Alnus acuminata is a native species used in reforestation and industrial forestry programs in Colombia. It is currently attacked by an insect recently described as Corthylus zulmae, which is inhibiting the use of this plant. Establishing management strategies for this pest is required, which is why it is necessary to understand basic aspects of its biology. In this study we evaluated the population dynamics of the pest over one year through destructive sampling of affected trees. The adult flight period was followed using traps with attractants; observations on the life cycle were made by following the insect biological stages collected in the field; and substrates for rearing the insect were evaluated in the laboratory. The results showed that this insect reproduces in decaying trees or trunks in the soil with low humidity content. This insect showed greater flight activity when temperature was higher, and precipitation and relative humidity were lower. A description of the life stages was made and we estimated that a complete cycle might last more than 48 days and, that eggs could require around 28 days in order to develop into an adult. The artificial diets reported for rearing other coleopterans in the laboratory were not efficient for establishing a confined colony of C. zulmae.
Alnus acuminata H.B.K. subsp. acuminata (Betulaceae), llamada comúnmente Aliso, es un árbol nativo de los Andes que crece entre los 1700 y 3300 msnm. En Colombia se encuentra naturalmente en las zonas de vida clasificadas de acuerdo con Holdridge (1987) como Bosque húmedo montano bajo (bh-MB), Bosque muy húmedo montano bajo (bmh - MB) y Bosque pluvial o Páramo pluvial montano (bp-M) en los departamentos de Caldas, Quindío, Risaralda, Huila, Boyacá, Nariño y Santander. Esta especie se adapta a temperaturas entre 8,5 y 18°C y precipitaciones anuales de 1.000 a 2.000 mm (Del valle y González 1988). Se destaca su uso en la protección y recuperación de cuencas hidrográficas, juega un papel importante en el control de la erosión y es utilizada con buenos resultados en sistemas agroforestales y silvopastoriles gracias a su asociación con microorganismos fijadores de nitrógeno (Briceño 2002). La madera de A. acuminata se usa en construcciones livianas, formaletas para concreto, en la industria de tableros de viruta y aglomerados, en la fabricación de lápices, fósforos, toneles, alacenas, puertas, ventanas, postes, entre otros.
En el año 2003 las plantaciones de A. acuminata localizadas en la cuenca del río Blanco, en el municipio de Manizales, Caldas, se vieron afectadas por un insecto barrenador identificado como Corthylus zulmae Wood, 2007 (Coleoptera: Curculionidae: Scolytinae) (Wood 2007). En los árboles afectados por este insecto se observó acumulación de aserrín blanquecino en la parte inferior de los orificios de entrada, produciendo pérdida de calidad de la madera; en forma general en las plantaciones se presentó marchitamiento de árboles, seguida de defoliación total, y por último, alta formación de rebrotes basales e intermedios; asociados a estos daños se encontraron hongos fitopatógenos como Fusarium solani (Link ex Gray), Fusarium sp. y Ceratocystis sp. y una levadura conocida como Pichia sp. (Gil et al. 2004; Ospina et al. 2005). Los únicos reportes que se tenían hasta entonces de este género en el país, corresponden al año 1989 cuando se observaron algunos individuos atacando varias especies de fabáceas como Inga spectabilis Vahl, Inga edulis Mart e Inga sp., sin embargo, su efecto no revistió mayor importancia (Posada 1989).
La larva de C. zulmae es vermiforme, de color blanco, ápoda y en forma de C, con mandíbulas esclerotizadas. La pupa es blanca cremosa y típicamente exarata. El adulto mide aproximadamente 3,0 mm de largo y 1,16 mm de ancho. El cuerpo es robusto, la cabeza y el tórax de color marrón a negro y en la parte anterior de los élitros presenta una mancha de color marrón - amarillo, el resto es de color marrón - negro. En las áreas superior y lateral del pronoto presenta líneas de espinas. La especie presenta dimorfismo sexual; las hembras tienen la frente cóncava con setas cortas y abundantes, la clava antenal es larga, con el margen anterior ornamentado por un grupo de setas largas. El macho presenta la frente convexa sin setas, la clava antenal no presenta el grupo de setas descrito para la hembra (Gil et al. 2004; Wood 2007).
Los coleópteros de las subfamilias Scolytinae y Platypodinae (Curculionidae) se encuentran entre el grupo de insectos de mayor importancia en plantaciones forestales. Son particularmente importantes debido a algunas características como el uso de feromonas de agregación, que genera la colonización en masa de los hospedantes y la interacción con hongos asociados que causan la muerte de los árboles. Debido a esta interacción son llamados comúnmente "coleópteros ambrosiales" (Furniss et al. 1987; Cibrian et al. 1995; Harrington 2005). Estos coleópteros desempeñan cuidado parental con la progenie (Normark et al. 1999) y construyen galerías para cultivar los hongos simbiontes, los cuales son utilizados como fuente de alimento durante todo su ciclo vital (Kuhnholz et al. 2001). Los huevos son colocados en el jardín del hongo que es cultivado en las galerías cavadas en el interior del xilema (Farrell et al. 2001).
Los adultos del género Corthylus perforan el fuste y las ramas de los árboles vivos débiles o derribados. En algunas especies, el ataque es iniciado por la hembra; en otras el macho realiza la entrada de la galería principal y parte inicial de una o dos galerías transversales de oviposición. La hembra copula con el macho, entra a la galería principal y continúa con la excavación de las galerías de oviposición en cada una de las cuales deposita un huevo; existen especies en las cuales las hembras depositan sus huevos libremente en las galerías y los estados jóvenes y adultos viven juntos en las mismas cámaras (Steinhaus 1947; Comstock 1962; Cibrian et al.1995). Al inicio de la colonización, los adultos, principalmente las hembras, transportan el hongo en la superficie del cuerpo o en estructuras especializadas denominadas "micangias" o micetangias definidas como una invaginación del integumento con glándulas o células secretoras que protegen las esporas y el micelio del hongo de la desecación y poseen acción contra microorganismos. Estas micangias se encuentran localizadas en ciertas partes del cuerpo como las mandíbulas, élitros y tórax y los hongos son transmitidos de la madre a la progenie (Beaver 1989; Tanada y Kaya 1993; Fraedrich et al. 2008).
Los insectos permanecen gran parte de su ciclo de vida dentro de las galerías hechas en las plantas hospedantes, en donde las condiciones del microclima son muy constantes. Las condiciones externas de temperatura, humedad relativa, precipitación y radiación solar, desempeñan un papel importante en la distribución de estos insectos (Wood et al. 1992).
La capacidad de vuelo de los organismos es afectada por factores primarios en la atracción que ejercen las oleoresinas, terpenos, alcoholes y otras sustancias emitidas por el tejido de los árboles recién cortados o muertos. Cuando uno de estos insectos encuentra un medio apropiado para alimentarse y reproducirse, empieza a barrenar para iniciar la liberación de compuestos volátiles producto de la fermentación de la madera, así como feromonas en sus excrementos, que atraen a ambos sexos de su propia especie (Wood 1982). En Canadá, el periodo de vuelo para C. punctatissimus (Zimmermann, 1868) se presenta principalmente entre junio y agosto (Finnegan 1967).
Las diferentes especies de Corthylus, presentan dos o más generaciones al año, dependiendo de las condiciones de altitud y latitud donde se encuentren (Cibrian et al. 1995). En el Sur de Indiana en los Estados Unidos, C. columbianus Hopkins, 1895 presenta dos generaciones por año; los huevos duran seis días aproximadamente, las larvas de 12 a 15 días, prepupa de dos a tres días, pupa de ocho a nueve días y los adultos de 34 a 40 días (Kabir y Giese 1966), mientras que en Ontario, Canadá, Corthylus puntactissimus presenta solo una generación por año (Finnegan 1967).
En América se conocen cerca de 90 especies del género Corthylus distribuidas desde Canadá hasta Argentina, 56 de éstas se encuentran en Norte y Centro América (Wood 1982). En Colombia se han reportado C. zulmae y Corthylus andinus Wood, 2007 en los departamentos de Caldas y Huila (Gil et al. 2004; Wood 2007), sin embargo, aún se hacen inventarios en otras partes del mundo.
La incidencia causada por C. zulmae en las plantaciones de A. acuminata en el departamento de Caldas ha desestimulado a los reforestadores y ha generado la necesidad de buscar alternativas de manejo para esta plaga. Esto hace importante conocer la etología, el ciclo de vida y las condiciones que favorecen la reproducción y dispersión de este insecto. Dado que en Colombia no se ha desarrollado estudios detallados sobre esta especie, el objetivo de esta investigación fue contribuir al entendimiento de la biología de C. zulmae; para lo cual se estudió la dinámica poblacional del insecto en el campo, se realizó una aproximación a su ciclo de vida y se evaluaron sustratos para su cría en condiciones de laboratorio.
Materiales y Métodos
Localización. Este estudio se realizó entre abril de 2008 y diciembre de 2009 en una plantación de Aliso en la finca La Fe (5°00,4'69″N 75°23.4′28″W), vereda Gallinazo del municipio de Villamaría en el departamento de Caldas, Colombia, perteneciente a la empresa Aguas de Manizales a una altitud de 2800 msnm, en un lote de aproximadamente 1,2 hectáreas, con una densidad de población de 715 árboles/ha. Los estudios de laboratorio se realizaron en el cuarto de cría de insectos del Centro Nacional de Investigaciones de Café, Cenicafé, con condiciones controladas de temperatura de 17,7°C y humedad relativa del 70%, similares a las de campo.
Dinámica poblacional de C. zulmae. Para determinar el número de árboles afectados en el lote experimental, se realizó un censo trimestral durante 12 meses, a partir del cual se obtuvo el porcentaje de infestación ocasionado por C. zulmae. Posteriormente, durante ese mismo tiempo y con una periodicidad mensual, se realizaron disecciones de árboles afectados. Para esto se cortaron los árboles, se tomaron las partes afectadas del tallo, se transportaron al laboratorio de cría, se fraccionaron longitudinalmente utilizando una sierra y se extrajeron los estados biológicos del insecto. Se estimó en cada tiempo de muestreo, el promedio de perforaciones por árbol y el rango de altura a las cuales se encontraron; la proporción de árboles y de perforaciones con presencia de huevos, larvas, pupas y adultos de C. zulmae y la relación de sexos en la población adulta. Se realizó prueba de comparación de Duncan, de los promedios mensuales para identificar los meses con mayor promedio en las variables descritas. Adicional a esto, se realizaron muestreos destructivos de troncos de Aliso que se hallaron en el suelo con numerosas perforaciones durante los meses de julio, agosto, septiembre y octubre de 2009 con el fin de registrar el número de estados presentes y la proporción de troncos que contenían individuos.
Se registró el vuelo de los adultos de C. zulmae a través del tiempo. Para ello, se instalaron 30 trampas de captura (BROCATRAP - www.biogardenlab.com) con una mezcla de metanol: etanol en una proporción de 3 a 1, distanciadas aproximadamente 20 metros entre sí. Se contabilizó el número de adultos capturados por trampa cada quince días durante un año; se estimó el promedio de adultos capturados por trampa, en cada tiempo de muestreo y se construyó la variable acumulado de adultos a través del tiempo, y con ella se estableció la relación lineal con el número de estados, con el fin de identificar si el vuelo estuvo asociado con mayor ataque de la plaga. Durante el tiempo de evaluación se llevaron los registros de clima, como temperatura, humedad relativa y precipitación, para complementar el análisis de la información.
Estimación del tiempo del ciclo de vida de C. zulmae. Con el fin de estimar preliminarmente la duración de los estados de vida del insecto. Se recolectaron muestras de individuos que se encontraban en el interior de las galerías en alisos afectados en campo y se llevaron al laboratorio de cría en Cenicafé. Se individualizaron secciones de troncos conservando los estados del insecto sobre las galerías de oviposición, en recipientes plásticos con algodón húmedo. Se realizó una descripción de cada estado y se hizo un seguimiento diario hasta el momento en que el estado observado murió.
Evaluación de un sustrato para la cría de C. zulmae. Se evaluaron cuatro sustratos de cría para mantener a C. zulmae y determinar su biología en condiciones de laboratorio. Se definió como unidad experimental un recipiente que contenía una pareja de insectos. Por cada sustrato, se tuvieron 60 unidades experimentales bajo un diseño completamente aleatorio, en el ambiente del laboratorio de entomología de Cenicafé. los sustratos fueron los siguientes:
Dieta artificial (Cenibroca modificada). Esta dieta es una modificación de la usada para la cría de la broca del café (Hypothenemus hampei Ferrari, 1867) en la cual se reemplazó el café por aserrín de Aliso en la misma proporción. Para la elaboración de la dieta se siguió el protocolo establecido por Portilla y Streett (2006) y se depositó una cantidad aproximada de 5 ml en viales estériles.
Troncos de Aliso con agar y aserrín (Kabir y Giese 1966 modificado). Se depositaron 50 cc de una mezcla de agar al 1% y aserrín de aliso, en frascos de vidrio de 200 cc у antes de la solidificación, se introdujo una sección cuadrada de tronco de Aliso (5 cm de ancho por 5 cm de largo y 4 cm de profundidad), conservando el lado de la corteza descubierto de la mezcla. Tanto el aserrín como los troncos fueron esterilizados previamente a 120°C de temperatura y a 15 libras de presión durante 25 minutos.
Troncos de Aliso. Se tomaron secciones de tronco de Aliso (15 cm de longitud por 10 cm de ancho y 6 cm de profundidad) esterilizados previamente y se depositaron en recipientes plásticos sobre una capa de algodón humedecido. El recipiente se cubrió con una tapa que permitía la circulación del aire para evitar proliferación de microorganismos.
Ramas secundarias de Aliso. Se introdujeron ramas de Aliso (10 cm de longitud y diámetro aproximado de 3 cm) esterilizadas previamente, en un recipiente plástico sobre una capa de algodón humedecido. El recipiente se cubrió con una tapa que permitía la circulación del aire para evitar la proliferación de microorganismos.
Una vez establecida cada unidad experimental para cada sustrato, se realizaron las infestaciones de los tratamientos tomando una pareja de C. zulmae recolectados en campo, se esterilizaron en hipoclorito de sodio al 3%, se lavaron con agua destilada y se introdujeron en cada uno de los recipientes conteniendo el sustrato. Cada 20 días durante dos meses, se tomaron aleatoriamente, 20 unidades experimentales de cada tratamiento, y mediante muestreo destructivo, se registró el número de huevos, larvas y pupas, mortalidad de cada estado y relación de sexos en individuos adultos. Con esta información se realizaron comparaciones estadísticas de las poblaciones del insecto en cada sustrato.
Resultados y Discusión
Dinámica poblacional de C. zulmae en campo. El porcentaje de infestación inicial en el lote fue del 13%; el ataque aumentó rápidamente en la plantación durante el tiempo evaluado, duplicándose el número de árboles infestados en tan solo tres meses, tiempo después del cual aumentó hasta niveles del 38% (Fig. 1). Los árboles presentaron síntomas de ataque reciente como exudado y acumulación de aserrín en la parte inferior de los orificios de entrada del insecto (Figs. 2A y 2B), y síntomas de ataques viejos donde se observaron orificios de penetración con alrededores de coloración oscura y seca (Fig. 2C). La mayoría de los árboles afectados no presentaron marchitamiento ni defoliación total, los cuales son síntomas reportados como característicos del daño causado por C. zulmae (Gil et al. 2004; Ospina et al. 2005). Se encontraron árboles afectados en toda la plantación, sin embargo, la tendencia al mayor ataque del insecto se observó en árboles ubicados en los bordes del lote que limitaban con áreas de bosque natural y una menor población de árboles atacados en áreas cercanas a claros y bordes de camino (Fig. 1).
Mapa del lote experimental en la finca La Fe, municipio de Villamaría, Caldas donde se muestran los árboles sanos , y afectados durante el primero (13%), segundo (29%), tercero (33%), cuarto (36,5%) y quinto censo (38,6%) y la ubicación de las trampas de alcohol . El porcentaje acumulado de infestación se indica entre paréntesis.
Síntomas del ataque, hongos asociados a los daños ocasionados y estados de desarrollo de C. zulmae en árboles de Aliso. A. Exudado. B. Aserrín. C. Perforación vieja. D. Hongos fitopatógenos asociados a las perforaciones. E. Micelio presente en el interior de las galerías de oviposición. F. Huevo. G. Larva recién emergida. H. Pupa. I. Adulto.
Según la prueba de Duncan (P<0,05), el mayor promedio para la variable número de perforaciones se presentó en diciembre (valores mínimos de 2 y máximos de 159 perforaciones por árbol); aunque se observaron diferencias significativas para las variables número y altura de perforaciones por árbol, no se pudo establecer el comportamiento de dichas variables a lo largo del tiempo evaluado. El ataque del insecto en el tronco se concentró entre los 127 y 198 cm sobre el suelo (Tabla 1); sin embargo, se hallaron perforaciones a alturas por debajo de este intervalo hasta los 21 cm y por encima hasta 575 cm.
Número promedio de perforaciones por árbol afectado y altura de los daños causados por C. zulmae en la finca La Fe, municipio de Villamaría, Caldas.
Medias con la misma letra no difieren estadísticamente entre sí, de acuerdo con la prueba de comparación Duncan al 5%. E.E.= Error estándar.
No se encontraron diferencias en el número de huevos, larvas, pupas y adultos a lo largo del año. A pesar de que el Aliso fue atacado por C. zulmae, se observó que una baja proporción de los árboles plantados contenían estados del insecto (35%, N=147). El 9,72% de las perforaciones evaluadas (N=1634) presentaron al menos un estado, observándose máximo 10 individuos por galería. Posiblemente estos árboles vivos con contenidos altos de humedad no proporcionaron las condiciones necesarias para la reproducción de la especie. Las observaciones en troncos del suelo que provenían de árboles talados en meses anteriores, demostraron que C. zulmae prefirió alimentarse y reproducirse sobre Alisos decadentes en el suelo; este material presentó estados del insecto en mayor proporción (90%, N=22) comparado con los muestreos destructivos, explicando la baja proporción de individuos presentes en los árboles plantados. Estos resultados son acordes con Wood (1982) y Cibrian et al. (1995) quienes reportan que los adultos de este género perforan el fuste y las ramas de los árboles vivos débiles o derribados, y además que el tejido de estos árboles recién cortados o muertos emite compuestos volátiles que atraen a ambos sexos de su propia especie.
Durante todo el año se hallaron 408 adultos en los muestreos de árboles plantados; 194 de los cuales fueron hembras y 214 machos. Esta misma relación de sexos (1:1) se presentó en el seguimiento realizado al material del suelo durante julio y octubre del 2009, periodo en el cual se obtuvieron 414 adultos (195 hembras y 219 machos).
En todas las perforaciones de C. zulmae se observaron hongos de coloración marrón que se extendían a lo largo afectando los haces vasculares (Fig. 2D). El interior de las galerías que presentaron cámaras de oviposición con larvas, pupas y adultos, se encontraron invadidas por un micelio gris que cubría los estados inmaduros del insecto (Fig. 2E). Estas observaciones sugieren que C. zulmae podría ser un insecto ambrosial, al igual que lo reportan para las diferentes especies de este género (Giese 1967; Cibrian et al. 1995). Gil et al. (2004), aislaron e identificaron en el interior de las galerías a Fusarium solani, (Mart.) Sacc. Fusarium sp., Verticillium sp, y una levadura cercana a las especies del género Pichia sp. En el avance de la lesión se aislaron F. solani y Ceratocystis sp. indicando que el ataque de C. zulmae en asociación con Ceratocystis sp. ocasionaba la muerte de los árboles y que los hongos del género Fusarium eran invasores secundarios. Considerando que los hongos fueron aislados de tejido afectado asociado a las heridas hechas por los insectos en el tallo y no directamente del micelio presente en el interior de las cámaras de cría que contenían estados inmaduros del insecto, no se podría asegurar que los hongos fitopatógenos reportados por Gil et al. (2004) correspondieran al microorganismo simbionte.
Se registraron ocasionalmente especies coexistentes con C. zulmae en los muestreos. Se encontró un insecto de la misma subfamilia (Scolytinae) cercano al género Amphicranus ocasionando perforaciones en árboles muertos en pie, pero tanto en los muestreos como en las trampas de alcohol, C. zulmae fue la especie más abundante (75%). Asociado al daño del insecto, en galerías recientes, se observaron larvas de un díptero de la familia Richardiidae y en galerías viejas se encontraron ácaros.
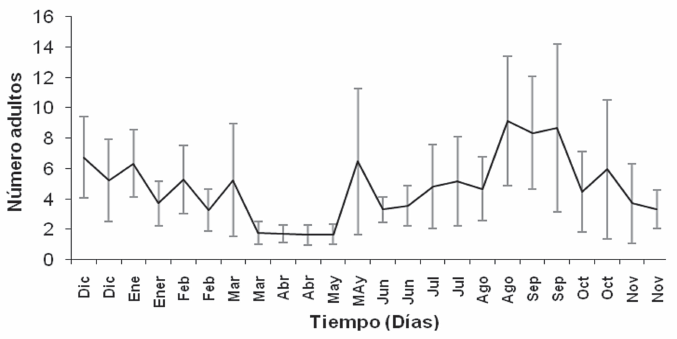
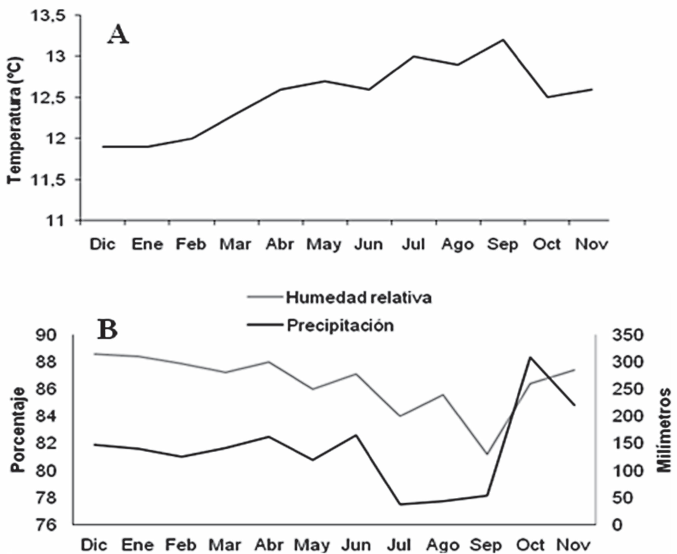
Las trampas de captura indicaron que hay individuos de C. zulmae durante todo el año, con marcada tendencia al aumento en agosto - septiembre, seguido de diciembre - enero y mayo. La Figura 3 muestra diferencias significativas entre los meses mencionados anteriormente y el periodo comprendido entre marzo y abril donde se dieron las menores capturas. Con relación a las variables del clima se observó que las temperaturas más altas se registraron en el periodo de agosto y septiembre (Fig. 4A) donde se dieron las mayores capturas de insectos, sin embargo, los meses donde la población de individuos fue menor, no muestran una diferencia marcada de la temperatura que pueda explicar el comportamiento poblacional de la especie durante este periodo. La humedad relativa y la precipitación (Fig. 4B) presentaron una tendencia similar, y a pesar que no fue posible establecer una relación directa entre las capturas del insecto y las variables de clima, pareciera que los periodos de sequía pudieran favorecer la dispersión de C. zulmae.
Promedio del número de adultos de C. zulmae capturados por trampa a través del tiempo, en 30 trampas atrayentes de alcoholes en el lote experimental de la finca La Fe, municipio de Villamaría, Caldas. Barras de error = intervalo de confianza 95%.
A. Temperatura promedio, B. humedad relativa promedio y precipitación acumulada del lote experimental de la finca La Fe del municipio de Villamaría, Caldas.
La mayor cantidad de adultos capturados en las trampas de alcohol fueron machos (2569 individuos) comparados con las capturas de hembras (756 individuos). Gil et al. (2004) y Wood (2007) reportan que las hembras de C. zulmae poseen antenas de clava larga con el margen anterior ornamentado por un grupo de setas, mientras que el macho no posee este grupo y las antenas son más cortas. Esta diferencia morfológica podría explicar la mayor atracción de machos a alcoholes y permitiría hipotetizar que las hembras tendrían una mayor capacidad para identificar volátiles específicos del huésped. Estudios realizados en algunas especies como C. columbianus y otros escolítidos han demostrado que las trampas de alcohol son selectivas a machos (Roling y Kearby 1975).
No fue posible establecer la relación entre el número acumulado de adultos capturados a través del tiempo y el número de estados encontrado en los muestreos en los árboles plantados, dado que se encontró un número muy bajo de poblaciones del insecto en éstos.
Estimación del tiempo del ciclo de vida de C. zulmae. Se realizó el seguimiento a 54 individuos de C. zulmae a partir de los cuales se estimó preliminarmente la duración de cada uno de los estados. Durante el desarrollo de esta actividad se estudiaron todos los estados de vida de C. zulmae. Los huevos son blanquecinos translúcidos, de forma elíptica, cuyo tamaño fue de 0,73 mm de largo y 0,46 mm de ancho aproximadamente (Fig. 2F) y son depositados de manera individual en cada galería; los demás estados observados corresponden con lo descrito por Gil et al. (2004) (Figs. 2G- I). Larvas blanquecinas, ápodas, en forma de "C" con mandíbulas esclerotizadas; pupas de color crema, típicamente exarata, las cuales, a medida que se desarrollaron, comenzaron a oscurecer sus alas posteriores y cabeza; los adultos recién emergidos presentaron una coloración marrón en todo el cuerpo; cuando completaron su crecimiento tomaron la coloración típica de la especie.
No fue posible estimar la duración del ciclo completo del insecto desde huevo hasta adulto, ya que solo 13 de los individuos observados pasaron al estado biológico siguiente, tiempo después del cual murieron. Pero se pudo estimar la duración promedio de huevos, larvas, pupas y adultos de manera independiente (Tabla 2). El 81% de los huevos no eclosionaron; el 71% de las larvas murieron y el 67% de las pupas no se desarrollaron completamente. El 33% de los adultos restantes que se obtuvieron a partir de pupas se les realizó seguimiento para estimar la duración del estado. Los resultados sugieren que los factores de humedad (70%) y temperatura (17,7°C) suministrados en laboratorio no fueron adecuados para el desarrollo del insecto o que la manipulación del hospedante afectó su alimentación. Wood et al. (1992), anotan que los insectos permanecen gran parte de su ciclo de vida dentro de las galerías hechas en las plantas hospedantes, donde las condiciones de microclima son constantes. Esto es corroborado por Kabir y Giese (1966), quienes mencionan que las larvas maduras de C. columbianus prefieren permanecer dentro de una cámara cerrada para poder completar su desarrollo. Se sugiere que esta evaluación se realice en condiciones naturales.
Duración promedio de los estados de C. zulmae observados en laboratorio.
Estado
Número de Observaciones
Duración Promedio (Días)
E.E.
Min. (Días)
Max. (Días)
Huevo
38
12,16
0,89
3
22
Larva
14
7,14
1,20
2
20
Pupa
11
9,36
1,00
1
13
Adulto
4
19,75
9,17
8
47
E.E.= Error estándar.
Se pudo establecer que la duración promedio del ciclo del insecto podría superar los 48 días y se requerirían más de 28 días para que los huevos se desarrollen hasta el estado adulto (Tabla 2). Los resultados sugieren que C. zulmae podría presentar más de dos generaciones al año en condiciones de campo, contrario a lo reportado para C. columbianus el cual tiene una duración de huevo de seis días aproximadamente, larva de 12 a 15 días, prepupa de dos a tres días, pupa de ocho a nueve días y los adultos de 34 a 40 días en condiciones de laboratorio, presentando dos generaciones por año (Kabir y Giese 1966).
Evaluación de un sustrato para la cría de c. zulmae. Se presentó alta mortalidad de los adultos de C. zulmae que conformaron las unidades experimentales por lo que se realizó un análisis descriptivo. los insectos comenzaron a perforar los sustratos pocos días después de infestados, mostrando tanto el macho como la hembra, capacidad de perforar y realizar galerías en los tratamientos. se presentó mayor proporción de sustratos perforados en el tratamiento de troncos de Aliso sobre algodón con un 70%. No se observó actividad de cópula de los insectos lo que indica que las condiciones necesarias para la fertilización probablemente ocurran durante la construcción de la galería.
Las primeras posturas se observaron a los 20 días después de la infestación, donde se obtuvieron 10 huevos de C. zulmae en el 1% de las unidades experimentales del tratamiento tres, allí el insecto realizó galerías con cámaras de oviposición en cada una de las cuales se encontraba depositado un huevo. Ningún otro sustrato presentó respuesta a la variable número de estados. Entre el 30 y el 50% de los adultos habían muerto en los diferentes sustratos. A los 40 días se presentó entre un 45 y un 70% de mortalidad de adultos, encontrándose reproducción del insecto en dos tratamientos, en el tratamiento dos se obtuvo un huevo y una larva muerta en una de las repeticiones, y en el tratamiento tres, cuatro larvas vivas y cuatro larvas muertas en dos de las repeticiones. En la última evaluación realizada a los 60 días se observaron muertos entre el 70 y el 100% de los adultos con los que se infestaron los tratamientos y únicamente ocho adultos muertos en una de las unidades experimentales del sustrato de ramas de Aliso. A pesar de que el insecto pudo ovipositar en algunos sustratos, ninguno de los tratamientos fue adecuado para la cría, dada la baja cantidad de estados encontrados. El desarrollo de la cría de un insecto ambrosial depende del almacenamiento de hongos simbiontes en las micangias y de su crecimiento en el sistema de galerías, asegurando alimento para la progenie (Francke-Grosmann 1967; Kuhnholz et al. 2001; Mizuno y Kajimura 2002; Biedermann 2007). Es posible que los métodos de esterilización usados en esta investigacion tanto para los sustratos como para los insectos, hayan limitado el establecimiento del hongo ambrosial del cual se alimentan las larvas de C. zulmae en las dietas artificiales, sugiriendo además que sus micangias podrían ser de origen ectodermal. Estudios posteriores para desarrollar dietas apropiadas para el crecimiento y reproducción de C. zulmae requieren de la identificación de micangias en esta especie y de la determinación del hongo ambrosial del cual se alimentan sus estados inmaduros. Igualmente se sugieren evaluar diferentes condiciones de humedad y temperatura.
Conclusiones
Corthylus zulmae no parece ser el responsable de la muerte de árboles de Aliso sanos; al igual que muchos xilomicetófagos, este insecto puede ser atraído por los árboles cuando las condiciones climáticas son desfavorables o cuando se encuentran senescentes o muertos. Durante este proceso se presentan hongos fitopatógenos oportunistas asociados a los daños mecánicos que debilitan los árboles y en condiciones extremas ocasionan la muerte del Aliso. El adecuado manejo de los residuos presentes en el suelo y la eliminación de los árboles en mal estado fitosanitario, podrían disminuir las poblaciones del insecto en plantaciones de Aliso.
Footnotes
Agradecimientos
Los autores expresan sus agradecimientos a Carlos Augusto Ramírez, Adriana Cifuentes y Freddy Sánchez de la disciplina de ETIA, al personal de la Disciplina de Entomología e Ingeniería Agrícola de Cenicafé. A los Ingenieros Jorge Uribe Jaramillo, Carlos Enrique Restrepo, al Señor Darío Murillo de la empresa Aguas de Manizales y a los funcionarios de Procuenca, por su colaboración en el desarrollo de este experimento. Esta investigación se realizó en el Centro Nacional de Investigaciones de Café con la cofinanciación del Ministerio de Agricultura y Desarrollo Rural.
References
1.
BEAVERR.A.1989. Insect-fungus relationships in the bark and ambrosia beetles. In: WildingN., CollinsN. M., HammondP. M. and WebberJ. F. (eds.), Insect-fungus interactions. London: Academic Press. pp. 121–143.
2.
BIEDERMANNP. H. W.2007. Social behavior in sib mating fungus farmers. Intra- and interspecific cooperation in ambrosia beetles. Masterarbeit der Philosophisch-naturwissenschaftlichen Fakultät der Universität Bern vorgelegt von. 85 p.
3.
BRICEÑOV. A.2002. El Aliso, Alnus acuminata H.B.K., hospedero de dos especies de Chrysomelidae (Coleoptera) en los andes venezolanos. Revista Forestal Venezolana46 (1): 53–57.
4.
CIBRIANT. D.; MENDEZM. J. T.; CAMPOSB. R.; YATESH. O.; FOLRESL. J.1995. Insectos Forestales de México. Universidad Autónoma de Chapingo, Chapingo, Estado de México, México. 382 p.
5.
COMSTOCKJ. H.1962. An introduction to entomology. Comstock Publishing associates a division of Cornell University press. Ithaca, New York. 1064 р.
6.
DEL VALLEJ. I.; GONZÁLEZH.1988. Rendimiento y crecimiento del cerezo (Alnus jorullensis) en la región central andina, Colombia. Revista Facultad Nacional de Agronomía. Medellin41(1): 61–90.
7.
FARRELLB. D.; SEQUEIRAA. S.; O'MEARAB.; NORMARKB. B.; CHUNGJ.H.; JORDALB. H.2001. The evolution of agriculture in beetles (Curculionidae: Scolytinae and Platypodinae). Evolution55: 2011–2027.
8.
FINNEGANR. J.1967. Notes on the Biology of the Pitted Ambrosia Beetle, Corthylus punctatissimus (Coleoptera: Scolytidae), in Ontario and Quebec. The Canadian Entomologist99: 49–54.
9.
FRAEDRICHS. W.; HARRINGTONT. C.; RABAGLIAR. J.; ULYSHENM. D.; MAYFIELDA. E.; HANULAJ. L.; EICKWORTJ. M.; MILLERD. R.2008. A fungal symbiont of the redbay ambrosia beetle causes a lethal wilt in redbay and other Lauraceae in the southeastern United States. Plant Disease92: 215–224.
10.
FRANCKE-GROSMANN. 1967. Ectosymbiosis in wood-inhabiting insects. In HenryS. M. (ed.), symbiosis. New York. Vol. II. p 141–205.
11.
FURNISSM. M.; WOOJ. Y.; DEYRUPM. A.; ATKINSONT. H.1987. Prothoracic Mycangium on pine-infesting Pityoborus spp. (Coleoptera: Scolytidae). Annals of the Entomological Society of America80: 692–696.
12.
GIESER. L.1967. The Columbian Timber Beetle, Corthylus columbianus (Coleoptera: Scolytidae). V. A description of the mycetangia. The Canadian Entomologist99: 54–58.
13.
GILP. Z. N.; BUSTILLOP. A. E.; GOMEZD. E.; MARINM. P.2004. Corthylus n. sp. (Coleoptera: Scolytidae), plaga del aliso en la cuenca de río Blanco en Colombia. Revista Colombiana de Entomología30(2): 171–178.
14.
HARRINGTONT.C.2005. Ecology and evolution of mycophagous bark beetles and their fungal partners. Pages 257–291 In: Ecological and Evolutionary Advances in Insect-Fungal Associations, VegaF. E. and BlackwellM., eds. Oxford University Press.
15.
HOLDRIDGEL.1987. Ecología basada en zonas de vida. IICA: 216 р.
16.
KABIRA. K. M. F.; GIESER. L.1966. The Columbian Timber Beetle, Corthylus columbianus (Coleoptera: Scolytidae). I. Biology of the Beetle. Annals of the Entomological Society of America59 (5) 883–894.
17.
KUHNHOLZS.; BORDENJ. H.; UZUNOVICA.2001. Secondary ambrosia beetles in apparently healthy trees: adaptations, potential causes and suggested research. Integrated Pest Management Reviews6: 209–219.
18.
MIZUNOT.; KAJIMURAH.2002. Reproduction of the ambrosia beetle, Xileborus pfeili (Ratzeburg) (Col., Scolytidae), in semiartificial diet. Journal of Applied Entomology126: 455–462.
19.
NORMARKB. B.; JORDALB. H.; FARRELLB. D.1999. Origin of a haplodiploid beetle lineage. Proceedings of The Royal Society Biological Sciences266: 1–7.
20.
OSPINAP. C. M.; HERNANDEZR. R. J.; GOMEZD. D. E.; GODOYB. J. A.; ARISTIZABALV. F. A.; PATIÑOC. J. N.; MEDINAO. J. A.2005. Serie cartillas divulgativas. El Aliso o Cerezo Alnus acuminata H. B. K. ssp. acuminata. Guías silviculturales para el manejo de especies forestales con miras a la producción de madera en la zona andina Colombiana. FNC-Cenicafé. 36 p.
21.
PORTILLAR. M.; STREETTD.2006. Nuevas técnicas de producción masiva automatizada de Hypothenemus hampei Ferrari sobre la dieta artificial cenibroca modificada. Revista Cenicafé57(1):37–50.
22.
POSADAL. O1989. Lista de insectos dañinos y otras plagas en Colombia. Instituto Colombiano Agropecuario ICA, Bogotá, Boletín Técnico No. 43. 662 p.
23.
ROLINGM. P.; KEARBYW. H.1975. Seasonal flight and vertical distribution of scolytidae attracted to ethanol in an oak-hickory forest in Missouri. The Canadian Entomologist107: 1315–1320.
24.
STEINHAUSE. A.1947. Insect Microbiology. Comstock Publishing Company. Ithaca, New York. 763 p.
25.
TANADAY.; KAYAH. K.1993. Insect pathology. Academic Press, inc. San Diego. Estados unidos. 666 p.
26.
WOODS. L.1982. The bark and ambrosia beetles of North and Central America (Coleoptera: Scolytidae) a taxonomic monograph. Great Basin Naturalist Memoires (Utah-EE.UU). (6) 1359 p.
27.
WOODS. L., STEVENSG. C., LEZAMAH. J.1992. Los Scolytidae (Coleoptera) de Costa Rica: Clave de la subfamilia Scolytinae, Tribu Cothylini. Revista de Biología Tropical40 (3): 247–286.
28.
WOODS. L.2007. Bark and ambrosia beetles of South America (Coleoptera: Scolytidae). M. L. Bean Life Science Museum. Brigham Young University. (Utah-EE.UU) 842 р.


 (13%), segundo
(13%), segundo  (29%), tercero
(29%), tercero  (33%), cuarto
(33%), cuarto  (36,5%) y quinto censo
(36,5%) y quinto censo  (38,6%) y la ubicación de las trampas de alcohol
(38,6%) y la ubicación de las trampas de alcohol  . El porcentaje acumulado de infestación se indica entre paréntesis.
. El porcentaje acumulado de infestación se indica entre paréntesis.