
Abstract
Transgenic plants possessing genes of the bacteria Bacillus thuringiensis (Bt) which codify for toxin production effective against some pests, are considered useful within the management of insects. However, these commercial cultivars do not control satisfactorily all noctuids, and besides, at a distance, induce resistance. Diverse populations of native Heliothis virescens, Helicoverpa zea, Spodoptera frugiperda and S. sunia strains were evaluated to determine the susceptibility to the toxin Cry1Ac of the cultivar Bollgard®, planted in Colombia. Cry1Ac was obtained from a Bt gene cloned in Escherichia coli and from MVP® (protoxin encapsulated in Pseudomonas). Neonate larvae were exposed to seriated dose of the toxin, incorporated within meridic diet and also fed with fresh transgenic cotton tissue. Weight and adult emergency of the survivors were determined. Results were submitted to an Anova and LC was obtained by Probit analysis. An acceptable susceptibility of H. virescens, LC50 of 3.52 and 3.81 µg/mL, 2005A y 2006A, respectively, and a 100% mortality when feeding neonates with terminal leaves was found. LC50 for H. zea varied from 3.42 to 6.12 µg/mL; as the toxin dose increased, the pupal weight decreased and a high percentage of deformed pupae were observed. For Spodoptera spp., LC50 oscilated between 192 y 1.178 µg/mL, showing its resistance to the toxin. It was concluded that, at the moment, the transgenic cotton provides satisfactory control of the Heliothine, but no of the Spodoptera complex.
Introducción
Desde que el hombre se volvió sedentario y se dedicó a la agricultura intensiva en forma de monocultivos en extensas áreas, le ha tocado luchar contra las plagas, con énfasis en los insectos herbívoros. Al encontrar algún método de control siempre pensó haber ganado la batalla, lo cual fue especialmente cierto con el advenimiento del control químico. La destrucción de la fauna benéfica y el surgimiento de resistencia a los insecticidas, logró hacer recapacitar y formular nuevos conceptos que finalmente se concentraron en un manejo integrado de las plagas. Este manejo tampoco ha sido aceptado por muchos agricultores por requerir atención especial a los cultivos, monitoreo y a menudo es considerado demasiado costoso y dispendioso. Dentro de los programas de MIP, el último “método de control” incorporado ha sido el uso de cultivares “insecticidas” transgénicos que contienen proteínas Cry de la bacteria del suelo Bacillus thuringiensis Berliner, 1915 (Bt), las cuales protegen las plantas contra la alimentación de larvas de algunas plagas del orden Lepidoptera (Gustafson et al. 2006; Carrière et al. 2001).
Las plantas transgénicas, entre ellas el algodonero (Gossypium hirsutum L., 1793) que poseen genes Bt que codifican para la producción de toxinas Cry, eficaces contra algunas plagas, son por lo tanto consideradas útiles como herramientas dentro del MIP. Pueden evitar aplicaciones de insecticidas químicos contra algunos nóctuidos, los cuales deterioran el ambiente, pero no controlan ácaros, ni áfidos o moscas blancas. El control de las plagas objetivo con las variedades transgénicas y la consecuente ausencia del control químico pueden resultar en el aumento de poblaciones de estas plagas secundarias. Así, se cambia el panorama, se presentan problemas nuevos y difí-ciles; ante todo existe luego la posibilidad de que los insectos chupadores adquieran mayor importancia y su manejo aumente el costo de producción, ya incrementado por el valor de la semilla del cultivar transgénico (ICAC 2005).
El gene Cry1Ac introducido al algodonero Bollgard (Monsanto Company, St. Louis, MO) que se siembra desde el año 2003 en Colombia, expresa cristales proteicos con actividad insecticida contra algunos lepidópteros, especialmente los belloteros. Esta variedad, de acuerdo con Jackson et al. (2003), provee control absoluto en los estados productores de algodón de los Estados Unidos contra Heliothis virescens (F., 1777), pero muestra ocasionalmente problemas de eficacia contra Helicoverpa zea (Boddie, 1850). Esta toxina no ejerce controles satisfactorios de Spodoptera frugiperda (Smith, 1797) (Garczynski et al. 1991; Bohorova et al. 1997; Ayra-Pardo et al. 2006), nóctuido común en el algodonero en nuestro medio.
A nivel mundial, una de las preocupaciones asociadas al algodonero Bt corresponde a la posibilidad del desarrollo de resistencia de las poblaciones plagas objetivo, basado en selección por resistencia al Cry1Ac, particularmente para los belloteros H. virescens, H. zea y Helicoverpa armigera (Hübner, 1805) (Lepidoptera: Noctuidae) (Ali et al. 2006; Jackson et al. 2007; Rausher 2001; Bird y Akhurst 2005). Además, el manejo de la resistencia se ha vuelto mucho más complejo y costoso, debido a que los refugios se establecen de manera mandatoria y existen consideraciones estratégicas de selección múltiple de cultivos transgénicos a través de las regiones agrícolas (Ali et al. 2006).
Antes de la comercialización de los cultivos transgénicos aparecieron las preocupaciones y los llamados de atención sobre la evolución de resistencia por parte de los insectos plagas al B. thuringiensis. En 1985, se alertó a la comunidad internacional sobre el riesgo de desarrollo de resistencia de insectos al Bt (McGaughey 1985). Bajo condiciones de laboratorio ya se seleccionaron hace algunos años razas que pueden cumplir su ciclo biológico en cultivos transgénicos (Tabashnik et al. 2003). Al trabajar con la raza HD-1 y una formulación comercial del B. thuringiensis ssp. kurstaki, Stone et al. (1989) demostraron el potencial del H. virescens para desarrollar resistencia a la delta - endotoxina del Bt. Presión de selección, ejercida durante cuatro generaciones en el laboratorio sobre una población de S. frugiperda con una formulación comercial de Bt (mezcla de cinco toxinas, incluyendo Cry1Ac) mostró un incremento de 4,6 veces en la CL50 (Borrero y Zenner de Polanía 1998).
Las siembras comerciales de algodón transgénico se iniciaron en 1996 en Estados Unidos y Australia. Durante este primer año se plantaron 1.800.000 acres en Estados Unidos (National Cotton Council 1997); James (2005) citado por Tabashnik et al. (2006) menciona que a nivel mundial en el año 2005, se sembraron 26 millones de hectáreas de algodón y maíz transgénico, área que podría permitir la adquisición rápida de resistencia a la toxina incorporada de las plagas clave. La tolerancia y resistencia de las plagas al Cry1Ac, entre otras causas, ocasionaron insatisfacción en EE.UU. y Australia, lo cual llevó a la construcción de una segunda generación del algodonero Bt, denominados algodones “gene-piramidales”: el Bollgard II que contiene los genes Bt que codifican la producción de las endotoxinas Cry1Ac y Cry2Ab, efectivas contra larvas de lepidópteros (Greenplate et al. 2003) y el Wide Strike (Dow AgroSciences) que produce las endotoxinas Cry1Ac y Cry 1F (Pellow et al. 2002, citado por Jackson 2007). El Cry1Ac incorporado al Bollgard todavía controla satisfactoriamente a Heliothis spp., pero no es eficaz contra otros lepidópteros plaga; el Cry adicional del Bollgard II es considerado también eficiente contra S. frugiperda, S. exigua (Hübner, 1808), Pseudoplusia includens (Walter, 1857) y Trichoplusia ni (Hübner, 1803) (ICAC 2004). De acuerdo con Haile et al. 2004, citados por ICAC (2005), el algodón Wide Strike, una variedad que también posee incorporados dos genes, aislados del Bt, el Cry1Ac y el Cry1F, muestra protección durante todo el desarrollo fenológico de la planta contra un amplio espectro de insectos del orden Lepidoptera, entre los cuales figuran los belloteros (complejo Heliothine), S. frugiperda, S. eridania (Cramer, 1782), S. exigua, Pectinophora gossypiella (Saunders, 1843), T. ni, P. includens y Agrotis ipsilon (Hufnagel, 1766). Las regulaciones de Monsanto prohíben comparaciones directas, presumiblemente en condiciones de campo, entre las variedades Bollgard, Bollgard II y WideStrikeTM (ICAC 2005); menciona en este contexto que, el desempeño en el rendimiento varía dependiendo del sitio, de la presión y del complejo de plagas presentes, incluso, si la variedad WideStrike™ se cultivara bajo las mismas condiciones.
Algodones transgénicos transformados para producir una novedosa proteína del Bt, la Vip3A, ya se están evaluando comercialmente en EE.UU. La Vip3A es considerada única, ya que muestra actividad tanto en su fase vegetativa como en la fase de esporulación, mientras que las actividades de las proteínas Cry1 y Cry2 se limitan a la fase de esporulación (Estruch et al. 1996). La proteína VIP es una exotoxina, también derivada de la bacteria del suelo B. thuringiensis, estructural, funcional y bioquímicamente diferente a las delta endotoxinas. Se expresa en todas las partes de la planta, incluyendo los componentes florales, para proteger la planta (ICAC 2004). Estudios recientes para evaluar la posibilidad de existencia de resistencia cruzada de razas resistentes de H. virescens al Cry1Ac al VIP3A, mostraron la ausencia de esta resistencia cruzada. La utilización de líneas de algodón que producen la proteína VIP, podría por lo tanto atrasar la evolución de la resistencia al Cry1Ac del bellotero (Jackson et al. 2007).
No todos los sistemas de producción de algodón son satisfactorios para el uso de variedades transgénicas y dadas las presiones competitivas del sector biotecnológico que desarrolla estos productos, es posible que en su diseño no se contemplen aspectos para el desarrollo de un cultivar óptimo (ICAC 2005). La razón de las fallas identificadas en algunos casos pueden estar en la presión de selección, en la variación en la expresión y actividad de la toxina por estacionalidad, en efectos de sitio, en desarrollo fenológico, en otros estresantes bióticos y abióticos y en efectos somaclonales (Halcomb et al. 1996).
Al formular este proyecto se desconocía literatura científica publicada en Colombia sobre los aspectos básicos y, a pesar del riesgo del desarrollo de resistencia, no se divulgó investigación sobre la susceptibilidad, expresada en concentración letal media (CL50 0 CL80), de S. frugiperda, H. virescens y H. zea al Cry1Ac incorporado al algodonero transgénico que se está sembrando en el país. Tampoco, se conocía la concentración de la toxina en las partes vegetativas y reproductivas de la planta a través de su desarrollo fenológico bajo nuestras diversas condiciones climáticas y edáficas, ni se publicaron recomendaciones validadas acerca del manejo de la variedad transgénica en cuanto a refugios y otros aspectos que podrían atrasar la posible adquisición de resistencia.
Por lo tanto, se propuso esta investigación para conocer la susceptibilidad de estas plagas al Cry1Ac y determinar simultáneamente la expresión de la toxina a través del desarrollo fenológico del algodón transgénico, para proponer a los cultivadores información concreta acerca del comportamiento y manejo de estas plantas transgénicas y del manejo de una posible resistencia. Conocer la línea base de susceptibilidad al Cry1Ac es indispensable para el monitoreo de la resistencia de los nóctuidos involucrados en este estudio y, posteriormente poder alertar a los agricultores y tomar medidas para el manejo de la resistencia, mientras que la expresión y concentración del gene que codifica la toxina a través del desarrollo fenológico de las plantas transgénicas puede ser crítico para el manejo de las plagas.
Materiales y Métodos
El Cry1Ac empleado en los bioensayos se obtuvo de un gene del Bt, donado por el Bacillus Genetic Sock Center (EE.UU.), extraído de la cepa ECE-53 de E. coli recombinante mediante la metodología recomendada por Akhurst et al. (2003) y de MVP® (Cry1Ac encapsulado en Pseudomonas), proporcionado por Dow CropSciences. Para establecer la concentración de la proteína purificada, se utilizó el método Bradford y para el desarrollo de la curva patrón, se emplearon diluciones seriadas con concentraciones entre 100-1.000 µg/mL de la solución comercial MVP, la cual tiene formulación líquida del 20% de la toxina Cry1Ac. Las lecturas se hicieron en un espectrofotómetro Spectronic 601 digital a una longitud de onda de 565nm.
El material biológico para las crías se recolectó en cultivos de algodón y maíz no transgénicos en los departamentos del Tolima, Casanare y Meta. La cría de H. zea se inició a partir de larvas de los dos últimos instares, obtenidas de mazorcas atacadas procedentes del Pie de Monte Llanero, municipio de Villavicencio. Se permitió su desarrollo en el laboratorio hasta pupa sobre pedazos de mazorca tierna, en vasos plásticos de 40 mL con tapa inyectada. Una vez obtenidas las pupas, se determinó su sexo y se colocaron en frascos grandes de vidrio para esperar la emergencia de los adultos. Éstos fueron alimentados con miel de abeja y para la oviposición se colocaron tiras de papel de color azul en el borde de los frascos. Con la descendencia parcial de diez parejas, aproximadamente 500 huevos, se inició la F1 a partir de la cual se mantuvo la cría masiva y se realizaron los bioensayos. Una metodología similar se empleó para H. virescens iniciándose la cría partiendo de huevos recolectados en hojas terminales en diversos sitios del Tolima y en Villanueva (Casanare) y alimentando las larvas con dieta importada específica para esta especie. Las larvas de S. frugiperda y de S. sunia (Guenée, 1852) fueron criadas a partir de posturas con dieta importada para luego seguir los mismos procedimientos.
La susceptibilidad de las larvas neonatas se evaluó incorporando concentraciones seriadas del Cry1Ac a dietas merídicas específicas (importadas de Southland Products INC., EE.UU.). Las dosis empleadas para H. virescens y H. zea oscilaron entre 0,001 y 100 µg/mL, mientras aquellas empleadas para las larvas neonatas del cogollero del maíz variaron de 0,001 a 1000 µg/mL, y de 0,002 a 2000 µg/mL. Todos los bioensayos se rigieron por un diseño de bloques al azar, seis repeticiones, cada una de ellas representada por un vaso plástico con dieta sobre la cual se colaron cinco larvas neonatas. La mortalidad se estimó a los siete días y los datos se sometieron a análisis Probit (SAS Institute Inc.) para determinar las concentraciones letales. El efecto de la alimentación con partes vegetales de plantas transgénicas sobre el desarrollo de las larvas de H. virescens y S. frugiperda se evaluó empleando en vez de la dieta con el Cry incorporado, tejido fresco transgénico. En siete repeticiones, diez larvas neonatas de la misma población por repetición, se colocaron sobre hojas terminales del algodón Bollgard; como testigo se emplearon hojas terminales de algodón convencional, sembrado en un refugio. Para algunas especies, se determinó con los sobrevivientes la duración del ciclo, el peso de las larvas, peso de las pupas y la emergencia de los adultos.
Para establecer la concentración del Cry1Ac, se tomaron, hojas y yemas terminales, botones florales, flores sin y con fecundación, sépalos, cápsulas, fibra y semillas, de un lote sembrado con semilla distribuida por “Cotton Seed Distributors”. Las partes, se liofilizaron y pulverizaron para luego tomar muestras homogéneas del material, llevarlo en 250 µL de buffer de extracción PBS (0,55%) y depositarlo en tubos de microcentrifuga por 24 horas en solución, para la determinación de la proteína Cry1Ac por la técnica de micro Elisa, empleando el kit “Envirologix Qualiplate para Cry1Ab/Cry1Ac”, kit diseñado para la detección cuantitativa para la proteina Cry1Ac en muestras de semillas y hojas de algodón Bollard®. La concentración, se midió utilizando un BioRad 'Benchmark' lector de microplacas (BioRad Laboratories; Tokio, Japón) a una longitud de onda de 450 nm. La lectura en línea de absorbancia se efectuó en un lector de Elisa Ref. Anthos 2020 (versión 2.0.5) en el laboratorio de investigaciones biotecnológicas de la Pontificia Universidad Javeriana. Todos los bioensayos se llevaron a cabo en el laboratorio de Biotecnología Agrícola de la Universidad de Ciencias Aplicadas y Ambientales U.D.C.A, bajo condiciones de temperatura promedia de 24,9°C y humedad relativa de 70%.
Resultados y Discusión
Concentración toxina Cry1Ac en el tejido vegetal
En la hojas cotiledonares, tejido de ocho días de edad, la concentración de proteina específica Cry1Ac fue de 4,625 ppm lo cual coincide con Bird y Akhurst (2005) quienes encontraron 5,6 ppm. Las concentraciones de allí en adelante, se muestran en la Tabla 1. En general, se observó una disminución de la concentración a medida que aumenta la edad de la planta, exceptuando la concentración obtenida en la flor fecundada. Según Olsen et al. (2005) a medida que el cultivo se desarrolla, la concentración de la proteína disminuye, por ende en este primer estado fenológico la planta expresará la mayor concentración de proteína Cry1Ac. Los cambios detectados, de acuerdo a Olsen et al. (2005), se pueden traducir en una disminución de la eficacia; se pueden deber a cambios en el nivel de expresión del gene y/o la constitución fisiológica de la planta y pueden ser inducidos por las condiciones ambientales.
Concentración máxima de proteína tóxica Cry1Ac en tejidos vegetales de Bollgard®.
Gusano bellotero del algodonero, Heliothis virescens
A las poblaciones, extremadamente altas y resistentes a todos los insecticidas empleados en su control, del gusano bellotero, se atribuye en parte las dos crisis algodoneras en Colombia siendo la plaga más importante durante los años 70s hasta comienzos de los 90s. A todos los insecticidas utilizados para su control, inclusive a los piretroides, que fueron la última arma esgrimida en forma masiva, mostró su capacidad para evolucionar a poblaciones resistentes. Luego, la plaga fue perdiendo importancia, su aparición se tornó esporádica y con bajas poblaciones, reduciéndose las aplicaciones para su control; llegando incluso a no reportar su presencia en algunas zonas del país durante una o más temporadas. En la actualidad, las poblaciones en los algodones convencionales, considerados “refugios”, son tan bajas o inclusive ausentes, por los menos en el departamento el Tolima, lo cual dificultó por un lado la obtención de muestras del insecto para los bioensayos y por el otro, la interpretación de los resultados. Durante el primer semestre del año 2007, las búsquedas de huevos de H. virescens en el área algodonera del municipio de El Espinal, por ejemplo, fue infructuosa, lo cual impidió la realización del bioensayo correspondiente.
El análisis Probit de los porcentajes de mortalidad, a los siete días, de las larvas de este bellotero recolectadas durante el semestre algodonero en el Tolima del año 2005, reveló una CL50 de 3,52 µg/mL, toxina total, (IC 95% 1,29 y 5,74 μg/ mL), una pendiente de la línea de regresión de 1,1655 y un R2 de 0,879. La CL80 correspondió a 6,09 µg/mL (IC 95% entre 3,87 y 8,32 µg/mL). Se observa una alta confiabilidad de estos resultados. Esto, desde el punto de vista práctico, significó un control satisfactorio de la plaga durante este semestre. En el bioensayo simultáneo con material vegetal fresco, hojas terminales del algodón Bollgard, en comparación con el testigo de algodón convencional, se encontró que al cabo de cuatro días el 100% de las larvas sometidas al algodonero transgénico murieron sin haber mudado al segundo instar, mientras que las larvas alimentadas con algodón convencional sobrevivieron y se desarrollaron normalmente.
Los resultados del bioensayo realizado con la población obtenida de un refugio de El Espinal del primer semestre del año 2006, con las mismas dosis seriadas incorporadas a dieta merídica, mostraron una CL50 de 3,81 µg/mL (IC 95% entre 1,36 y 6,14 µg/mL), una pendiente de la línea de regresión de 1,16741 y un Rβ de 0,917. La CL80 correspondió a 6,49 µg/mL (IC 95% entre 4,01 y 8,51 µg/mL). Nuevamente, se observó una alta confiabilidad de los resultados, lo cual, todavía, asegura un buen control de la población analizada. El aumento de la concentración letal entre los dos semestres fue insignificante, lo cual se atribuye a la presencia de huéspedes alternos del H. virescens, durante el semestre no algodonero.
Resultados no muy diferentes fueron obtenidos por Rodríguez Chalarcá et al. (2007) al establecer la línea base para el bellotero para tres poblaciones del Valle del Cauca y una de la costa norte del país. Las muestras del insecto procedentes de Buga, Palmira y Bugalagrande arrojaron, respectivamente, una DL50 de 3,2; 5,0 y 10,0 µg/mL, mientras que aquella obtenida para la población de la costa fue de 5,0 µg/mL. Estos y los anteriores resultados muestran la alta variabilidad de nuestras poblaciones de la plaga. Ali et al. (2006) estudiaron en los Estados Unidos la susceptibilidad de H. virescens de poblaciones de diversa procedencia, utilizando la misma metodología. Para una población de algodón obtuvieron una CL50 de 1,20 µg/mL (IC 95%: 0,83-1,78), relativamente menor a la obtenida en los bioensayos con la población de El Espinal durante dos semestres seguidos. Esto comprueba lo ya mencionado por muchos autores que la susceptibilidad de las diversas poblaciones geográficas a la toxina del Bt varía enormemente y depende de los más diversos factores, entre bióticos y abióticos.
Las concentraciones calculadas 4,625 ppm y 4,422 ppm de la toxina en las hojas cotiledonares y en las hojas terminales respectivamente, donde el bellotero acostumbra colocar sus huevos y, se alimentan las larvas neonatas, explica la mortalidad obtenida. Esto comprueba para el control de esta población de H. virescens la eficacia del material transgénico, pero también la necesidad de manejar la posible futura resistencia al Cry1Ac con la siembra de algodón no transgénico en refugios de áreas adecuados y a distancias apropiadas. Sin embargo, a medida que avanza el desarrollo de las plantas, como ya se mencionó, disminuye la concentración de la toxina de una manera marcada. Las concentraciones obtenidas (1,675 ppm en botones florales y 1,82 en cápsulas inmaduras) podrían todavía causar mortalidad a las larvas de primer instar, teniendo en cuenta las concentraciones enmarcadas dentro de los intervalos de confianza de las concentraciones letales encontradas en los dos bioensayos.
Como una ventaja de la población de H. virescens en el Tolima, para atrasar la ocurrencia de resistencia y prolongar la vida de la variedad transgénica, se considera la existencia de huéspedes alternos, la escobita girasol (Lagaxea mollis) y el pega pega, Desmodium sp., en ausencia del algodón. Durante el segundo semestre del 2005, se recolectó precisamente en esta última planta un alto número de larvas de todos los instares de la plaga, para repetir los bioensayos realizados durante el primer semestre. El 99% de las larvas resultó afectado por el parasitoide Cardiochiles nigriceps (Viereck, 1912) (Hymenoptera: Braconidae), impidiendo la realización de los trabajos. La presencia del parasitoide actuando sobre las larvas que se desarrollan en la arvense, se debe tener muy en cuenta para futuros estudios para evaluar además el efecto de la toxina sobre este insecto benéfico.
Los insectos procedentes de Villanueva (Casanare), donde todavía no se ha sembrado algodón Bt, utilizados en otro bioensayo con material vegetal fresco, hojas terminales de algodón transgénico procedentes del Tolima, dio también como resultado una mortalidad del 100% al día siete de haberse iniciado el experimento, mientras que en el testigo no transgénico las larvas se desarrollaron normalmente. Esto podría también significar una alta susceptibilidad al Cry1Ac de la población de H. virescens de esa región.
A pesar de los resultados positivos obtenidos hasta el momento con el algodonero transgénico Bollgard, no se debe descontinuar el monitoreo de la resistencia de H. virescens y, ante todo insistir en las siembras de refugios y la no destrucción de las arvenses mencionadas, escobita girasol y pega pega, sino más bien su conservación y hasta la siembra en los bordes de los lotes. Se debe recalcar que las poblaciones del bellotero durante la última década y específicamente los últimos tres años, incluyendo el primer semestre del año 2007, fueron muy bajas en el Tolima. La situación podría cambiar en cualquier momento y, si hay explosiones demográficas de la plaga como ocurre periódicamente en esta zona algodonera, ésto podría traer consecuencias negativas, inclusive al sembrar fuera del Bollgard I, el Bollgard II, el WideStrike o el Vip3A, sino se maneja adecuadamente la prevención de la resistencia.
Gusano de la mazorca, Helicoverpa zea
Antes de ser liberadas las variedades transgénicas que poseen el gene CrylAc Lutrell et al. (1999), durante los años 1992 y 1993 estudiaron los Heliotinos de diversas regiones geográficas, encontrándose una variable susceptibilidad, la cual fue más acentuada para H. zea que para H. virescens. Aproximadamente diez años más tarde Ali et al. (2006) sugirieron una CL50 mayor en comparación con aquella reportada por Lutrell et al. (1999), inclusive descubrieron, entre todas las poblaciones evaluadas recientemente, una variación de 130-veces entre las concentraciones letales. De acuerdo con Gore et al. (2003a), a pesar de la susceptibilidad del insecto al Cry1Ac, el algodón Bt (primera generación), no ha proporcionado controles adecuados bajo ciertas situaciones. Jackson et al. (2004) confirman estos hallazgos mediante estudios en condiciones de laboratorio y campo. Se considera que, fuera de la menor susceptibilidad de H. zea al Cry1Ac, controles no satisfactorios de la plaga son el resultado de la variación temporal y espacial en la expresión del Cry1Ac entre las partes de la planta (Adamczyk et al. 2001).
La mortalidad de la población autóctona del insecto procedente del Piedemonte llanero colombiano se evaluó mediante tres bioensayos. La CL50 calculado a los siete días de haberse iniciado los bioensayos con las larvas neonatas varió de 3,45 a 6,12 y la CL80 de 7,12 a 10,19 µg/mL (Tabla 2). Los intervalos de confianza muestran que la CL50 corresponde a un valor cercano a 5 µg/mL, lo cual es considerablemente menor que la CL50, 10,06 µg/mL, obtenida para el promedio de todas las muestras de poblaciones de H. zea recolectadas durante dos años de maíz en Estados Unidos (Ali et al. 2006). Si se comparan las concentraciones letales medias con la concentración del Cry1Ac en el cultivar algodonero de Bollgard®, el NuCOTN 33B de 8,56 ± 0,704 (ppm ± D.E) obtenidos por Adamczyk y Gore (2004), vemos la posibilidad de que algunos de los individuos de nuestra población de H. zea escapen a la acción de la toxina, si en la Orinoquía colombiana, específicamente en la zona algodonera de Villanueva, Casanare, se sembrara el algodón transgénico que tiene incoporado la toxina Cry1Ac.
Mortalidad de Helicoverpa zea en los tres bioensayos realizados con dieta merídica con dosis, µg/mL, seriadas del Cry1Ac incorporado (análisis Probit). (Modificado de Zenner de Polanía et al. 2008).
IC = Intervalo de confianza, Rβ = Coeficiente de determinación.
Analizando estos datos, Zenner de Polanía et al. (2008), conceptúan que la variación entre las concentraciones letales entre los tres bioensayos indica una población genéticamente variable del gusano de la mazorca. El amplio rango de susceptibilidad al Cry1Ac podría tener como consecuencia la supervivencia y rápido desarrollo de tolerancia del insecto al Cry1Ac. Un aumento de los dos Heliotinos a poblaciones económicamente relevantes, podría poner en peligro el beneficio actual del Bollgard y requerir aplicaciones de insecticidas para prevenir pérdidas, como lo sugieren también Gore et al. (2003b). Igualmente, Jackson et al. (2003) mencionan una relativamente baja y variable susceptibilidad del bellotero al Cry1Ac en comparación con H. virescens, mientras que Stone y Sims (1993) determinaron una variación de valores de CL50 de cuatro a 60 veces mayor que las estimadas para H. virescens. Varios autores evaluaron el beneficio y recomiendan para un aumento significativo del control del gusano de la mazorca, la adición de la segunda proteina, el Cry2Ab (Gore et al. 2003a; Jackson et al. 2003). Vale la pena también destacar los resultados obtenidos por Chitkowski et al. (2003) quienes determinaron la existencia de poblaciones mucho más bajas de H. zea en Bollgard II que en Bollgard. En nuestro caso, los valores de las concentraciones letales son, sin embargo, solamente un poco mayor para el gusano de la mazorca que para el bellotero, lo cual se podría deber a la ausencia del algodón Bt en el área de procedencia del H. zea. Sin embargo, si en el futuro la diferencia se acentúa, se considera que teniendo en cuenta la dificultad que existe para diferenciar a nivel de campo las larvas de las dos especies, la mayor susceptibilidad de H. virescens podría servir para distinguirlas entre sí.
La concentración de la toxina tuvo una marcada influencia sobre el desarrollo larval y pupal del insecto, expresada en la disminución del peso y el aumento en la duración del estado a medida que aumentaba la dosis. Concentraciones cercanas a o mayores que 1,0 ppm ocasionaron alta mortalidad larval y un elevado porcentaje de pupas deformes. Sin embargo, entre los pesos logrados con dosis hasta 0,1 µg/mL y el testigo absoluto no hubo diferencia estadísticamente significativa (Tabla 3) (Zenner de Polanía et al. 2008).
Peso promedio de las pupas sobrevivientes de Helicoverpa zea de los bioensayos I y II (Modificado de Zenner de Polanía et al. 2008).
Promedios, dentro de la misma columna, seguidos por la misma letra no son significativamente diferentes de acuerdo a la prueba de Tukey (a = 0,05).
Al comparar el peso promedio de las pupas del testigo de la raza colombiana, 402,37 ± 28,10 (Tabla 3), con los pesos, 451,82 ± 3,17 y 418,83 ± 5,27, encontrados por Giolo et al. (2006) al utilizar dos dietas merídicas, se observa un peso significativamente bajo en el estudio actual y además una DMS mucho mayor. Las diferencias se podrían atribuir a deficiencias nutricionales en la dieta, a la variedad del gusano de la mazorca y a las condiciones ambientales de los laboratorios. Con una dieta, consistente en pedazos de mazorca tierna el peso de las pupas aumentó a 422 µg ± 0,0639. Se advierte la influencia de la calidad del alimento, de la variabilidad genética y de la procedencia sobre el peso de las pupas de H. zea, lo cual podría implicar una variación en la supervivencia del insecto en el algodón transgénico. Concentraciones mayores que 0,1 ppm en el tejido vegetal, atacado por H. zea, aunque no causaría un 100% de mortalidad, en teoría, sí disminuiría considerablemente la emergencia de adultos viables. Teniendo en cuenta la concentración máxima obtenida en botones florales y cápsulas inmaduras, partes del algodonero afectadas preferencialmente por la plaga (Tabla 1), la posibilidad de una marcada supervivencia de la plaga al algodonero transgénico que se siembra en el país es poco probable.
Cogollero del maíz, Spodoptera frugiperda y gusano rasputín, S. sunia
Si con las plagas Heliotinas, se considera el algodonero transgénico justificado desde el punto de vista de su costo y adopción, con Spodoptera frugiperda se observa todo lo contrario. Sus poblaciones se han incrementado a niveles y daños tan altos desde que se inició la siembra de estos cultivares que, ya algunos técnicos y agricultores manifiestan que sus siembras no son competitivas.
El complejo Spodoptera es considerado en nuestro medio un grupo de especies insectiles de mayor importancia económica en el algodonero, mientras que en Estados Unidos, donde se crearon y evaluaron los algodones transgénicos no tiene importancia en este cultivo y no fue por lo tanto considerado como plaga objetivo. A pesar de que varios autores mencionaron que el Cry1Ac no ejerce controles satisfactorios del S. frugiperda (Garczynski et al.1991; Bohorova et al. 1997; Ayra-Pardo et al. 2006), las indicaciones iniciales de los productores de semilla, sí incluyeron a la plaga dentro de las controladas por la variedad transgénica sembrada en el país.
Los resultados de los primeros tres bioensayos realizados durante el año 2005, comprueban lo expresado por los autores mencionados; así mismo, Zenner de Polanía et al. (2005) concluyeron que: 1) Después de solo una generación de alimentación con tejido vegetal de algodonero transgénico, el ciclo de vida de la plaga se alarga, lo cual podría disminuir el número de generaciones por año, 2) la concentración de la A-endotoxina Cry1Ac existente en cotiledones, hojas terminales y botones florales del algodonero utilizado como alimento para las larvas, no causa mortalidad alguna al insecto, pero si influye negativamente en el peso de las pupas, 3) El menor peso de las pupas, tanto de machos como de hembras, no afecta la fertilidad y fecundidad de la plaga.
Posteriormente, trabajando con larvas neonatas del cogollero, procedentes de posturas recolectadas en hojas de algodón transgénico de El Espinal (Tolima), y alimentándolas con hojas cotiledonales, se observó un desarrollo inicial normal durante el primer instar, el cual tuvo una duración promedia de 2,5 ± 0,5 días. Se alimentaron sobre el envés de las hojas cotiledonales, ocasionando las ventanitas características del daño inicial de la plaga. La duración del II instar fue muy homogénea, la totalidad de las larvas mudaron al cabo de tres días. Durante el III instar se comenzó a observar un comportamiento anormal, aunque, exceptuando dos larvas, todas lograron entrar al cuarto instar, al cabo de cuatro días en promedio. Luego, sin embargo, dejaron de alimentarse, se encogían y construían un tipo de celda, como para iniciar la transformación en pupa. Inicialmente, se pensaba en un posible efecto de la toxina del Bt, pero luego se constató que estaban parasitadas por el parasitoide huevo-larva Chelonus insularis (Cresson, 1865) (Hymenoptera: Braconidae). El porcentaje de parasitismo fue del 90,47% y en el testigo (alimentado con dieta merídica) del 100%.
Los primeros adultos del parasitoide emergieron a los 27 días del inicio del experimento; este dato coincide con aquel obtenido por Medina et al. (1988), quienes encontraron que el ciclo del C. insularis corresponde a 29,05 días de huevo a la emergencia del adulto. Un 83,60% de los parasitoides alcanzó el estado adulto, lo cual indica que la alimentación de las larvas parasitadas de S. frugiperda con tejido fresco de algodón transgénico, cotiledones y hojas terminales, no tuvo efecto negativo sobre el desarrollo normal del parasitoide, por lo menos hasta el estado adulto.
C. insularis es uno de los parasitoides más eficientes del cogollero del maíz y se encuentra distribuido desde el norte de la Argentina hasta el sur de Estados Unidos (Ashley 1983; Cruz et al. 1991; Fernández y Clavijo 1984). La ausencia de un efecto negativo de la toxina del Bt incorporada al algodonero, del cual se alimenta su huésped el cogollero del maíz, sobre el desarrollo del parasitoide es positivo, pero se debe primordialmente a la dosis subletal del Cry1Ac en el follaje, la cual no ocasiona la muerte a la plaga. Si la mortalidad de la plaga hubiese sida alta, lo mismo hubiera ocurrido con el insecto benéfico.
La posibilidad de recolectar en el campo posturas del cogollero parasitadas por C. insularis es relativamente alta, ante todo durante la segunda generación del cogollero (Zenner de Polanía et al. 2006). Por esta razón no se deben emplear las larvas procedentes de estas posturas para la realización de bioensayos, sino criarlas hasta la F2 y así asegurar la ausencia del parasitismo.
Las posturas del cogollero, empleadas para el tercer ensayo fueron recolectadas en un lote de algodón transgénico en El Espinal, criadas hasta la F2 y las larvas neonatas que emergieron empleadas directamente. Simultáneamente, se realizó la misma evaluación para el gusano Rasputin, S. sunia, plaga importante en el Tolima y a veces confundido en el estado larval con el cogollero del maíz.
La respuesta dosis mortalidad calculada por Probit, a través de 20 días del desarrollo larval de las dos especies de Spodoptera, se consigna en la Tabla 4 y, en la Figura 1, se ilustra el porcentaje de mortalidad observado durante este lapso. Los resultados muestran que el cogollero es un poco más susceptible a la toxina Cry1Ac que el gusano Rasputín, pero, ninguna de las dos especies puede ser controlada con el cultivar transgénico. La susceptibilidad de ambas especies aumenta lentamente a través del tiempo de exposición, lo cual es confirmado por la pendiente poco pronunciada de la línea respuesta dosis-mortalidad.
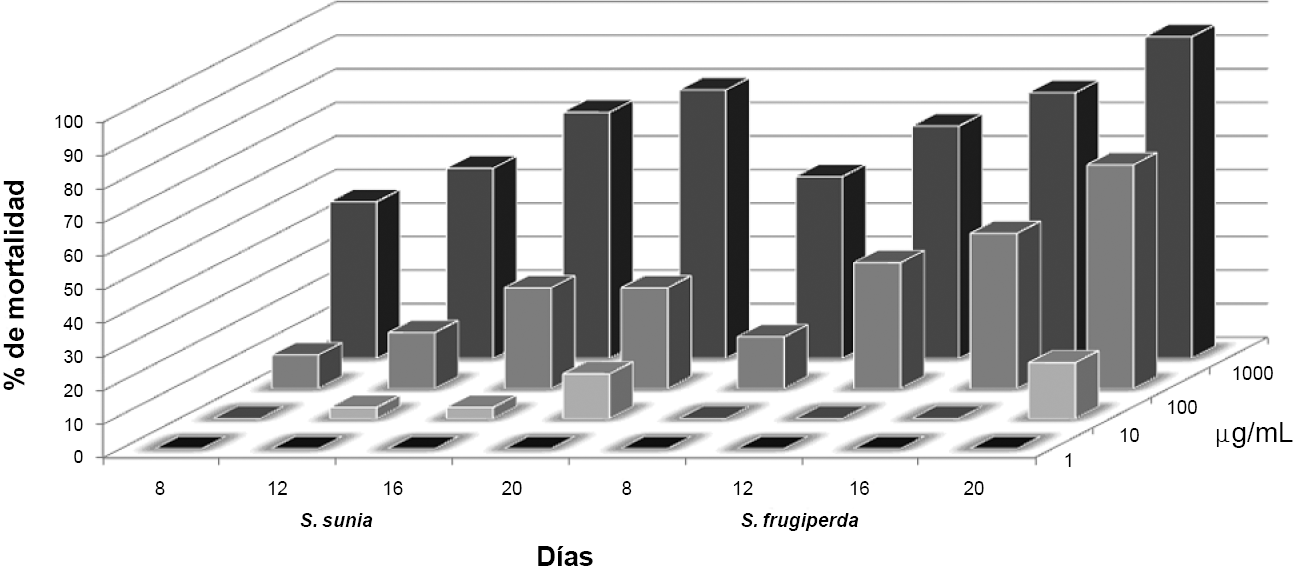
Porcentaje de mortalidad de Spodoptera sunia y S. frugiperda a través del tiempo de exposición a dosis seriadas del Cry1Ac incorporado en dieta merídica.
Concentraciones letales, µg/mL, de Spodoptera frugiperda y S. sunia alimentados con dieta merídica con dosis seriadas de incorporadas (Probit) (semestre 2005A).
IC = Intervalo de Confianza, Rβ = Coeficiente de determinación.
La Figura 1 muestra un porcentaje de mortalidad algo mayor de las larvas de S. frugiperda que de S. sunia a través del tiempo. A los 20 días, cuando las larvas del testigo de ambas especies ya estaban llegando al VI instar, y aquellas en la dieta con la dosis de 1,0 ppm al V instar, las que se desarrollaban con 1.000 ppm apenas llegaban al II instar. La dieta con 10 ppm de la toxina tuvo un efecto algo menor sobre el cogollero que sobre el gusano Rasputin durante los primeros 16 días y solo ocasionó una mortalidad del 16,66% y 13,33% a los 20 días, respectivamente. Esto, bajo condiciones de campo significa la ausencia de un control de las dos especies con el algodón transgénico sembrado actualmente en el país
En la Tabla 5 se comparan las concentraciones letales calculadas de poblaciones del S. frugiperda procedentes del Meta y del Tolima y de diferentes huéspedes y en diferentes semestres. Analizando estos datos resalta inicialmente la similitud de las concentraciones letales (CL) medias obtenidas para las razas procedentes del Meta y del Tolima, obtenidas de maíz y algodón respectivamente. Este resultado confirma preliminarmente, para Colombia, lo observado por Martinelli et al. (2006) en el Brasil que las poblaciones de S. frugiperda que se desarrollan en maíz y en el algodonero corresponden a la variante de maíz (Pashley et al. 1985), mientras que posibles diferencias genéticas entre estas poblaciones se deben más bien a variantes que se desarrollaron en diferentes regiones geográficas (Zenner de Polanía et al. 2007). En este contexto sería de gran interés desde el punto de vista manejo, comparar genéticamente poblaciones del cogollero procedentes de los Valles Interandinos colombianos, de la Costa Atlántica y de la Orinoquía recolectadas en las diversas especies huéspedes de la plaga.
Respuesta dosis mortalidad para Spodoptera frugiperda en bioensayos realizados con dieta merídica con dosis, ì g/mL, seriadas del Cry1Ac incorporado (análisis Probit).
IC = Intervalo de confianza, rβ = Coeficiente de determinación.
La disminución considerable de la CL observada en larvas del cogollero procedentes de soca de maíz se atribuye a la dilución de la resistencia al Cry1Ac, debido al número de generaciones del insecto transcurrido en este cultivo y probablemente otro en la soca, lo cual podría sumar un total de cuatro generaciones sin presión de selección. La población se desarrolló en el semestre con veda para la siembra del algodonero, así que no hubo migraciones entre cultivos de maíz y algodonero transgénico, ni posibilidad de intercambio de genes que imparten resistencia. Esta ausencia de presión de selección en el segundo semestre del año en el departamento del Tolima, se debería aprovechar con siembras de maíz convencional, para lograr una mayor susceptibilidad del insecto al Cry1Ac y así lograr un manejo más adecuado de la plaga. Sin embargo, resalta la pronunciada pendiente de la línea de regresión (3,43), la cual indica que a medida que el insecto esté nuevamente sujeto a presión de selección, la tolerancia aumentará mucho más rápido que en ocasiones anteriores. Desde el punto de vista manejo del cogollero del maíz en el algodonero transgénico, los datos aquí presentados confirman lo observado por Zenner de Polanía et al. (2005) que la toxina Cry1Ac en las concentraciones disponibles en la planta no ejercen un control de S. frugiperda.
Recomendaciones de manejo del algodonero transgénico
Fuera de las exigencias de dejar refugios apropiados de algodón convencional para que las plagas controladas por la variedad transgénica se puedan desarrollar sin la presión de selección que, a la larga ocasiona el fenómeno de resistencia y hace que la tecnología pierda su efecto, se debe tener en cuenta en nuestro medio: 1) en los cultivos, huéspedes alternos, no utilizar productos con base en B. thuringiensis para el control de los insectos plagas del Orden Lepidoptera, 2) permitir, en el semestre no algodonero refugios adicionales de sus huéspedes alternos, a pequeña escala, para el bellotero, H. virescens. 3) usar controles biológicos para el manejo del complejo Spodoptera en el maíz, el sorgo y el algodonero transgénico y convencional.
Footnotes
Agradecimientos
A COLCIENCIAS y a la Universidad de Ciencias Aplicadas y Ambientales U.D.C.A por la financiación del proyecto. Al Ingeniero Agrónomo Guillermo Álvarez Viecco, a Remolino S.A. y a FENALCE por su colaboración logística y técnica.