
Abstract
The stages of post-embryonic development of
Introducción
Los gérridos están ampliamente distribuidos en los ecosistemas acuáticos lóticos y lénticos; en general están asociados a la película superficial del agua. En Colombia se han publicado varios estudios taxonómicos sobre los gérridos, entre estos los de Aristizabal (2002), Padilla y Nieser (2003), Polhemus y Polhemus (1995) y trabajos como los de Morales-Castaño y Molano-Rendón (2009) en relación con el género
La especie objeto de estudio es
El objetivo de este trabajo es determinar la variación temporal y altitudinal de
Materiales y Métodos
Área de estudio
Se muestrearon seis localidades, ubicadas en cuatro municipios: Pasto, Buesaco, Funes y Tangua (Fig. 1). Todas las localidades presentan un régimen de precipitación bimodal, la época más seca comprende los meses de junio a septiembre, la más lluviosa son octubre y noviembre (IGAC 1985). A continuación se citan las localidades de muestreo, y las fechas en las cuales se realizaron los muestreos, durante el 2009.
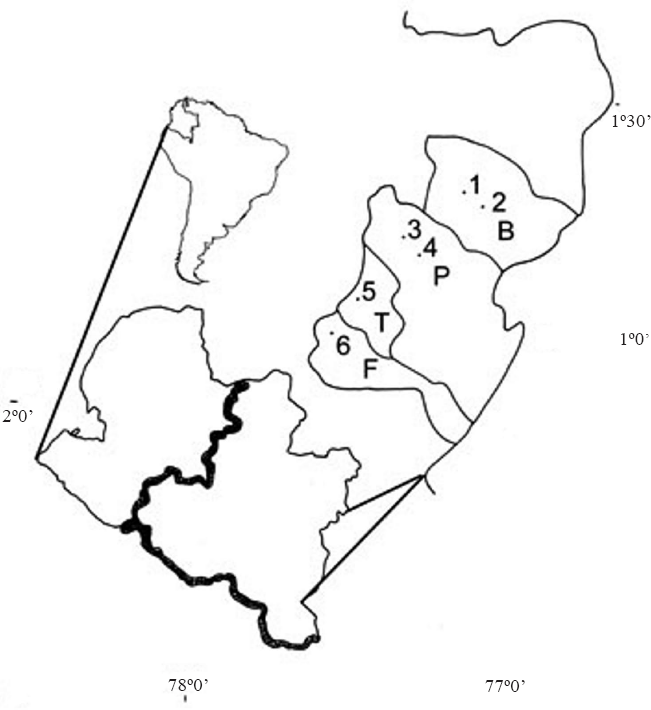
Localización de las poblaciones estudiadas de
En el Municipio de Pasto, 1°10’12”N -77°12’36”W, las localidades: Daza y San Fernando, con una altitud de 2700 m y 2640 m, respectivamente. Daza: Junio 30, Octubre 12; San Fernando: Junio 17, Noviembre 11. En el Municipio de Buesaco, 1°23’38”N -77°08’54”W, las localidades: Villamoreno y Rosal el Monte, con una altitud de 2330 m y 2420 m, respectivamente. Junio 19, Octubre 12.
En el Municipio de Funes, 0°48’-1°03’N-77°53-33’W, Vereda La Cocha a 2340 m de altitud. Junio 3, Noviembre 2. En el Municipio de Tangua, 3°19’N-77°24'W, Vereda El Porvenir a 2260 m de altitud. Junio 24, Noviembre16.
Muestreo
Cada localidad se visitó dos veces, una en época seca y otra en época lluviosa. Los muestreos se realizaron en las horas de la mañana. Se colectó el material con una red. Cuatro colectores trabajaron simultáneamente en cada sitio de muestreo. Los especímenes fueron transportados al laboratorio en frascos con etanol al 70%, donde posteriormente se procesaron y determinaron cada uno de los estadios. Los adultos fueron sexados y separados en ápteros y macrópteros (con ala completa); los estadios fueron asignados con base en las medidas del largo total y el ancho del pronoto.
Todos los especímenes examinados para el estudio de su desarrollo postembrionario proceden de la localidad de Funes, único sitio donde se observaron y colectaron huevos. En la descripción de los estadios ninfales se utilizan las siguientes abreviaturas: largo y ancho: L/A; largo total: Lt; ancho de la cabeza, a través de los ojos: Ac; ancho del pronoto, en su parte más ancha: Ap; ancho máximo del mesonoto: Am; abdomen largo total y ancho máximo a través del conexivo: abdomen L/A; ninfa hembra: Nh; ninfa macho: Nm. Todas las medidas en milímetros.
Para la comparación entre poblaciones se realizó un conteo total de los especímenes por estadio ninfal sin tener en cuenta el sexo de los estadios IV y V. Todos los adultos ápteros y macrópteros se contaron. Las abreviaturas adoptadas para cada estadio son: Ma, macho áptero; Ha, hembra áptera; Mm, macho macróptero; Hm, hembra macróptera; estados ninfales n1-n5. Todos los especímenes fueron depositados en la colección de entomología de la Universidad de Nariño (PSO-CZ).
Se evaluó el efecto de altitud y la estacionalidad frente al número de individuos según su estadio de desarrollo, adultos ápteros macho y hembra, adultos macrópteros macho y hembra, y cinco estadios ninfales. Se aplicó el análisis de varianza ANOVA/MANOVA, mediante el software Statgraphics (2000).
Resultados
Desarrollo post-embrionario
Se caracteriza el desarrollo post-embrionario de la forma áptera de
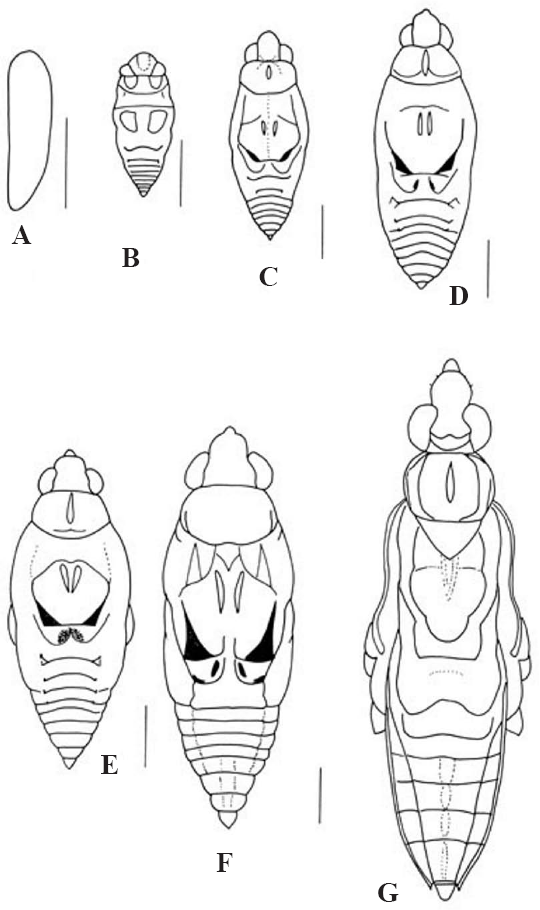
Vista dorsal de los estadios de
En los estadios IV y V se acentúa la coloración castaño oscuro mesopostnotal, en el metanoto, y el delineamiento castaño oscuro de los segmentos del abdomen dorsal.
Dimorfismo sexual: la Nh es un poco más robusta que la Nm y el abdomen de la Nm es más angosto. El conexivo de la Nm es ligeramente levantado en los segmentos I-IV y en la Nh es horizontal; el esterno abdominal VII tanto de la Nm como de la Nh es igual de largo pero en la Nh es más ancho; el esterno abdominal VIII en la Nh presenta una línea media difusa y la Nm carece de ésta.
Caracterización del hábitat
En varias localidades de Buesaco, no consideradas para este estudio, de Rosal del Monte y Vereda Pajajoy,
En las localidades de Daza, Villamoreno, Rosal del Monte y Tangua también se observaron especímenes de
Análisis comparativo de las poblaciones
Se colectaron y caracterizaron 1342 individuos, 831 adultos y 511 ninfas de los diferentes estadios ninfales de las seis localidades. El número total de individuos por época en cada una de las localidades se indica en la Tabla 1. La localidad donde se colectó el mayor número de individuos fue Rosal del Monte (334) y aquella con el menor número de individuos fue Daza (106). No hubo diferencias en la abundancia de los estadios de
Número total de individuos, ninfas y adultos capturados de
Los estadios por localidad presentaron diferencias significativas (P < 0,05), para cuatro localidades: San Fernando (F = 39.75), Villamoreno (F = 7,04), Rosal del Monte (F = 10,29), y Funes (F = 10.85). Las dos localidades que no presentaron diferencias significativas fueron Daza y Tangua (P > 0,05). La altitud no marcó diferencias significativas (F = 1,91, P > 0,05).
En San Fernando, los estadios con diferencias significativas fueron los adultos ápteros machos y hembras (Fig. 3A); en Villamoreno las diferencias las presentaron los estadios adultos ápteros machos, las hembras ápteras y el estadio ninfal 1 (Fig. 3B); en Rosal del Monte se presentaron cuatro grupos, los adultos ápteros machos, las hembras ápteras, los adultos macrópteros machos y hembras, y el resto de estadios ninfales (Fig. 3C). En Funes, se presentan tres grupos, machos ápteros, hembras ápteras y el resto de los estadios (Fig. 3D).
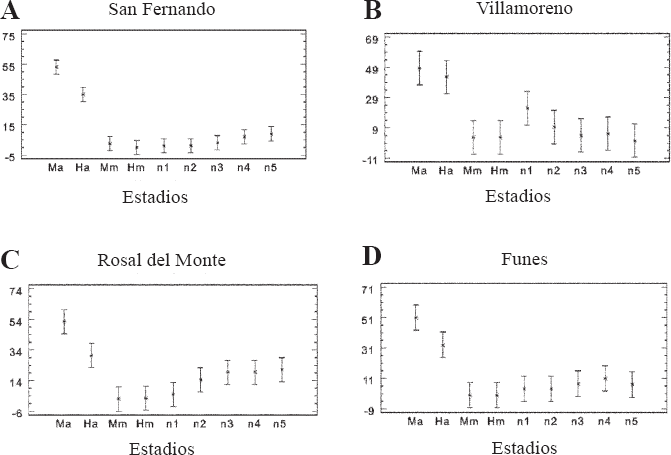
Variación del número de individuos por estadio de
Es de señalar que en Daza y Funes no se registraron adultos macrópteros y en San Fernando y Rosal del Monte, se colectaron sólo en época lluviosa; en las otras dos localidades predominaron en época lluviosa.
En la época seca, en Tangua los estadios ninfales uno y dos y en Funes, el estadio ninfal uno, no se registraron. Predominan los machos ápteros con más del 55% (55-79%) para casi todas las localidades excepto para Villamoreno, en época lluviosa, donde dominaron las hembras ápteras con un 66% y Tangua, también en época lluviosa, con el 52% de hembras ápteras. En época seca, se colectaron cuatro individuos macrópteros en Villamoreno (dos) y en Tangua (dos). En época lluviosa los machos de la forma macróptera fueron levemente más frecuentes con el 51% que las hembras 49%.
Discusión
En Argentina Mazzucconi y Bachmann (1993a, 1993b) registraron formas micrópteras para
La presencia de mayor número de individuos alados en la época lluviosa, probablemente está relacionada con la dispersión de la especie, a través de la colonización de nuevas charcas temporales, formadas en época de mayor precipitación. Por otra parte como señala Andersen (1982) el polimorfismo alar puede ser favorecido, si los hábitats preferidos son una mezcla de sitios permanentes y temporales, tal parece ser el caso de
El rango altitudinal en el cual se colectaron los especímenes (2.260-2.700 m) no marco una diferencia significativa en las poblaciones. Se amplió el rango altitudinal, indicado para esta especie por Molano-Rendón
No hubo diferencias significativas entre las épocas seca y lluviosa; también se observó el predominio de adultos ápteros y la presencia de todos los estadios ninfales. Esto lejos de ser resultados concluyentes debido al muestreo, nos permiten verificar la existencia de estas poblaciones y conocer algunos aspectos sobre su estado actual espacio-temporal e igualmente sirven como punto de partida para el planteamiento de otros interrogantes sobre la biología y ecología de esta especie como son: cuáles son las características y duración del ciclo de vida?, cuántos huevos son ovipositados por hembra y en qué lapso?, se mantienen las frecuencias de emergencia de los adultos formas ápteras y macrópteras por población/año?, qué factores ambientales influyen para el mantenimiento de las poblaciones a través del año?, las poblaciones presentan suficiente cantidad de adultos y estadios ninfales para asegurar su continuidad temporal y espacial?, qué factores inciden para mantener la abundancia de los distintos estadios?
Footnotes
Agradecimientos
Agradecemos a los estudiantes G. Montenegro y N. Nicola, por su ayuda en las salidas de campo. A la Vicerrectoría de Investigaciones (VIPRI) de la Universidad de Nariño por financiar esta investigación. A Carlos E. Sarmiento, editor de la revista y a dos revisores anónimos por la revisión del manuscrito.