embargo, estudios sobre el desarrollo de este parasitoide en campo son necesarios, debido a que las condiciones de temperatura durante el día son variables. Parámetros como potencial de búsqueda por el hospedero, capacidad de dispersión, estrategias e intervalos de liberación, son factores importantes que de no ser considerados podrían afectar el buen desempeño de T. pretiosum como agente de control biológico de S. cosmioides en campo.
Footnotes
Agradecimiento
A FINEP (Financiadora de Estudos e Projetos), CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico) por el financiamiento de parte de los estudios.
References
1.
ARAÚJOC. R.2009. Aspectos biológicos de Spodoptera cosmioides Walker, 1858 (Lepidoptera: Noctuidae) nas cultivares de algodoeiro DeltaOPAL e NuOPAL (Bollgard I). Tesis de Maestría en Agronomía, Universidade Estadual Paulista.
2.
BAVARESCOA.GARCIAM. S.GRÜTZMACHERA. D.FORESTIJ.RINGENBERGR.2001. Efeito de fontes de carboidratos sobre o desempenho reprodutivo de Spodoptera cosmioides (Walk., 1858) (Lepidoptera: Noctuidade). Revista Brasileira de Agrociência7 (3): 177–180.
3.
BAVARESCOA.GARCIAM. S.GRÜTZMACHERA. D.FORESTIJ.RINGENBERGR.2002. Biologia e exigências térmicas de Spodoptera cosmioides (Walk.) (Lepidoptera: Noctuidae). Neotropical Entomology31 (1): 49–54.
4.
BAVARESCOA.GARCIAM. S.GRÜTZMACHERA. D.FORESTIJ.RINGENBERGR.2003. Biologia comparada de Spodoptera cosmioides (Walk.) (Lepidoptera: Noctuidae) em cebola, mamona, soja e feijão. Ciência Rural33 (6): 993–998.
5.
BAVARESCOA.GARCIAM. S.GRÜTZMACHERA. D.RINGENBERGR.FORESTIJ.2004. Adequação de uma dieta artificial para a criação de Spodoptera cosmioides (Walk.) (Lepidoptera: Noctuidae) em laboratório. Neotropical Entomology33 (2): 155–161.
6.
BUENOR. C. O. F.PARRAJ. R. P.BUENOA. D. F.2009. Biological characteristics and thermal requirements of a Brazilian strain of the parasitoid Trichogramma pretiosum reared on eggs of Pseudoplusia includens and Anticarsia gemmatalis. Biological Control51 (3): 355–361.
7.
CARVALHOB. C. L.2005. Manual do cultivo da mamona. EBDA, Salvador. 65 p.
8.
CÔNSOLIF. L.PARRAJ. R. P.1996. Biology of Trichogramma galloi and T. pretiosum (Hymenoptera: Trichogrammatidae) reared in vitro and in vivo. Annals of the Entomological Society of America89 (6): 828–834.
9.
FRANCOD. A. S.GABRIELD.2008. Aspectos fitossanitários na cultura do pinhão manso (Jatropha curcas L.) para produção de biodiésel. Biológico70 (2): 64–64.
10.
GOODENOUGHJ. L.HARTSTACKA. W.KINGE. G.1983. Developmental models for Trichogramma pretiosum (Hymenoptera: Trichogrammatidae) reared on four hosts. Journal of Economic Entomology76 (5): 1095–1102.
11.
GREENEG. L.LEPPLAN. C.DICKERSONW. A.1976. Velvetbean caterpillar: a rearing procedure and artificial medium. Journal of Economic Entomology69 (4): 487–488.
12.
HABIBM. E. M.PALEARIL. M.AMARALM. E. С.1983. Effect of three larval diets on the development of the armyworm, Spodoptera latifascia Walker, 1856 (Noctuidae, Lepidoptera). Revista Brasileira de Zoologia1 (3): 177–182.
13.
HADDADM. L.PARRAJ. R. P.MORAESR. C. B.1999. Métodos para estimar os limites térmico inferior e superior de desenvolvimento de insetos. FEALQ, Piracicaba. 29 p.
14.
HASSANS. A.1997. Seleção de espécies de Trichogramma para o uso em programas de controle biológico. p. 183–206. En: PARRAJ. R. P.ZUCCHIR. A. (eds.). Trichogramma e o controle biológico aplicado. FEALQ. Piracicaba. Brasil. 324 p.
15.
LEITER. C. D. C.LEALM. R. L. V.2007. O biocombustível no Brasil. Novos Estudos - CEBRAP (78): 15–21.
16.
MOLINAR. M. S.FRONZAV.PARRAJ. R. P.2005. Seleção de Trichogramma spp., para o controle de Ecdytolopha aurantiana, com base na biologia e exigências térmicas. Revista Brasileira de Entomologia49 (1): 152–158.
17.
OLIVEIRAJ. S.LEITEP. M.DE SOUZAL. B.MELLOV. M.SILVAE. C.RUBIMJ. C.MENEGHETTIS. M. P.SUAREZP. A. Z.2009. Characteristics and composition of Jatropha gossypiifolia and Jatropha curcas L. oils and application for biodiesel production. Biomass and Bioenergy 3PARRA, J. R. P. 1997. Técnicas de criação de Anagasta kuehniella, hospedeiro alternativo para produção de Trichogramma p. 121-150. En: PARRA, J. R. P.; ZUCCHI, R. A. (eds.). Trichogramma e o controle biológico aplicado. FEALQ. Piracicaba. Brasil. 324 p.
18.
PARRAJ. R. P.2009. A evolução das dietas artificiais e suas interações em ciência e tecnologia. p. 91–174. En: PANIZZIA. R.PARRAJ. R. P. (eds.). Bioecologia e nutrição de insetos: base para o manejo integrado de pragas. Embrapa Informação Tecnológica. Brasília, DF. Brasil. 1164 p.
19.
PARRAJ. R. P.ZUCCHIR. A.2004. Trichogramma in Brazil: feasibility of use after twenty years of research. Neotropical Entomology33 (3): 271–281.
20.
PASTRANAJ. A.2004. Los lepidópteros argentinos: sus plantas hospedadoras y otros sustratos alimenticios. Sociedad Entomológica Argentina, Buenos Aires. 334 p.
21.
POUSAG. P. A. G.SANTOSA. L. F.SUAREZP. A. Ζ.2007. History and policy of biodiesel in Brazil. Energy Policy35 (11): 5393–5398.
22.
PRATISSOLID.PARRAJ. R. P.2000. Desenvolvimento e exigências térmicas de Trichogramma pretiosum Riley, criados em duas traças do tomateiro. Pesquisa Agropecuária Brasileira35 (7): 1281–1288.
23.
PRATISSOLID.ZANUNCIOJ. C.VIANNAU. R.ANDRADEJ. S.PINONT. B. M.ANDRADEG. S.2005a. Thermal requirements of Trichogramma pretiosum and T. acacioi (Hym.: Trichogrammatidae), parasitoids of the avocado defoliator Nipteria panacea (Lep.: Geometridae), in eggs of two alternative hosts. Brazilian Archives of Biology and Technology48 (4): 523–529.
24.
PRATISSOLID.ZANUNCIOJ. C.VIANNAU. R.ANDRADEJ. S.ZANOTTIL. C. M.SILVAA. F. D.2005b. Biological characteristics of Trichogramma pretiosum and Trichogramma acacioi (Hym.: Trichogrammatidae), parasitoids of the avocado defoliator Nipteria panacea (Lep.: Geometridae), on eggs of Anagasta kuehniella (Lep.: Pyralidae). Brazilian Archives of Biology and Technology48 (1): 7–13.
25.
SAS INSTITUTE. 2002. SAS System - SAS/STAT. 9.1 ed. Cary, NC. SAS Institute.
26.
SHANKERC.DHYANIS. K.2006. Insect pests of Jatropha curcas L. and the potential for their management. Current Science91 (2): 162–163.
27.
SILVAINJ. F.LALANNE-CASSOUB.1997. Distinction entre Spodoptera latifascia (Walker) et Spodoptera Cosmioides (Walker), bona species [Lepidoptera, Noctuidae]. Revue Française d'Entomologie19 (3-4): 95–97.
28.
SPECHTA.SILVAE.LINKD.2004. Noctuídeos (Lepidoptera, Noctuidae) do Museu Entomológico Ceslau Biezanko, Departamento de Fitossanidade, Faculdade de Agronomia “Eliseu Maciel”, Universidade Federal de Pelotas, RS. Revista Brasileira de Agrociência10 (4): 389–409.
29.
STEINC. F.PARRAJ. R. P.1987. Aspectos bioecologicos de Trichogramma sp. em diferentes hospedeiros. Anais da Sociedade Entomológica do Brasil6 (1): 163–169.
30.
SUAREZP. A. Z.MENEGHETTIS. M. P.FERREIRAV. F.2006. O biodiesel e a política de C & T brasileira. Química Nova29 (6): 1157–1157.
31.
van LENTERENJ. C.2003. Commercial availability of biological control agents. p. 167–179 En: van LENTERENJ. C. (ed.) Quality control and production of biological control agents: theory and testing procedures. CABI Publishing. Wallingford. Reino Unido. 327 p.
32.
YUD. S. K.HAGLEYE. A. C.LAINGJ. E.1984. Biology of Trichogramma minutum Riley collected from Apples in Southern Ontario. Environmental Entomology13 (5): 1324–1329.
33.
ZUCCHIR. A.QUERINOR. B.MONTEIROR. C.2010. Diversity and hosts of Trichogramma in the new world, with emphasis in South America. p. 219–236. En: ConsoliF. L.ParraJ. R. P.ZucchiR. A. (Eds.). Egg parasitoids in agroecosystems with emphasis on Trichogramma. Progress in Biological Control. Volumen 9. Springer. Países Bajos. 482 р.