
Abstract
The responses of adult ichneumonids to colors were studied through comparing the effectiveness of standardized interception traps in seven different colors, in Liupanshan, Daxinggou and Xinbin (China). The captured ichneumonids belonged to 13 subfamilies: Anomaloninae, Banchinae, Campopleginae, Cryptinae, Ctenopelmatinae, Diplazontinae, Ichneumoninae, Mesochorinae, Ophioninae, Orthocentrinae, Phrudinae, Pimplinae, and Tryphoninae. Field trapping results indicated that yellow and green traps collected more ichneumonids than did the traps of other colors. Black was the least efficient color for collecting ichneumonids in Liupanshan and Xinbin. The most frequently collected specimens were from the subfamilies Ctenopelmatinae, Campopleginae and Cryptinae, while the least frequently collected were from Phrudinae, Ophioninae and Diplazontinae in Liupanshan and Xinbin. In Daxinggou, black and yellow traps collected more ichneumonids than did other color traps, with white traps collecting the least. Most captured ichneumonids at this site were from Campopleginae and Cryptinae, whereas only a small number of specimens were collected from Banchinae, Phrudinae, Ophioninae and Diplazontinae were collected.
Introduction
The vision of parasitoid wasps plays an important role in locating a host, mating, avoiding predators and searching for an oviposition site (Land 1997; Raine and Chittka 2007). Parasitoid wasps can find hosts by distinguishing the color or shape of hosts or host habitat (as well as by various olfactory signals) (Brown et al. 1998; Fischer et al. 2004; Segura et al. 2007). Itoplectis conquisitor (Say) (Hymenoptera: Ichneumonidae) can learn the characteristics of light reflected by artificial host microhabitats (Arthur 1966).
Insects have different types of photoreceptors that can detect different wavelengths of light, such as green, blue and UV light. Some hymenopterans not only can detect colors at 340 nm, 430 nm, and 535 nm, but also can detect those of long wavelengths up to 600 nm (Hardie 1986; Peitsch et al. 1992; Briscoe and Chittka 2001). The capture of insects in traps is influenced by the insect sensitivity to different colors among other variables. Based on these characteristics, entomologists use passive and active traps to collect insects. Malaise traps are widely used interception traps for catching flying insects, especially Diptera and Hymenoptera (Nieves-Aldrey and Rey del Castillo 1991; Steinbauer et al. 2000; Ganho and Marinoni 2003). These traps have also been extensively used for collecting Ichneumonidae (Townes 1962, 1972; Darling and Packer 1988; Anento and Selfa 1997; Skillen et al. 2000; Kumagai and Graf 2002; Sääksjärvi et al. 2004; Mazón and Bordera 2008).
Ichneumonidae is the largest family within Hymenoptera, which is probably the most diverse order in the world (Grissell 1999). Townes (1969) estimated that there could be about 60,000 ichneumonid species in the world whilst Gauld (1997) revised this figure upwards to potentially more than 100,000 species. They can be highly significant in suppressing populations of holometabolous pest insects and an important component in all terrestrial ecosystems. In order to better understand the biodiversity and population dynamics of Ichneumonidae in forest ecosystems, we carried out a comparative field trapping study using an improved interceptive trap on the effect of trap color on the captures of ichneumonids in Liupanshan, Ningxia Hui Autonomous Region, Daxinggou, Jilin Province and Xinbin, Liaoning Province, P. R. China.
Materials and Methods
Experimental sites
Field trapping experiments were conducted in the forests located at following three locations: Liupanshan, Xinbin and Daxinggou.
Liupanshan site is located at Liupanshan Mountains, in the border region of southwest Ningxia Hui Autonomous Region, east Gansu Province and west Shaanxi Province, which is the natural ecological shelter and water source areas of the western Loess Plateau, and plays an important role in eco-environmental conservation. The forest coverage rate reaches up to 72.8%, with a 5.8 °C annual mean temperature and 676 mm annual mean precipitation. The forest stand is composed of mixed deciduous angiosperms and evergreen conifers, mainly Pinus tabulaeformis Carr., Hippophae rhamnoides L., Elaeagnus angustifolia L. and Caragana korshinskii Kom.
Xinbin site is located at an extending branch of the Changbai Mountains, in the eastern part of Liaoning Province. Forest coverage is up to 64%, annual mean precipitation is 750-850 mm and annual mean temperature is about 6.1 °C. The forest stand is composed of deciduous angiosperms, mainly Quercus wutaishanica Blume and some shrubs.
Daxinggou site is a part of Laoyeling, Changbai Mountains, in the mid-western part of Wangqing County, Jilin Province. The forest coverage is up to 80%, annual mean precipitation is 550-600 mm and annual mean temperature is about 4.5 °C. The forest stand is composed of mixed deciduous angiosperms and evergreen conifers, mainly Larix gmelinii (Ruprecht) Kuzeneva, some Fagaceae and shrubs.
Traps
The interceptive traps used in current study were designed and constructed based on the Townes Malaise trap (Townes 1962) (Fig. 1). The trap like a typical ridge-pole house, with a vertex of triangular fork angled at 60 degrees. The traps were produced with seven colors of nylon netting: yellow, green, black, white, purple, red and blue.
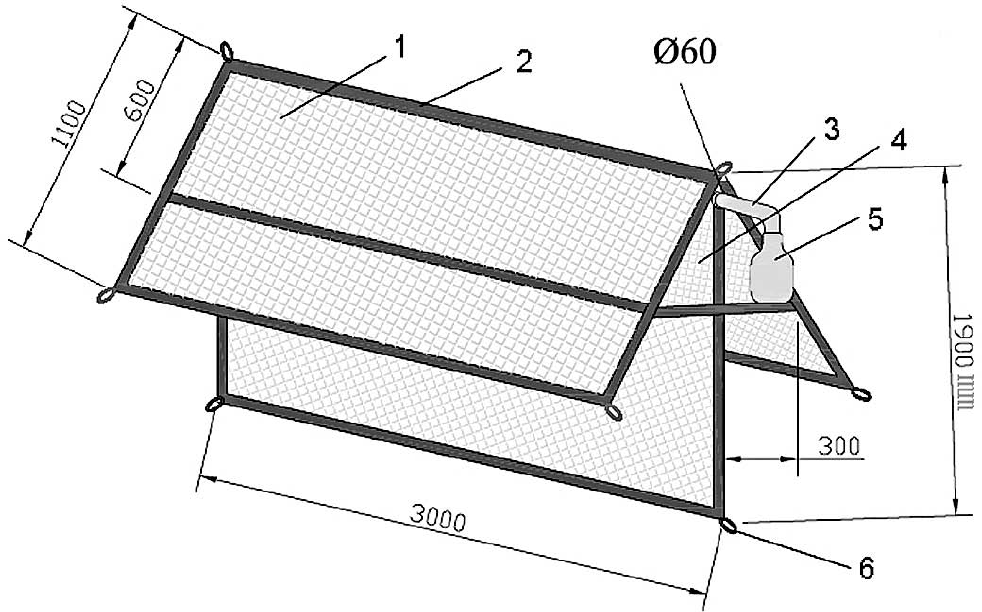
Interception trap. 1. Nylon net. 2. Frame. 3. PVC tubing 4. Triangular fork 5. Collecting bottle 6. Button loop.
The location of the trap is critical. It should be set up where insect flight tends to be concentrated by local circumstances of topography, vegetation, wind and water. Suitable places are across a small stream or gully, across a wide trail in the forest, at a low pass on a ridge or between two types of vegetation, etc. (Gressitt and Gressitt 1962).
Each trap was suspended between trees with the roof held open by ropes which run over a branch of a tree at an appropriate height and location, tied to the tree-trunk and the bottom secured with stakes. The lower edge was about 10-20 cm above ground level. The PVC tubing part was higher than other parts. Ichneumonids will run to the higher part and drop into the collecting bottle when they were intercepting by trap. The collecting bottle was filled with 70-80% ethanol.
Seven different color traps were set up randomly with 100 m apart between two neighboring traps in each experimental site. Specimens trapped were collected and ethanol in collection bottles was refreshed every week from June to October 2005 in Liupanshan and Daxinggou, from June to October 2009 in Xinbin. Identification and recording were carried out in the laboratory.
Statistical analysis
Trap catch data (numbers of ichneumonids per trap per week) were transformed by log (x + 1) and analyzed by one-way ANOVA, followed by the Ryan-Einot-Gabriel-Welsh (REGW) multiple Q test (SPSS 17.0 for windows) at α = 0.05.
Results
All specimens of ichneumonids were identified to subfamily level. Most trapped ichneumonids belong to following 13 subfamilies: Anomaloninae, Banchinae, Campopleginae, Cryptinae, Ctenopelmatinae, Diplazontinae, Ichneumoninae, Mesochorinae, Ophioninae, Orthocentrinae, Phrudinae, Pimplinae, Tryphoninae.
In Liupanshan site, green traps caught the highest numbers of ichneumonids, with an average of 168 per week (N = 17). Black traps were less successful with an average of just two ichneumonids per week (N = 17). There were no significant differences in ichneumonid catches between green and yellow traps (F = 43.97; df = 6, 112; P > 0.05), but both caught significantly more attractive than did the other color traps (P < 0.001). The blue traps caught only a few individuals but were still statistically higher than black traps (Fig. 2).
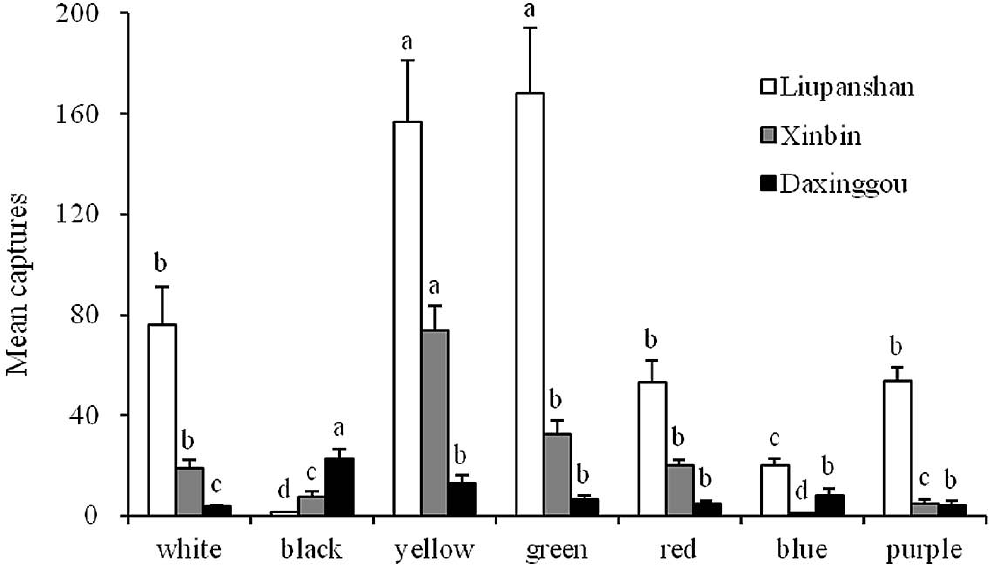
Mean numbers (+ SE) of ichneumonids captured per week in seven different color traps suspended in Liupanshan (N = 17) (June to October 2005), Xinbin (N = 17) (June to October 2009) and Daxinggou (N = 16) (June to October 2005), P. R. China. Bars marked with the same letters within the same site are not significantly different (ANOVA followed by REGWQ test: P > 0.05).
In Xinbin site, yellow traps caught the highest numbers of ichneumonids, with an average of 74 per week (N = 17), while blue traps were less successful with an average of just one ichneumonid per week (N = 17). There were significant differences in trap catches between yellow and other color traps (F = 49.56; df = 6, 112; P < 0.001). There were no significant differences in trap catches of ichneumonids between black and purple color traps (P > 0.05), but they both caught significantly more ichneumonids than did the blue color traps (P < 0.001) (Fig. 2).
In Daxinggou site, the overall trap catches were very low, the black traps being the best with an average of 23 ichneumonids per week (N = 16), and white traps being the less successful with an average of just four ichneumonids per week (N = 16). The black color traps caught significantly more ichneumonids than did other color traps (F = 5.23; df = 6, 105; P < 0.001), and there were no significant differences among other color traps except the white color traps (P > 0.05) (Fig. 2).
The number of ichneumonids at subfamily level captured in the three most attractive color traps (black, yellow and green) from each site were further analyzed (Figs. 3-5). There were significant differences in the trapping efficiency to different ichneumonid subfamilies among these three color traps.
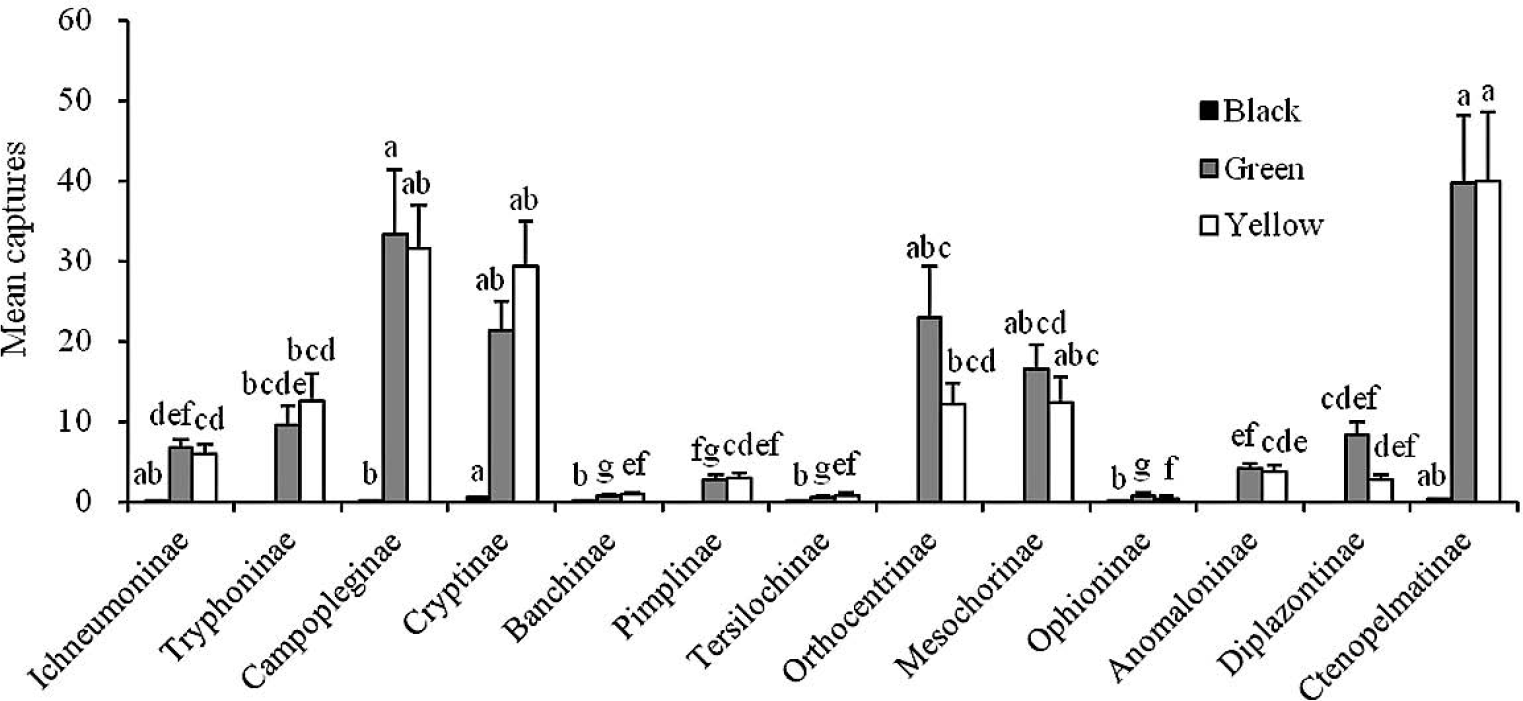
Mean numbers (+ SE; N = 17) of the major subfamilies of ichneumonids captured per week by black, green and yellow traps in Liupanshan. Bars marked with the same letters within each color are not significantly different (ANOVA followed by REGWQ test: P > 0.05).
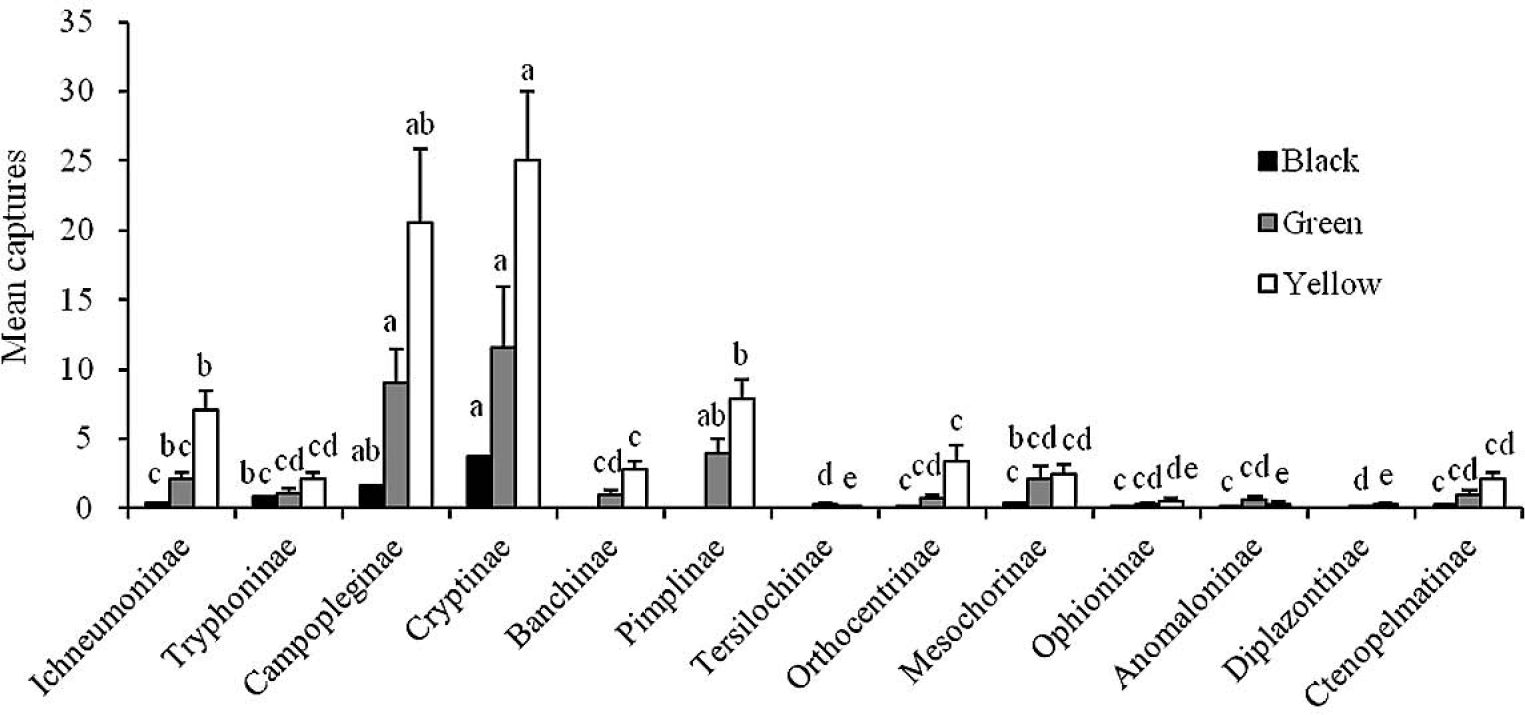
Mean numbers (+ SE; N = 16) of the major subfamilies of ichneumonids captured per week by black, green and yellow traps in Xinbin. Bars marked with the same letters within each color are not significantly different (ANOVA followed by REGWQ test: P > 0.05).
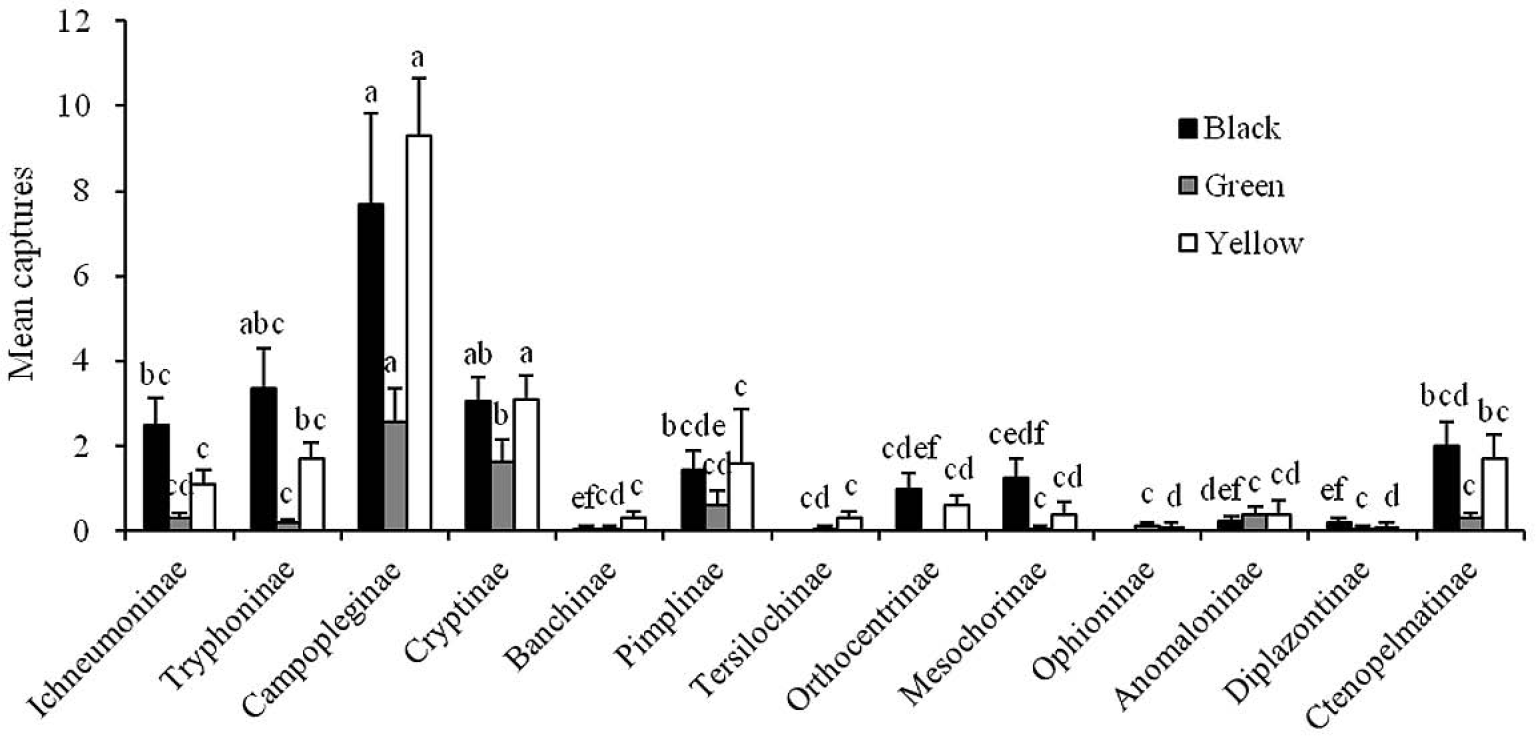
Mean numbers (+ SE; N = 16) of the major subfamilies of ichneumonids captured per week by black, green and yellow traps in Daxinggou. Bars marked with the same letters within each color are not significantly different (ANOVA followed by REGWQ test: P > 0.05).
In Liupanshan, Ctenopelmatinae, Campopleginae, Cryptinae, Orthocentrinae and Mesochorinae were the major subfamilies, there were no significant differences among these five subfamilies by green traps and there were no significant differences among Ctenopelmatinae, Campopleginae, Cryptinae and Mesochorinae by yellow traps (F = 22.76; df = 12, 208; P > 0.05) (Fig. 3). The numbers of ichnuemonids from Ctenopelmatinae, Campopleginae and Cryptinae seemed to be significantly higher than most of the minor subfamily group for both green and yellow traps (P < 0.001). There were only a few individuals of ichneumonids caught in the black traps at this site with Cryptinae, Ctenopelmatinae and Ichneumoninae being the key subfamilies. No specimens from the following subfamilies were collected in the black traps: Anomaloninae, Diplazontinae, Mesochorinae, Orthocentrinae, Pimplinae and Tryphoninae.
In Xinbin, Cryptinae and Campopleginae were two major subfamilies caught by yellow, green and black traps, and there were no significant differences between these subfamilies (F = 8.94; df = 12, 195; P > 0.05), but both caught significantly more richness than Ctenopelmatinae (P < 0.001) (Fig. 4). No specimens from following subfamilies: Diplazontinae, Tersilochinae, Pimplinae and Banchinae, were captured by black color traps, and only a few individuals of them were collected from the green and yellow traps.
Unlike the other two sites, black was the most attractive color to the ichneumonids at Daxinggou site. Cryptinae and Campopleginae were two major subfamilies captured by black and yellow color traps, followed by Tryphoninae, Ichneumoninae, Ctenopelmatinae and Pimplinae. There were no significant differences between Cryptinae and Campopleginae captured by black or yellow color traps (F =15.94; df = 12, 117; P > 0.05) (Fig. 5). No specimens from Ophioninae and Tersilochinae were captured by black traps. There were significant differences between Cryptinae and other subfamilies captured by green trap (P > 0.05), and only a few individuals of other subfamilies were captured by green traps. No specimens from Orthocentrinae were captured by green traps.
Discussion
Different insect families show preferences for different trap colors. Hoback et al. (1999) published a list of insect families with their preferences for different trap colors. Most insects that have been examined prefer yellow traps (Prokopy and Owens 1983). Farmland sawflies showed strong responses to colored traps, especially to yellow ones (Barker et al. 1997). Yellow is a strong innately preferred color by female Venturia canescens Gravenhorst, 1829 (Hymenoptera: Ichneumonidae) over orange and blue (Lucchetta et al. 2008).
In our study, the capture efficiency of color interception traps for ichneumonids were evaluated. Yellow and green proved to be much better than other colors in collecting ichneumonids and black was the less successful color in Liupanshan and Xinbin. However, there was a strong variation among sites: the least attractive color, black, in the above mentioned two sites, was the most attractive one at Daxinggou site. The real causes for such geographical (site) variations are unknown; however, differences in local microhabitats, forest stand structures and ichneumonid fauna might contribute to the disparity in color preference. Ctenopelmatinae, Campopleginae and Cryptinae were the most common subfamilies in Liupanshan. The latter two were also the most dominant subfamilies in Xinbin and Daxinggou. The diversity and relative abundance of the ichneumonids at subfamily level might be closely related to local habitat types. At Liupanshan site, the main vegetations are shrubs and pine plantations with coverage up to 95% of the ground area, suggesting a better environment and higher abundance of insects. Traps in Liupshan caught more ichneumonids in total than those in the other sites. In Xinbin and Daxinggou, the main vegetations are oak forests and coniferous forests respectively, with less insect biomass overall and fewer ichneumonids caught. Woodlands play an important role in the maintenance of biodiversity of parasitoids (Fraser et al. 2007). The response of ichneumonids to yellow, green and black color traps need further investigation.
Species belonging to the subfamilies Rhyssinae, Poemeniinae and Metopiinae are commonly found in the study areas but only a few specimens were captured by the interception traps. It might be due to the fact that these groups of ichneumonids are not strongly attracted to colors.
Footnotes
Acknowledgements
We are especially grateful to Dr. Gavin R. Broad (The Natural History Museum, London, UK) and Dr. Qing-He Zhang (Sterling International, Inc., USA) for their valuable comments and suggestions. We are thankful to two anonymous referees for their criticism and important suggestions. We also want to express our sincere gratitude to Xiao-Ren Xu, Feng-Cai He, Li-Xia Song and Shu-Feng Pan for their help in the course of field experiments. This project was funded by the National Natural Science Foundation of China (NSFC, No. 30872035; No. 31010103057).