
Abstract
Tutin inhibits the receptors of the neurotransmitter y-aminobutyric acid (GABA). In this work, we compared tutin with its derivatives, 2-(4-methoxybenzoyloxy)-tutin (T1), 2-(3,5-dinitrobenzoyloxy)-tutin (T2), 2-(6-chloronicotinoyloxy)-tutin (T3), and 2-(4-nitrobenzoyloxy)-tutin (T4), in terms of their effects on feeding and the feedback regulation mechanism of GABA metabolism. Spectrophotometry was used to determine the glutamic acid decarboxylase (GAD) and y-aminobutyric acid transaminase (GABA-T) activities of these compounds in 3rd instar larvae of Pseudelatia separata. Clear antifeedant activities were manifested by T1 and especially T3, whereas T4 stimulated feeding in P. separata. GAD and GABA-T activities in larvae treated with all five toxins differed significantly from the control and from one other (P < 0.05) in terms of treatment types and exposure times. The effects of the toxins on GAD and GABA-T were time-dependent for 48 h. GAD activities were inhibited by tutin, T2, T3, and T4, and were enhanced by T1. GABA-T activities were increased by all the toxins in varying degrees. Variation of GABA content in P. separata larvae resulted from the disturbance of GAD and GABA-T by tutin and its derivatives. Results suggest that T3, with its nicotinoyl group, is the most promising novel active ingredient for pest control.
Introduction
Tutin, a toxic compound of sesquiterpene extracted from Coriaria sinica Maxim (Coriariaceae: Coriaria), has neurotoxic, gastrotoxic, and antifeedant effects in many insects, and inhibits pathogenic microorganisms, such as Botryosphaeria berengeriana Notaris. f. sp. piricola (Nose, 1933) Koganezawa and Sakuma (1984), Alternaria macrospora Zimm., 1904, Phytophthora infestans (Mont.) de Bary (1845), and Pseudomonas solanacearum (Smith, 1896) (Zhang et al. 2006; Zhou et al. 2006). We therefore hypothesized that the use of tutin or its derivatives as a new pesticide warrants further study. We previously synthesized three derivatives (Li et al. 2007) that had higher antifeedant activities than tutin; however, these were still insufficient for adequate pest control. In the present study, we synthesized four new tutin derivatives based on our previous laboratory studies. Previous research has shown tutin to be an inhibitor of y-aminobutyric acid (GABA) receptors, which are nerve-muscle synapse inhibitory neurotransmitters in crustaceans and insects (Li et al. 2000a, 2000b, 2003).
When tutin acts on GABA receptors, the biosynthesis of GABA is affected (Nistri et al. 1997). The molecular structure of GABA is transformed in rats treated with tutin, leading to the accumulation of GABA because it cannot normally combine with its receptors. Thus, GABA biosynthesis and metabolism are affected (Schousboe et al. 1973; Erecinska et al. 1996; Sze 1997). These findings indicate that tutin can affect GABA biosynthesis in the physiological metabolism of organisms via feedback regulation. The research of Deewatthanawong et al. (2010) on GABA contents in CO 2 -treated tomatoes showed that GABA influences enzymatic activities during GABA biosynthesis.
Feedback regulation generally exists in the physiological metabolism of organisms. Shi et al. (2011) studied breast cancer cells and proved that the β2-adrenergic receptor and human epidermal growth factor receptor 2 comprise a positive feedback loop. Many enzymatic reactions in the process of cholesterol biosynthesis feature feedback regulation; an increase in cholesterol concentration causes a decrease in hydroxymethylglutaryl coenzyme A concentration, and vice versa (Sever et al. 2003).
Given the findings above, we hypothesize that the inhibition of GABA receptors by tutin will influence GABA contents and even GABA biosynthesis enzymes because of feedback regulation. This has not yet been confirmed in any previous study. GABA is involved in many metabolic activities, and its metabolic bypass is a branch of the citric acid cycle. Glutamic acid decarboxylase (GAD) and y-aminobutyric acid transaminase (GABA-T) are the GABA synthesizing and metabolizing enzymes, respectively (Sherif and Ahmed 1995). In the metabolic process, glutamate generates GABA by the catalysis of GAD, and GABA generates succinic semialdehyde by the catalysis of GABA-T (Zhu et al. 2007). Therefore, GABA content is influenced by GAD and GABAT activities. The object of this study was to assayed GAD and GABA-T activities in Pseudelatia separata (Walkers, 1865) (Lepidoptera: Noctuidae) to compare tutin with its derivatives in terms of their effects on GABA metabolism. P. separata is one of the most important crop pests in China and other countries in Asia. It can cause great threat on Agricultural production and food security. Cui et al. (2007) studied the toxicity of tutin with P. separata in the former researches too. We then used the results of these assays as a basis for selecting an optimal pesticide amongst tutin's derivatives.
Materials and Methods
Test insects
P. separata, were reared on fresh wheat leaves at the Pesticide Research Center, Northwest Agriculture and Forestry University, Yangling, Shaanxi, China. As normally done, two days after molting, third instar larvae with similar body sizes were selected for the experiments.
Toxins
Tutin was extracted with 95% alcohol by ultrasonic extraction method from dry seeds of Coriaria sinica Maxim in Integrated Pest Control Laboratory which locates on College of Forestry, Northwest A & F University. Then the extract was concentrated by rotary evaporation instrument and pure tutin was separated by chromatography (Cui 2007). The benzoyl derivatives of tutin 2-(4-methoxybenzoyloxy)-tutin (T1), 2-(3,5-dinitrobenzoyloxy)-tutin (T2), 2-(6-chloronicotinoyloxy)-tutin (T3), and 2-(4-nitrobenzoyloxy)-tutin (T4) were respectly synthesized by change 2-hydroxy group of tutin with 4 - methoxy benzoyl chloride, 3,5-dinitrobenzoyl chloride, 6 - chlorine chloride and 4-nitrobenzoyl chloride (Fig. 1).
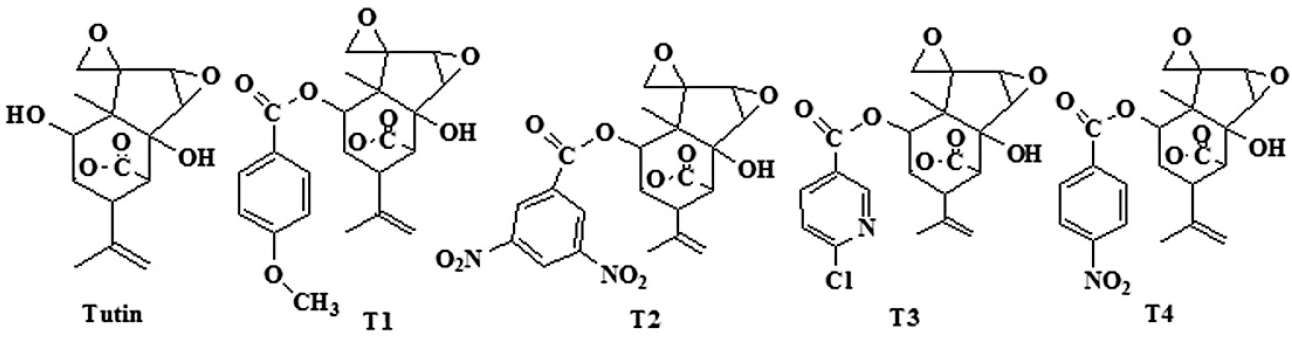
Structures of tutin and its derivatives: 2-(4-methoxy-benzoyloxy)-tutin (T1), 2-(3,5-dinitrobenzoyloxy)-tutin (T2), 2-(6-chloronicotinoyloxy)-tutin (T3), and 2-(4-nitrobenzoyloxy)-tutin (T4).
Chemicals
Analytically pure acetone, β-mercaptoethanol, phosphate buffer (NaH 2 PO₄·2H 2 O-Na 2 HPO₄·12H 2 O), borate buffer (H₃BO₃-NaOH), phenol, and sodium hypochlorite solution were used. GABA, L-glutamic acid, 5-pyridoxal phosphate, NAD, and α-ketoglutarate were purchased from WOLSEN.
Determination of antifeedant activities
For the toxin sample solutions, tutin and its derivatives were dissolved in acetone to a final concentration of 2.0 mg/mL following Guo et al. (2009). Acetone was used as the control.
The nonrestrictive leaf-disc method was used to determine the antifeedant activities of the compounds (Mu 1997). Fresh young wheat leaves were cut into round leaf-discs 10 mm in diameter, and each leaf-disc was evenly coated with 5 µL of the sample solution on one side. Once the acetone had evaporated, the infected leaf-discs were ready for testing. A piece of qualitative filter paper with diameter 9 cm was then laid at the bottom of 9 cm-diameter Petri dishes and tap water was sprayed onto it to maintain the relative humidity at 60-90%. We used a total of 30 Petri dishes and 180 test larvae. Every five Petri dishes were grouped as one, and each dish placed six 3rd instar larvae starved for 12 h and thirty infected leaf-discs were placed.
Each of the groups of five dishes in a group was separately treated with the infected leaf-discs of control, tutin, T1, T2, T3, or T4 for 4, 8, 12, 24, and 48 h. The eaten areas of the leaves were measured with quadrille paper (Feng and Shi 2006) after 4, 8, 12, 24, and 48h of incubation at 18-22°C and light-dark cycling with L:D = 12:12h. Each experiment was repeated three times. Water and the corresponding infected leaf-discs were replenished and recorded in the process. Antifeedant activity was measured by calculating the antifeedant rate: Antifeedant rate = [(eaten area of control – eaten area of toxin) / eaten area of control] x 100%.
Preparation of crude enzymes
Every group of six test larvae was weighed after 4, 8, 12, 24, and 48 h of treatment. The larvae, approximately 20mg quartz sand, and phosphate buffer (1 mL, pH 7.0, 0.04 M) were then homogenized in an ice bath using a pestle. The homogenate was transferred to a centrifuge tube (2.0 mL), along with the washing liquor from and phosphate buffer to a volume of 2.0 mL. The tube was centrifuged at 10000 rpm for 15 min at 4°C (Eppendorf centrifuge, 5415R, Germany) (Jayakumar et al. 1999). The supernatant was then recovered as crude enzyme and stored at 4°C until use.
Determination of GAD activity
Determination of GAD activity was based on the principles of the reaction of Berthelot (Xu et al. 2004; Tao et al. 2006). The crude enzyme (V₀= 0.2 mL) and solution I composed by 0.2 mL, pH 7.0 phosphate buffer containing 10 mM L-glutamic acid and 0.1 mM 5-pyridoxal phosphate were placed into a test tube (10 mL, polypropylene) and incubated at 30°C for 30 min. The incubated mixture was placed in an ice bath, after which borate buffer (0.4 mL, 0.2 M) was added to terminate the reaction. Phenol (2 mL, 6%) and sodium hypochlorite solution (0.4 mL, containing 10% active chlorine) were also added to finish the color reaction. The mixture was mixed thoroughly by hand, placed in a hot water bath (100°C) for 10 min, and then rapidly cooled in an ice bath for 20 min. The absorbance of the solution at 630 nm was then measured and recorded using a 752N type UV-VIS spectrophotometer (Shanghai Precision and Scientific Instrument CO., LTD). GAD measurements were performed in triplicates for each crude enzyme extract. GAD activity was expressed as absorbance per unit insect weight, as indicated in the formula: GAD activity (U/g) = A₆₃₀/(FW × V₀/V), where FW = larvae weight (g), V₀ = the determining volume of crude enzyme (mL), and V = total volume of crude enzyme (mL). The GAD relative activity, which shows the relative effect between the toxins and the control, was calculated using the formula: GAD relative activity = [(average GAD activity of toxin - average GAD activity of control) / average GAD activity of control] × 100% (Xu et al. 2004; Tao et al. 2006).
Determination of GABA-T activity
We followed the procedure outlined by Niladri et al. to determine GABA-T activity (Basu et al. 2010). Crude enzyme (V₀ = 0.3 mL) was added to phosphate buffer (3.5 mL, 100 mM, pH 8.75, containing 1mM NAD and 3.5 × 10⁻β mM β-mercaptoethanol). The mixture was shaken and solution II (0.2 mL, containing 4.8 mM α-ketoglutarate and 18mM GABA, pH 7.0) was added. The mixture was incubated at 30°C for 30 min, after which its absorbance at 340 nm was measured and recorded using a 752N type UV-VIS spectrophotometer (de Graaf et al. 2006; Suzuki et al. 2009; Li et al. 2007). GABA-T measurements were performed in triplicates for each crude enzyme extract. GABA-T activity was presented as absorbance per unit insect weight, as indicated in the formula: GABA-T activity (U/g) = A₃₄₀/(FW × V₀/V). GABA-T relative activity was calculated in the same manner as the GAD relative activity.
Data analysis
Analyses of variance were used to assess the effects of toxins and time (independent variables) on GABA shunt enzymes (dependent variables), and Duncan's multiple range tests (SSR method) were used to determine pair-wise differences. The critical level of significance for all statistical analyses was set at α = 0.05 using SPSS (Statistical Product and Service Solutions) version 13.0. Graphs were drawn using Originlab version 8.0 software.
Results
Antifeedant activity
All the antifeedant activities of P. separata treated with actone and toxins were similar in the 48 h. The antifeedant activities of P. separata treated with T1, T2, and T3 were higher than that treated with tutin, whereas T4 had the lowest antifeedant activity and even stimulated the appetite of P. separata. The highest antifeedant rate was observed after 8 h in T3 (88%), while the second highest rate was observed in T1 (Fig. 2).
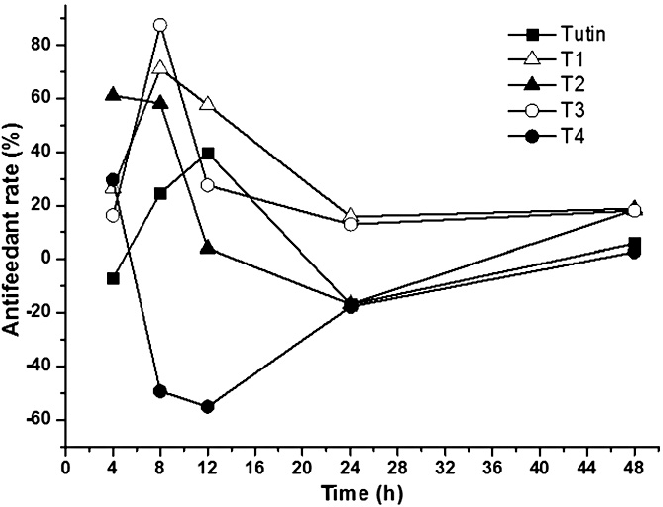
Antifeedant rates in Pseudelatia separata larvae after exposure to tutin and its derivatives (T1, T2, T3, and T4). Negative values represent increasing food intake and positive values represent decreasing food intake.
GAD activity
Except for T1, all toxins inhibited GAD activities, with T3 and T4 inhibiting to a greater degree (Fig. 3). Inhibition of GAD activity was higher in treatments with the control, tutin, and T1 than in those with T2, T3, and T4 (Fig. 4). GAD activities for all six groups (the control and the five toxins) showed a decreasing trend with time.
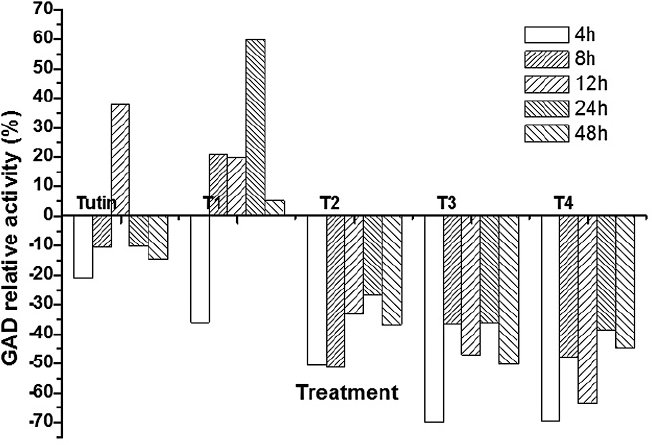
Relative activities of GAD in P. separata larvae after exposure to tutin and its derivatives (T1, T2, T3, and T4). Negative values represent inhibition of enzyme activity and positive values represent promotion of enzyme activity.
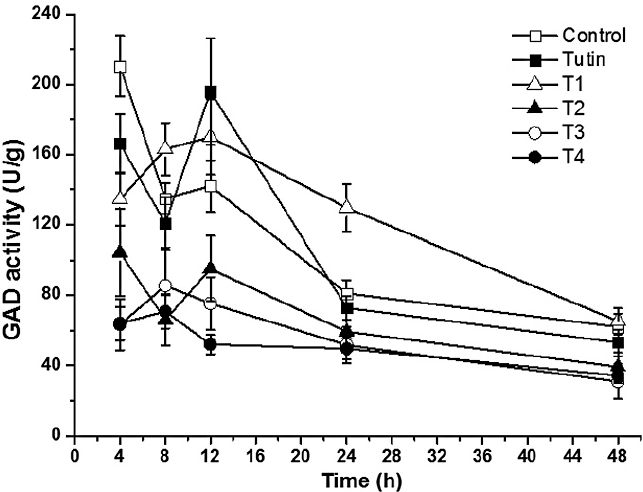
Dynamics of GAD activities with different treatments.
GAD activities in P. separata treated with toxins were significantly different from the control (Fig. 5). Using Duncan's multiple range tests, we found that insects treated with tutin, T2, and T3 showed significant differences in inhibition of GAD activity after different lengths of exposure time. Generally, changes in GAD activities were similar between the 4 and 8 h treatments, and both were significantly different from the 12, 24, and 48h treatments. The best inhibited effect was observed at 48h, and the lowest GAD activity was in the sample treated with T3.
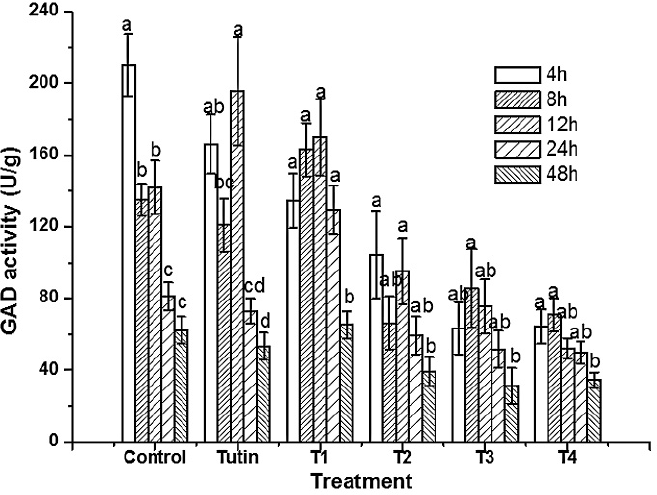
Differences in GAD activities amongst different treatment types and exposure times. The small letters represented the differences between groups.
GABA-T activity
T1 treatment completely promoted GABA-T activities, whereas T2, T3, and T4 yielded fluctuating effects on GABA-T activities within 48 h of exposure (Fig. 6). Before 12 h, tutin promoted GABA-T activities, but inhibited them after 24 h. Except for T1, all the other derivatives disturbed GABA-T activities in test larvae, particularly the T3 treatment. GABA-T activities decreased with increasing exposure time (Fig. 7). At 4 h, although all toxins showed fluctuating effects on GABA-T activity, they all promoted it. Significant differences in GABA-T activities were observed amongst the five toxins and the control and amongst different assessment times (Fig. 8).
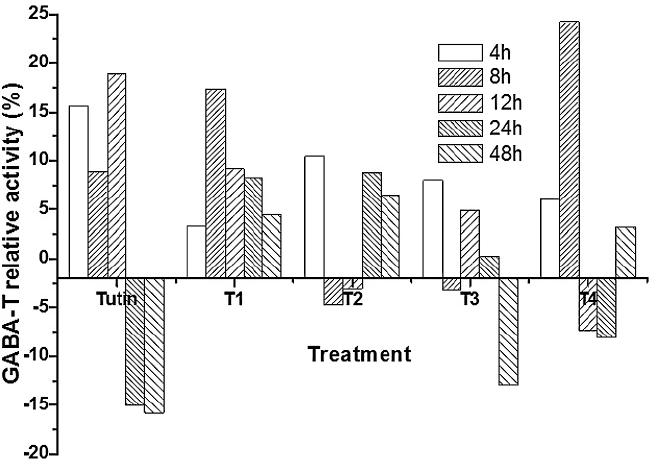
Relative activities of GABA-T in P. separata larvae after exposure to tutin and its derivatives (T1, T2, T3, and T4).
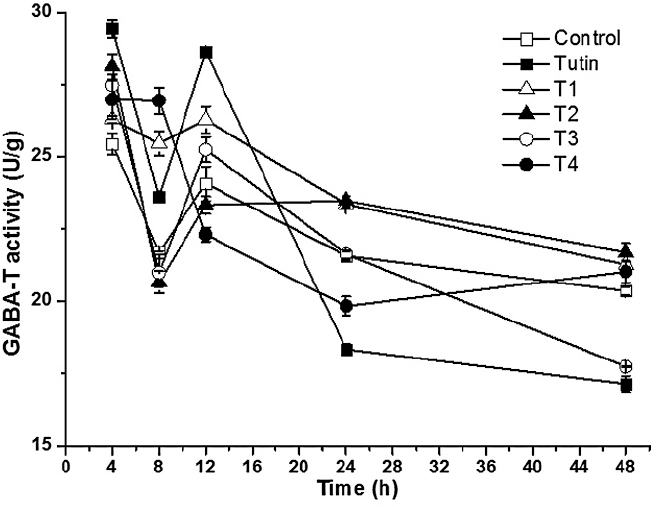
Dynamics of GABA-T activities with different treatments.
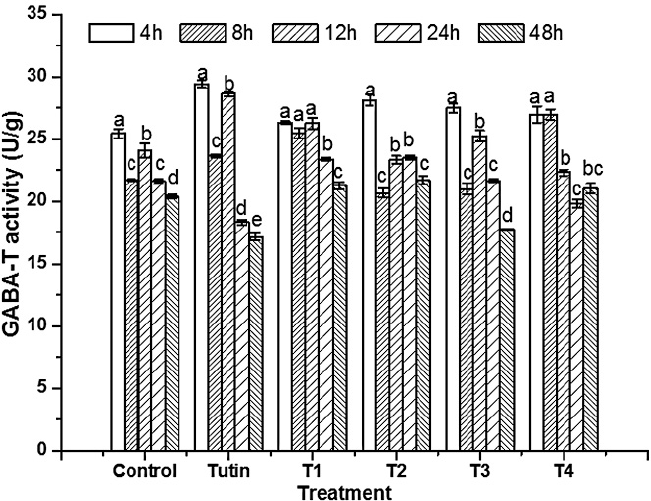
Differences in GABA-T activities amongst different treatment types and exposure times. The small letters represented the differences between groups.
Discussion
Li et al. (2007) found that tutin had antifeedant effects on P. separata, and 2-iso-butenoyltutin, one of the derivatives synthesized from tutin, significantly increased antifeedant activity compared with tutin. In the present study, the 2-hydroxy group of tutin was replaced into four other groups, resulting in four novel derivatives (T1 to T4). We fed 3rd instar larvae of P. separata with wheat leaves inoculated with tutin and these new derivatives, and found that the antifeedant rate of T3 was higher than that of 2-iso-butenoyltutin, making T3 a promising substance for pest control.
We further studied the relationship between antifeedant activity and GABA shunt enzymes. The content of the neurotransmitter GABA in the central nervous system of the mammalian brain is related to the available amounts of GAD, GABA-T, and other enzymes (Sherif and Ahmed 1995). The inhibition of GAD and GABA-T activities showed that GABA synthesis and metabolism were blocked, and vice versa. The results of the present study show that tutin and its benzoyl derivatives have significant effects on both GAD and GABA-T activities in the test larvae. The tutin derivatives, T2, T3, and T4, stably inhibited GAD activities within 48 h of exposure, whereas T1 promoted them. T3 had the strongest inhibitory effect on GAD activity. Tutin and its derivatives also showed fluctuating effects on GABA-T activities. T2, T3, and T4 disturbed GABA-T activities in test larvae, particularly T3. This led to a variation in GABA content and a disturbance in the metabolic rate, resulting in disorders in nerve conduction and physiological metabolic difficulties in the test larvae. The results also further explain the study that Glu and GABA contents were influenced by tutin in test insects (Li et al. 2000a). Tutin and its derivatives acted on GABA receptors (Fuentealba et al. 2007), however, according to the feedback regulation mechanism, GAD and GABA activities were influenced, which subsequently affects GABA biosynthesis and metabolism (de Graaf et al. 2006).
We found that GABA has a feedback effect on GAD and GABA-T, and that T3 [2-(6-chloronicotinoyloxy)-tutin], which is derived from tutin, is the most promising novel active ingredient for pest control. T1 had the lowest inhibitory effect on GAD activities, and was significantly different from the three other derivatives. T3 showed the highest antifeedant activity and significant effects on GAD and GABA-T activities. The structures of the derivatives may provide reasons for these differences observed. There exists a methoxy group in T1, a nitro group in T2, a nicotinoyl group in T3, and a double-nitro group in T4. The nicotinoyl group, which results from modification of the 2-hydroxy group of tutin into a benzoyl group, appears to affect enzyme activities more than either the nitro or methoxy groups. From Duncan's multiple range tests, no significant differences between the effects of T2 and T4 were observed on GAD and GABA-T activities. Thus, substituting the 2-hydroxy group of tutin for benzoyl derivatives with one or two nitro groups exerts similar effects on enzymes activities. Such as imidacloprid, thiacloprid, acetamiprid, and nitenpyram, contain nicotinoyl groups that cause contact toxicity, stomach toxicity, absorption toxicity, and so on. Linking a nicotinoyl group to the 2-hydroxy group of tutin not only enhanced its antifeedant activity in test larvae, but also promoted the advantages of nicotinoyl insecticides.
Footnotes
Acknowledgements
This work was supported by the National Natural Science Foundation of China (No. 30872032).