
Abstract
Resumen
Buscando entender los diferentes patrones de variación estacional de la estructura trófica del ensamblaje de coleópteros, se realizó un trabajo en la Amazonia Colombiana, durante tres años consecutivos en época de lluvia y de sequía, en bosque de tierra firme y várzea. Se realizaron transectos donde se instalaron trampas: de caída, Corner, Winkler, Malaise y adicionalmente se realizaron capturas manuales y zarandeo. El material se identificó a nivel de familia. Se capturaron 3.691 individuos, correspondientes a 32 familias. Las familias más abundantes en número de individuos fueron Scolytidae (31,7%), Scarabaeidae (222%) y Staphylinidae (18,1%). Los xilomicetófagos (32,1%) fueron el grupo dominante, seguido por los copro-necrófagos (22,3%), depredadores (21,7%) y herbívoros (11%). Al comparar entre años y entre hábitats se encontraron diferencias en la abundancia y composición de las familias, así como en el aporte de los roles tróficos. La época seca demuestra una tendencia al empobrecimiento. La marcada variación espacial y temporal entre estaciones y entre años, se asocia a la alta diversidad y estaría apoyando la hipótesis que el recambio estaría jugando un importante papel en la dilución de la competencia interespecífica por el recurso.
Introducción
El orden Coleoptera presenta más de 400 mil especies descritas, las cuales se encuentran contenidas en cerca de 166 familias (Lawrence y Newton 1995), encontrándose en todos los ecosistemas y representando todos los roles tróficos (Crowson 1981). La mayoría de los coleópteros son considerados fitófagos, presentándose algunos roles poco comunes y muy especializados como los micetófagos o los queratinófagos (Arnett y Thomas 2000). A raíz de esta estrecha relación con algunos procesos al interior de los ecosistemas, muchas familias de coleópteros han sido utilizadas como bioindicadores (Brown 1991; Pearson 1994; Halffter y Favila 1993), demostrando su importancia en planes de conservación como herramientas de monitoreo (Brown 1991; Kremen et al. 1993).
Trabajos realizados por Erwin (1982, 1983) en bosques neotropicales, proponen la existencia de una alta riqueza, que puede alcanzar el valor de ocho millones de especies de coleópteros. Adicionalmente a este valor hay que destacar la fuerte dinámica temporal y espacial, la cual en ciertas localidades puede ser marcada causando un importante incremento en la riqueza local (Erwin y Scott 1981). El conocimiento de esta dinámica estacional en un determinado ecosistema puede ofrecer información importante para la generación de estrategias y planes de conservación enfocados a mantener la funcionalidad del mismo (Pimm 2002; Tilman y Downing 1994; Naeem et al. 1995; Johnson et al. 1996).
En este sentido son escasos los trabajos que han intentado acercarse al estudio de la estructura temporal del ensamblaje de coleópteros (Elton 1973; Krasnov y Ayal 1995; Zerm y Joachim 2001; Crouau-Roy et al. 1992; Blom et al. 2002), especialmente al análisis del efecto que los roles tróficos tienen en el funcionamiento de un ecosistema (Mikkelson 1993; Lawton 1995; Escobar y Chacón de Ulloa 2000; Paarmann et al. 2001; Arias-Díaz et al. 2007). Algunos trabajos muy específicos en la amazonía, se han acercado al estudio de ciertas variables pero en grupos delimitados (Klein 1989; Pearson y Cassola 1992). El único trabajo que hasta la actualidad ha estudiado la estructura trófica del ensamblaje de coleópteros en la región amazónica (Brasil - Amazonia Central) es la investigación de Didham et al. (1998).
Desde mediados del siglo pasado diferentes investigadores han mostrado un profundo interés en estudiar la variación estacional en insectos (Davis 1945; Dobzhansky y Pavan 1950), a pesar de ello, la mayoría de los trabajos en el caso especifico de los coleópteros, solo han abarcado algunas pocas familias o se han centrado en aspectos comportamentales puntuales (New 1979; Krasnov y Shenbrot 1997; Yahiro y Yano 1997; Gök y Durán 2004; Boyer et al. 2003). Igualmente son pocos los trabajos realizados en la región tropical: Africa (Paarmann 1979), las Antillas (Gruner 1975), México (Montes de Oca y Halffter 1995), Costa Rica (Janzen 1983), Panamá (Wolda y Estribi 1985; Chandler y Wolda 1986), Ecuador (Peck y Forsyth 1982) y Brasil (Pinheiro et al. 2002).
Para el caso de Colombia y especialmente en la región amazónica, son pocos los estudios realizados acerca de la dinámica de insectos y menos en el orden Coleoptera. Vale la pena resaltar el trabajo efectuado por Howden y Nealis (1975) en Leticia (Amazonas - Colombia), los cuales trabajaron únicamente con la familia Scarabaeidae comparando los cambios entre dos años de muestreo y el trabajo de Martín Piera y Fernández (1996), quienes estudiaron la variación estacional de coleópteros en la región de Chiribiquete (Caquetá- Colombia).
Todo esto demuestra la escasa información acumulada, tanto de la composición y estructura, como de la dinámica espacial y temporal de esta provincia biogeográfica en el país, más aun si se resalta que en ninguno de los trabajos mencionados se ha incluido un análisis de la dinámica a nivel de los roles tróficos, por todo lo anterior este estudio se constituye en el primer trabajo de este tipo para la amazonia colombiana, donde se presenta por primera vez el acumulado de tres años de muestreo consecutivo.
Materiales y Métodos
Área de estudio
El trabajo se llevó a cabo en la Amazonía Colombiana, en la Comunidad Monifue Amena (Resguardo Indígena Ticuna-Uitoto), a 71 msnm (4°06′46,25″S - 69°55′52,11″W), a 9,5 km de la ciudad de Leticia, Departamento del Amazonas - Colombia (Fig. 1). En esta zona se presenta un ciclo pluvial monomodal, con una época de sequía de junio a octubre. El muestreo se llevó a cabo en dos unidades de paisaje: Bosque de Tierra Firme (BTF) y Bosque Inundable o Várzea, en época de lluvias (marzo-mayo) y en época seca (septiembre-octubre), durante tres años consecutivos: 2002, 2003 у 2004.
Localización de la zona de estudio, comunidad Monifue Amena, Leticia Colombia.
Muestreo
Los muestreos se realizaron con diferentes métodos de captura: trampas Pitfall, trampas Winkler, trampas Malaise, recolección manual y zarandeo; usando 924 trampas Pitfall, 12 Malaise, 24 Winkler, para un total de 576 horas efectivas (Fig. 2).
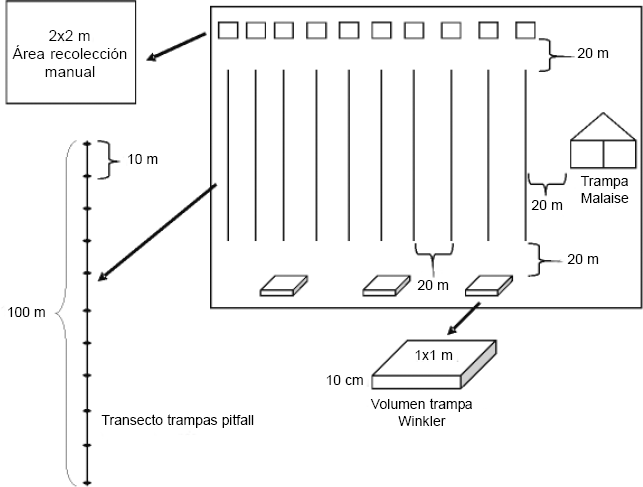
Disposición de los transectos, parcelas y trampas colocadas en cada unidad de muestreo, por cada época en cada año, en la comunidad Monifue Amena, Leticia - Colombia.
Cada uno de los muestreos tuvo un periodo de 48 horas por trampa, para los dos hábitats. En cada época se realizó un muestreo consistente en la instalación de un promedio de 10 transectos (± 1, el número de transectos varió debido al número de personas que asistían a cada uno de los muestreos) de 100 m, separados 20 m, en cada uno de los dos tipos de bosques. Sobre el transecto se colocaron 11 trampas tipo pitfall de 9 onzas, separadas 10 m entre sí, cebadas con salchichón en descomposición y con alcohol como liquido preservante. Para cada unidad, adicionalmente se seleccionó un área de 2 x 2 m, la cual fue muestreada por tres personas durante una hora con jamás y pinzas. Los muestreos y la colocación de las trampas se hicieron en horas de la mañana en BTF y en horas de la tarde en varzea.
Cerca de cada transecto, se seleccionaron cuatro árboles con un diámetro a la altura del pecho (DAP) de 8 cm, en los cuales se efectuaron tres episodios seguidos de zarandeo sobre una sabana blanca, en donde se recolectó todo lo que caía. Al final de tres transectos alternados a una distancia de 20 m se tomó un mβ de tierra para pasarlo por un saco Winkler, en cada uno de los bosques. Finalmente en cada unidad de paisaje, a un mínimo de 50 m de los transectos, se colocó una trampa Malaise de 200 x 100 x 50 cm, rasante al piso, que permaneció 48 horas activa.
Identificación taxonómica
Los ejemplares recolectados se mantuvieron en etanol al 70%, posteriormente se realizó un montaje en seco, con alfileres entomológicos, montando un ejemplar por cada morfotipo encontrado. Con el material montado se realizó la identificación a nivel de familia, utilizando claves generales y especializadas en este grupo (Costa Lima 1952; White 1983; Papp 1984; Bland y Jaques 1978; Arnett y Thomas 2000; Borror et al. 1989; Arnett et al. 1980; Arnett et al. 2002). El material finalmente fue etiquetado para ser depositado en la Colección Entomológica del Museo de la Pontificia Universidad Javeriana. Es importante anotar que en el muestreo de la época de lluvias del 2003, se presento una pérdida de material, ocasionada por un mal transporte del mismo.
Asignación de los grupos tróficos
Para la caracterización de los roles tróficos de cada una de las familias, se realizó una revisión de la información de historia de vida y tendencias alimenticias a partir de diferentes trabajos (Crowson 1968, 1981; Paulian 1988; Lawrence y Newton 1995; Arnett y Thomas 2000; Arnett et al. 2002). A raíz de encontrar discrepancias entre los diferentes autores, especialmente relacionadas con variaciones tróficas al interior de algunas familias, se llegó a un consenso, tomando la tendencia general.
Análisis estadístico
Se realizó una prueba de ANOVA, para determinar la existencia de diferencias significativas entre los años, entre las épocas de muestreo, entre los hábitats y la combinación entre ellos, por medio del programa STATISTICA v. 6. Adicionalmente a esto, se calculó el Índice de Similitud entre años, épocas y unidades de paisaje, utilizando el índice de Sorensen como la medida de distancia para definir la similitud y como método de unión de grupos el promedio entre grupos (UPGMA). Para la ordenación de los muestreos se utilizo un Análisis de Correspondencia Rectificado (DCA), para evidenciar gráficamente la existencia de agrupamientos, por medio del programa PC-ORD v. 4.
Resultados
Abundancia y riqueza
Se capturaron un total de 3.691 individuos, distribuidos en 32 familias (Anexo 1), de las cuales las más abundantes fueron Scolytidae (31,7%), Scarabaeidae (22,2%) y Staphylinidae (18,1%). Se encontraron familias con hábitos generalistas temporales, que fueron capturados durante todos los años independientemente de la época de muestreo, tales como Scolytidae, Staphylinidae, Scarabaeidae, Carabidae y Nitidulidae; especialistas espaciales que solo se encontraron en BTF como Cicindelidae o Noteridae y familias raras de las cuales solo se capturaron uno o dos individuos como Buprestidae, Coccinelidae, Dermestidae, Meloidae, Oedemeridae y Anobiidae, en un solo periodo y un hábitat específico.
Familias de Coleoptera capturadas durante los tres años de muestreo en la comunidad Monifue Amena, Amazonas - Colombia (taxonomía según Arnett et al. 2002).
El año que mayor abundancia presentó fue el 2003 con un total de 1366 individuos (37,01%), seguido del 2002 (1247, 33,78%) y el 2004 (1078, 29,21%). Con respecto a la riqueza, no se presentaron grandes diferencias en cuanto al número de familias; el promedio de familias fue de 24 para los tres años, siendo el 2002 el menor año con 23 familias y el 2004 el mayor con 25 familias. En cuanto a la abundancia se encontraron diferencias significativas entre los tres años (F = 0,987; P < 0,009).
Hubo mayor abundancia en la época de lluvias (2085, 56,49%), que en la época de sequía (1606, 43,51%), a excepción de lo encontrado en el año 2003 donde esta relación se invirtió (Fig. 3). Con respecto al número de familias, se presentó una tendencia a aumentar en época seca (Fig. 3) y en el hábitat BTF (Fig. 4). El porcentaje total de individuos fue mayor en BTF (2423, 65,9%) que en Várzea (1259, 34,1%), mostrando una tendencia a mantener la proporción a lo largo de los diferentes muestreos (Fig. 3). Se encontraron diferencias significativas entre las abundancias y las riquezas de los dos hábitats (F = 0,984; P < 0,003), así como entre las épocas al interior de los años (F = 0,987; P < 0,001).
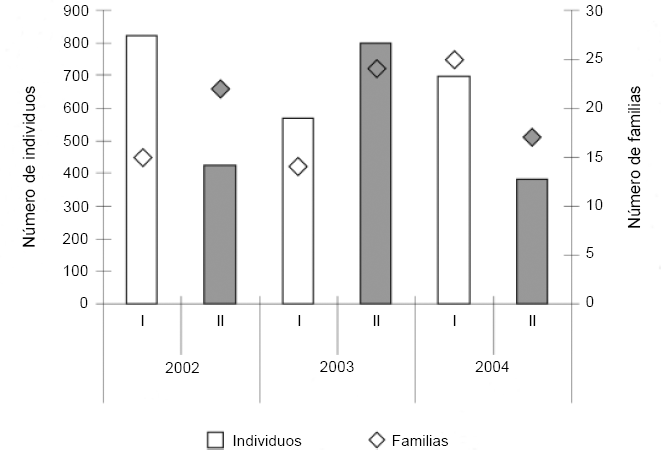
Número de individuos (barras) y de familias (rombos) en cada una de las dos estaciones (lluvias: I y seca: II), en los tres años de muestreo, en la comunidad Monifue Amena, Leticia - Colombia.
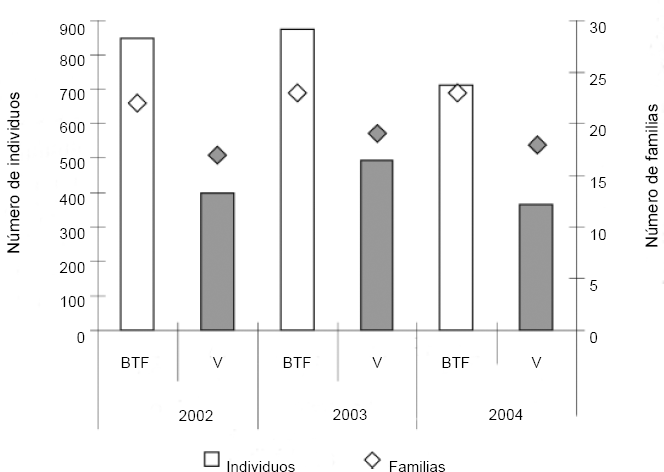
Número de individuos (barras) y de familias (rombos) en cada una de las unidades de paisaje (BTF y Várzea), en los tres años de muestreo, en la comunidad Monifue Amena, Leticia - Colombia.
No se encontraron diferencias significativas entre las épocas de mayor o menor precipitación; entre los hábitats al interior de cada año y entre los hábitats y las épocas. El análisis de similitud muestra una relación (> 85%) entre las épocas de lluvias de los años 2003 y 2004, independientemente de la unidad de paisaje y también una relación (90%) entre las unidades de paisaje durante la época de lluvias del año 2002. El análisis de correspondencia tiende a agrupar los muestreos por años y dentro de estos por unidad de paisaje (Fig. 5).
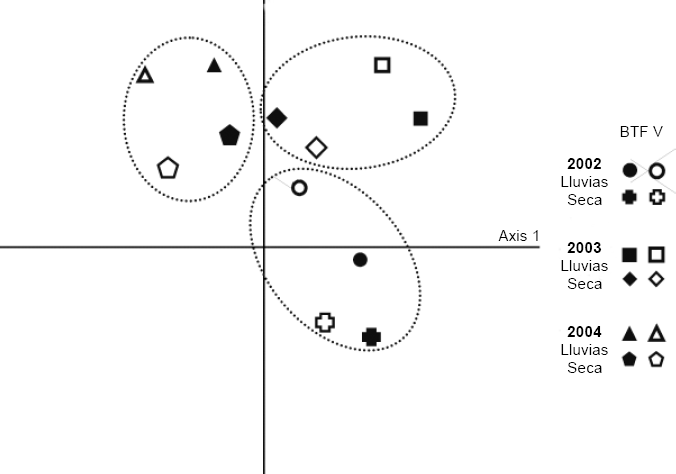
Análisis de correspondencia entre los años, las estaciones pluviométricos y los hábitats muestreados en la comunidad Monifue Amena, Leticia - Colombia.
Estructura trófica
Se establecieron ocho roles tróficos a partir de las familias recolectadas, dentro de los cuales el que presentó mayor abundancia correspondió a los xilomicetófagos (32,1%), seguido por los copro-necrófagos (22,3%) y los depredadores (21,7%), mientras el resto de roles presentaron números bajos de individuos (Fig. 6A). Con respecto a la riqueza de familias se encontraron valores diferentes reflejando una composición no consistente con la abundancia; el rol que mayor número de familias presentó fue el herbívoro con 13 (40,6%), seguido por los depredadores con ocho (25%) y los saprófagos con cuatro (12,5%), mientras el resto de roles presentaron un número bajo de familias (Fig. 6B). No se presentaron familias con hábitos parasitoides.
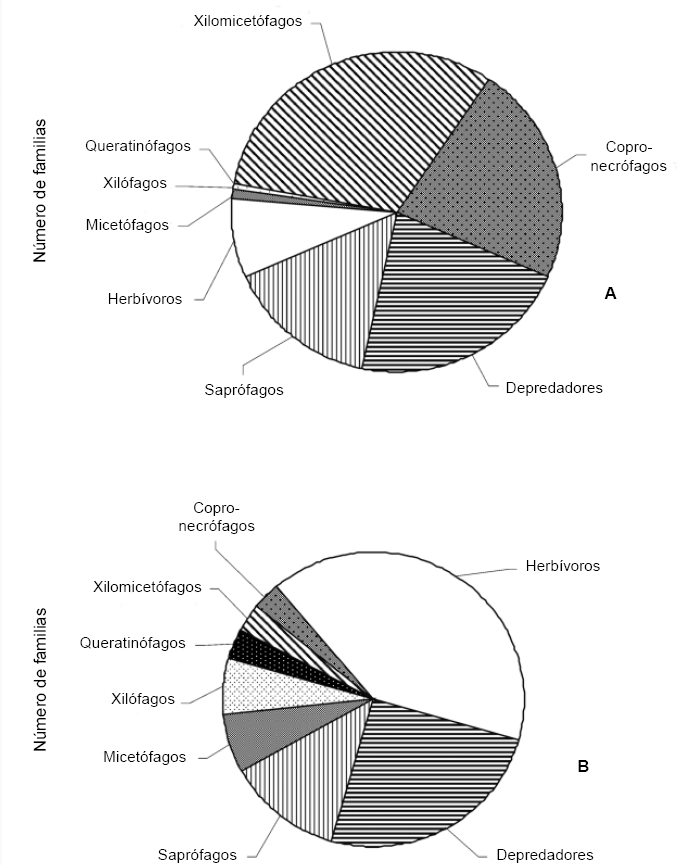
Proporción del número de individuos (n = 3691) (
Estacionalidad
En la variación en la riqueza de familias por unidad de paisaje/año, BTF presenta una mayor riqueza de familias que Várzea (Fig. 4), mientras en el caso de las épocas de mayor precipitación no se presentó un patrón evidente (Fig. 3).
Roles tróficos.
En cuanto a los roles tróficos y la estacionalidad, se observaron algunos cambios importantes entre los muestreos; para algunos roles como los xilomicetófagos y los copro-necrófagos se observó una marcada estacionalidad, mientras que para los demás roles los valores de abundancia fueron relativamente constantes (Fig. 7). Los roles raros como queratinófagos, xilófagos y micetófagos desaparecen para algunos muestreos, especialmente en el 2003 y en la época de lluvias (Fig. 7).
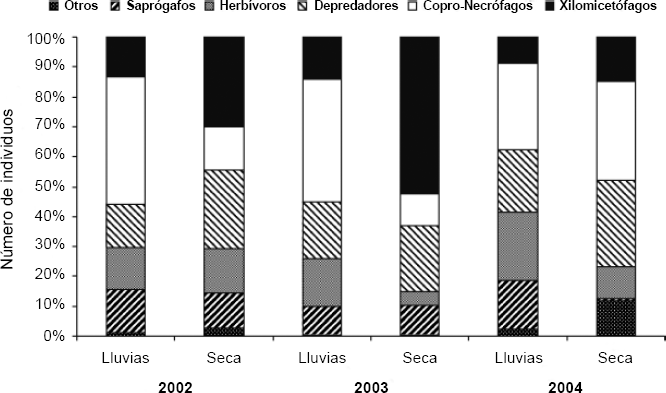
Porcentaje del número de individuos en cada uno de los roles tróficos del ensamblaje, en cada estación por año de muestreo, en la comunidad Monifue Amena, Leticia - Colombia.
Discusión
Los resultados concuerdan con varios estudios que evidencian una marcada estacionalidad de muchos grupos de insectos, especialmente en el caso de los coleópteros (Wolda 1978; Janzen 1983; Wolda 1988; Chandler y Wolda 1986; Montes de Oca y Halffter 1995), pero adicionalmente plantea que la estacionalidad no solo está relacionada con la abundancia y riqueza de los ensamblajes sino con la funcionalidad de los mismos. Al presentarse cambios en la estructura trófica, es de esperar que determinados procesos funcionales se vean afectados de igual manera (Didham et al. 1998; Boyer et al. 2003).
La mayor abundancia de los individuos en la época de lluvias puede estar relacionada con una mayor oferta de recursos a nivel cualitativo y cuantitativo, que genera una mejor disponibilidad espacio-temporal (Huston 1996). De igual forma las condiciones climáticas son más favorables, como se demuestra en algunos estudios a nivel neotropical (Lauer 1992; Losos y CTFS Working Group 2004). Adicionalmente, otro factor que podría influir es la agregación espacial que se produce en época seca de ciertos recursos, lo cual aumentaría la competencia interespecifica (Foster 1996). A todo lo anterior hay que agregar la existencia de recursos específicos que solo aparecen en época de lluvias, presentando una marcada estacionalidad y condicionando a las especies que han establecido relaciones estrechas a mantener estos mismos ciclos (Garwood 1996; Leigh y Windsor 1996).
Adis (1988) reporta una fuerte estacionalidad para los bosques de la amazonía central en Brasil, donde algunas familias de coleópteros aparecen solo en una determinada época del año, similar a lo encontrado en este estudio. Aunque algunos trabajos han arrojado una mayor riqueza y abundancia en época seca (Watanabe y Ruaysoongnerm 1989), es evidente que para el caso del neotrópico y los bosques de la amazonia la mayor abundancia y riqueza están estrechamente asociadas con la llegada de las lluvias (Holt y Spain 1986; Frith y Frith 1990; Krasnov y Shenbrot 1997; Gök y Durán 2004; Boyer et al. 2003).
La baja riqueza de familias en la época seca puede estar obedeciendo a diversos factores como la existencia de grupos especializados a estas condiciones o a recursos específicos de esta época (Losos 2004; Levings y Windsor 1996). También es posible considerar que las familias que aparecen en esta época sean malas competidoras, las cuales estarían optando por una estrategia suboptima en una época más difícil, pero con una menor competencia. Es interesante anotar que las familias presentes en esta época son herbívoras, beneficiándose del estrés hídrico que podría disminuir los mecanismos de defensa de algunas plantas (Waterman y McKey 1992), dándoles una ventaja sobre la calidad del recurso existente, aumentando la diversidad, pero manteniendo poblaciones bajas.
Las diferencias encontradas en el número de familias e individuos entre BTF y Várzea, responden a una mayor complejidad estructural a nivel vegetal presente en BTF (Duivenvoorden 1996), lo cual genera no solo una mayor cantidad y diversidad de recursos, sino microhábitats potenciales, en donde por ejemplo las presas pueden escapar de los depredadores (Prance 1992; Huston 1996; Losos y CTFS Working Group 2004). Es importante mencionar que el área de cobertura de BTF es mucho mayor que el área de la Várzea, lo cual puede llegar a afectar los valores encontrados, especialmente en el caso de las abundancias totales por hábitat.
Algunos trabajos han evidenciado que tanto la composición vegetal del bosque como la composición de los artrópodos puede verse afectada por las inundaciones (Beck 1972; Gasdorf y Goodnight 1963; Uetz et al. 1979), como se reporta para este estudio. A pesar de ello, algunas especies pueden escapar a las inundaciones, por medio de migraciones verticales, sobreviviendo debajo del agua, o coordinando sus ciclos para reproducirse cuando el suelo está seco (Irmler 1979; Paarmann 1982; Adis 1988). También es posible que se presenten migraciones de Várzea a BTF en la época de inundaciones (Uetz et al. 1979).
Mientras menor sea la inundación y entre menos eventos de inundación existan en el año, mayor es la riqueza encontrada, por lo que los valores de riqueza fluctuarían más en zonas con fuertes inundaciones periódicas (Uetz et al. 1979). Algunas especies y familias pueden estar coordinadas con el inicio de la época de lluvias (Morais 1985; Adis 1988), mientras que otras pueden presentar rangos más amplios, especialmente aquellas especies con una alta capacidad de desplazamiento (Uetz et al. 1979), como sucedería en este estudio con la familia Scarabaeidae.
Aunque algunos autores proponen que la presencia de disturbios intermedios periódicos puede aumentar la diversidad de un lugar (Connell y Slatyer 1977), la existencia de periodos de inundación en la Várzea, es sin duda un evento que impide su utilización por parte de los insectos, ya que son pocos los grupos que pueden adaptarse a este régimen, lo cual explicaría porqué la diversidad disminuye drásticamente en esta época (Uetz et al. 1979). Las diferencias entre años podrían atribuirse a variaciones climáticas, tanto por desplazamiento de los periodos de lluvias, como por la intensidad de los mismos (Lauer 1992). De igual manera, aunque los muestreos se realizaron en las mismas áreas, variaciones en la ubicación microespacial del muestreo pueden afectar la abundancia y riqueza de lo capturado (Melbourne 1999).
Otro aspecto que puede estar afectando las variaciones entre los años, es la existencia de ciclos ambientales o biológicos que trascienden la temporalidad anual, presentando intervalos más amplios de los tres años de muestreo. Igualmente es posible que con las fuertes dinámicas existentes en los bosques neotropicales (Losos y CTFS Working Group 2004; Leigh et al. 1996; Vallego et al. 1996), se hayan muestreado ensamblajes diferentes, tanto por la escogencia de microhábitats diferentes, como por eventos aleatorios que ocasionen una mayor heterogeneidad al interior de cada año.
En relación con los roles tróficos, se ha evidenciado una marcada estacionalidad de diferentes recursos (Leigh y Windsor 1996), como es el caso de la hojarasca y la madera, recursos que son más fáciles de degradar en época húmeda por hongos, mientras que en época seca los xilófagos y xilomicetofagos pueden dominar el recurso, generando una fuerte competencia y una dinámica marcada entre estos dos grupos (Bagyaray 1992; Brokaw 1996; Esser y Lieth 1992; Swift y Anderson 1992). En este sentido el rol menos afectado por la precipitación sería el de los depredadores, aunque a nivel general en época seca la mayoría de los roles disminuyen por la falta de una humedad adecuada para llevar a cabo algunas funciones, como en el caso de los coprófagos donde el excremento llega a ser inutilizable (Cambefort y Hanski 1991).
Existe un mayor número de individuos asociados a aquellos recursos que presentan mayor abundancia, por lo que los recursos efímeros en espacio y tiempo, presentan una mayor cantidad de roles pero pocos individuos (Hanski 1990; Hanski y Cambefort 1991). En este sentido podría pensarse que los roles dominantes, que no desaparecen en el tiempo, tienen una mayor importancia para el constante funcionamiento del ecosistema, aunque la desaparición de roles no tan comunes, podría afectar los procesos en una escala temporal a largo plazo.
Aquellos roles tróficos raros o asociados a recursos poco abundantes, tendrían una mayor posibilidad de presentar extinciones locales al ser más susceptibles (Larsen et al. 2005), por lo que deberían ser grupos bandera en estrategias de conservación. Algo similar estaría sucediendo con niveles tróficos superiores, los cuales presentan una mayor susceptibilidad a la extinción que grupos basales (Holt 1996). En este sentido debe plantearse el uso de especies funcionales, que aporten información más completa sobre el estado de conservación de un hábitat, ya que es posible que ciertas especies aparezcan o desaparezcan, sin afectar el equilibrio funcional, pero la desaparición de un rol puede ocasionar cambios en el ecosistema. Como propone Didham et al. (1998) los depredadores son un grupo que se incrementa con eventos de disturbio, siendo un interesante grupo de monitoreo.
Lineamientos de investigación
Es importante corroborar los resultados con otros grupos megadiversos como himenópteros, donde se puedan establecer roles tróficos semejantes, así como replicar este estudio a diferentes rangos latitudinales donde se comparen los resultados. Se sugiere igualmente el tener unidades de muestreo más específicas en el tiempo, realizando recolecciones mensuales, con el objetivo de verificar si las variaciones entre cada uno de los periodos escogidos obedecen a patrones generales o son eventos puntuales.
Deben seguirse acumulando datos de esta localidad, con el fin de poder detectar cambios a una escala temporal mayor. Otro aspecto que podría tenerse en cuenta es la asignación más específica de los roles tróficos, ya que en algunos grupos es algo general, presentándose variaciones a nivel especifico como es el caso de Staphylinidae, Curculionidae y Tenebrionidae, por lo que en un análisis más detallado (morfoespecies, géneros o especies) podrían evidenciarse cambios más sutiles en las dinámicas estacionales.
Se recomienda incluir otros microhabitats no muestreados, que podrían estar albergando otra fauna, especialmente en la época de verano, como es el caso de las bromelias (Paoleiti et al. 1991), o del dosel, donde se ha demostrado su importante aporte a la estructura del ensamblaje (Erwin 1983; Watanabe y Ruaysoongnerm 1989, Lowman et al. 1996). Igualmente se deberían incluir otras áreas de muestreo como la Chagra, donde las dinámicas estacionales pueden diferir e implementar otros métodos de captura no utilizados.
Finalmente este estudio demuestra que existe una fuerte dinámica estacional en la riqueza de las familias encontradas, lo cual puede estar muy relacionado con la dinámica propia de los recursos o con un mecanismo de dilución de la competencia. A pesar de presentarse esta fuerte dinámica estacional, la estructura de los roles tróficos se mantiene relativamente estable, lo cual está señalando la importancia de la regulación de los mismos con respecto a la funcionalidad de un ecosistema; a pesar de aparecer y desaparecer diferentes familias del ensamblaje, siguen existiendo familias que cumplen con los roles necesarios para mantener esta funcionalidad en el ecosistema.
Footnotes
Agradecimientos
A todos los estudiantes que contribuyeron en el trabajo de campo, durante los tres años de muestreo. A la comunidad indígena Monifue-Amena por la hospitalidad brindada durante la elaboración de los muestreos. A dos revisores anónimos y al editor de la revista por todos sus comentarios y aportes los cuales fueron sumamente valiosos para mejorar la calidad del manuscrito. Este trabajo se realizó bajo el convenio No. 1662, entre CorpoAmazonía, Comunidad Monifue-Amena y Pontificia Universidad Javeriana.