
Abstract
Trialeurodes vaporariorum is one of the most important and difficult to control pests of snap beans and dry beans in Colombia. To understand the distribution of the insect on the plant and the preferred feeding sites of immatures and adults, to quantify an action threshold previously established by CIAT, and to develop sampling methods, six trials were conducted in the field in the area of Pradera (Valle). In both snap beans and dry beans, adults colonize and oviposit on the young leaves, then migrate within the plant to the upper levels (strata) as the crop ages, while the nymphs and pupae are found on lower plant strata. Populations of the insect are affected by rainfall. A pattern of clumped distribution on the leaves was found for all stages of T. vaporariorum on snap beans and dry beans. For nymphs, a sample size of 50 leaflets taken at random on lower strata leaves was calculated. The recommended sampling patterns are in the form of an “M” or “Z” that encompass the borders. A sequential sampling plan was developed for first instar nymphs on snap beans based on an action threshold of 12 nymphs/2.25cmβ per leaflet.
Introducción
En el mundo se han identificado cerca de 1200 especies de moscas blancas, principalmente por medio de características morfológicas basadas en diferencias en el cuarto ínstar o pupa (Bink-Moenen y Mound 1990), dentro de estas se encuentra la mosca blanca de los invernaderos Trialeurodes vaporariorum (Westwood) (Hemiptera: Aleyrodidae). Esta especie tiene una distribución cosmopolita y es polífaga con más de cien plantas hospedantes registradas (Byrne et al. 1990). Otra de las moscas blancas más importantes es la mosca blanca de la batata Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae), reconocida como una de las más relevantes en algodón y otros cultivos en Arizona y el sur de California (Naranjo y Flint 1994). Actualmente, esta mosca blanca ha sido más estudiada que T. vaporariorum con relación a su distribución espacial, dinámica de población y métodos de muestreo. Cardona (1995), menciona que en 1984 la mosca blanca de los invernaderos T. vaporariorum hizo su aparición en la Zona Andina como plaga importante de fríjol y otros cultivos; la respuesta inmediata de los agricultores fue la utilización masiva de insecticidas como única alternativa de control. El excesivo uso de agroquímicos, posiblemente en combinación con otros factores ambientales favorables entre 1987 y 1991, originó una explosión masiva de las poblaciones del insecto (Cardona 1995).
En el Valle del Cauca, T. vaporariorum ha sido reportada como plaga principal en cultivos de solanáceas (tomate, tabaco, berenjena y pimentón), cucurbitáceas (zapallo y pepino), crucíferas (repollo y col), cilantro, geranio y leguminosas como fríjol arveja y habichuela, en un rango de altitud de 828 hasta 2040 metros sobre el nivel del mar, lo cual evidencia su marcada polifagia y capacidad de adaptación biológica (Rodríguez y Cardona 2001). Las poblaciones de este insecto han venido incrementándose a medida que aumenta el área sembrada en hortalizas en el Valle del Cauca. La mosca blanca, asociada con Thrips palmi Karny, puede ocasionar pérdidas cercanas al 44.5% en habichuela (Rendón et al. 2001), además de incrementar los costos de producción por el uso excesivo de plaguicidas utilizados para su control (Cardona et al. 1991).
La mosca blanca es un insecto con metamorfosis incompleta (hemimetábolo); su ciclo de vida incluye huevo, cuatro instares ninfales y adulto (Gerling 1992). Los huevos son depositados individualmente o en grupos en el envés de las hojas superiores. Los huevos pueden ser puestos por hembras fecundadas produciéndose machos y hembras; igualmente pueden ser puestos por hembras vírgenes (con partenogénesis del tipo arrenotoquia) donde sólo se obtienen machos (Louise 1975). La ninfa efectúa tres mudas del exoesqueleto. Cuando el adulto emerge, deja un orificio en forma de “T” en la exuvia. Generalmente la relación de los sexos es 1:1 (Vet et al. 1980).
De la Cruz y colaboradores (1990) estudiaron el ciclo de vida de T. vaporariorum en condiciones de laboratorio en fríjol. Encontraron que la duración en días de cada fase del ciclo de vida fue la siguiente: huevo (5.74 ± 2.16), primer instar (2.60 ± 2.16), segundo ínstar (3.10 ± 1.10), tercer ínstar (3.00 ± 1.27), cuarto instar (4.25 ± 1.47) y pupa (3.74 ± 1.99). Las hembras de T. vaporariorum tienen una longevidad de 5 a 28 días, según las condiciones ambientales en que se encuentre la cría (Vet et al. 1980). La fecundidad promedio por hembra a 19°C sobre ICA Pijao fue de 127 huevos en 36 días (Manzano et al. 2000). La temperatura del medio y la planta hospedera ejercen una gran influencia en el tiempo de desarrollo de la mosca blanca. A temperaturas constantes entre 15 – 25°C, el desarrollo de los estados de T. vaporariorum no se ve afectado (Gerling 1990).
El tamaño de los cambios en las poblaciones de los insectos es el resultado de nacimientos, muertes y migraciones. Si estos procesos y los factores que influencian la dinámica de población de un insecto plaga pudieran ser predichos, sería posible diseñar estrategias de manejo de plagas en los cultivos y proporcionaría un avance en la advertencia de epidemias (Lewis 1997).
La distribución espacial de los organismos es una característica biológica importante para muchos ecólogos; las tres formas de distribución son al azar, uniforme o agregada (Ricklefs 1998). Entender los patrones de agregación de los organismos ha sido una tarea recurrente de la ecología de poblaciones y por lo tanto, se han propuesto diferentes modelos matemáticos para describir las distribuciones espaciales (Tonhasca et al. 1994).
Uno de los métodos para determinar la distribución espacial es el de la Ley de Poder de Taylor, en donde el valor de b de la regresión entre el logaritmo de la media de adultos por folíolo y el logaritmo de la varianza de adultos por folíolo pueden considerarse como una medida de agregación (Pedigo y Zeiss 1996). En consecuencia, si se obtiene una pendiente menor a 1, se entiende que el insecto está disperso uniformemente; si es igual a 1, indica dispersión al azar y si la pendiente es mayor que 1, indica agregación.
En cultivos ornamentales como poinsettia (Euphorbia pulcherrima) y crisantemo (Chrysantemum indicum), se encontró que los valores de b fueron significativamente mayores que uno (rango entre 1.5 – 2.5) para todos los estados de las dos especies de mosca blanca estudiadas (B. tabaci y T. vaporariorum), indicando que estos insectos se encuentran de forma agregada (Liu et al. 1993). En tomate bajo invernadero, el conteo de adultos de T. vaporariorum en trampas amarillas con pegante, indicó una distribución agregada cuando estos conteos fueron analizados usando la Ley de Poder de Taylor (Jong-Kwan et al. 2001).
La determinación de los patrones de distribución en un hospedero en particular es un requisito fundamental para desarrollar una técnica eficiente de muestreo de las poblaciones (Lynch y Simmons 1993).
Liu et al. (1993) observaron que en Crisantemo y Poinsettia, la mayoría de adultos, huevos y primeros ínstares ninfales de T. vaporariorum son encontrados en las hojas jóvenes de los estratos superiores y en menor cantidad en las hojas totalmente abiertas. Las ninfas de segundo y tercer ínstar están en las hojas más desarrolladas, totalmente abiertas, menos tiernas, al igual que la mayoría de pupas. El porcentaje de cada estado varía dependiendo del número de hojas de la planta.
En Poinsettia se encontró que, para B. tabaci y T. vaporariorum, la unidad de muestreo puede reducirse a una revisión en el envés de las hojas de los estratos superiores y medio, obteniendo así un estimado para poblaciones dentro de la planta (Liu et al. 1993).
Para estimar poblaciones de insectos, se deben seleccionar unidades de muestreo. El número de unidades de muestra tomadas en un programa es usualmente el resultado de un arreglo entre la precisión, la destreza y el costo (Pedigo 1999). Cuando el patrón de distribución de una plaga es agregado, se necesitan más muestras que cuando el insecto plaga está distribuido uniformemente en el cultivo (Xu Rumei 1982).
Una herramienta de uso potencial en programas de manejo integrado de plagas es el muestreo secuencial o programa de decisión secuencial. Cada programa está basado en un patrón de dispersión del insecto y en niveles económicos de decisión, permitiendo dar acomodo a las poblaciones en una, dos o más categorías (Pedigo 1999). Con el muestreo secuencial, el número total de muestras a ser tomadas en el campo es variable. Usando una gráfica, se puede empezar a muestrear y continuar hasta que la población pueda ser clasificada (Pedigo 1999) como de riesgo o no para tomar decisiones de control.
Dada la importancia económica del insecto y la necesidad de desarrollar planes de manejo para reducir su impacto económico, se hicieron siembras de habichuela y fríjol en las cuales se trató de conocer cómo se desarrollaban las poblaciones de huevos, ninfas y adultos de T. vaporariorum en condiciones naturales de infestación para de esta manera poder definir un método de muestreo. Se describió el daño ocasionado por el insecto a través del período vegetativo y reproductivo del cultivo, sus patrones de dispersión espacial, y se calculó el tamaño óptimo de muestra. También se desarrolló un plan de muestreo secuencial en habichuela para ninfas y se identificaron las mejores formas de muestreo en campo.
Materiales y métodos
Los ensayos se hicieron bajo condiciones de infestación natural de T. vaporariorum en la vereda la Floresta, Municipio de Pradera, Departamento del Valle del Cauca, a una altitud de 980 msnm, con una temperatura promedio de 23°C y 72% de HR. Se sembraron seis ensayos (tres en cada cultivo). Los dos primeros de cada cultivo se hicieron para identificar la forma de dispersión del insecto, establecer un tamaño de muestra y conocer la fluctuación de población de los diferentes estados del insecto en los cultivos de habichuela y fríjol. Cada cultivo se dividió en ocho cuadrantes imaginarios, tal como lo sugieren Pedigo y Zeiss (1996), siendo cada uno de ellos, una repetición para efectos de muestreo. Con la segunda siembra, se logró determinar el mejor método de muestreo en cada cultivo y las dos siembras restantes fueron hechas para cuantificar el umbral de acción de mosca blanca y así, con el tamaño de muestra, lograr establecer un modelo de muestreo secuencial.
Se usó la variedad de habichuela “Lago Azul” por ser la preferida por los agricultores de la zona y la variedad de fríjol “ICA Pijao” por adaptarse bien al medio y ser tolerante a enfermedades. Las distancias de siembra (0.7 m entre surcos dobles y 0.45 m entre surcos sencillos para habichuela y 0.5 m entre surcos y 0.2m entre plantas, para fríjol) y las practicas agronómicas, fueron iguales a las utilizadas por los agricultores de la zona. No se usaron insecticidas, pero se controlaron las enfermedades mediante la aplicación frecuente de fungicidas.
Los recuentos de población se hicieron semanalmente hasta cuando el cultivo llegó al estado de soca. En habichuela, las poblaciones de adultos y huevos empezaron a contarse a los 7 días después de siembra (DDS), etapa V1 (cotiledones abiertos). Las ninfas se contaron a partir de los 14 DDS, etapa V2 (primer trifolio abierto), mientras que las pupas se contaron a partir de los 35 DDS. Los conteos se hicieron en los cotiledones y en el envés de cada folíolo central de cada estrato (unidad de muestreo). Los adultos se registraron por medio de observaciones directas en campo y los estados inmaduros fueron contados en el laboratorio bajo el estereoscopio para diferenciar entre huevos, ninfas y pupas en un área estándar preestablecida de 2.25 cmβ. La planta se dividió en cuatro estratos, por lo que el número de muestras tomadas fue de diez folíolos centrales por estrato y por cuadrante.
En fríjol, las evaluaciones también fueron semanales, comenzando el conteo de adultos a los 8 DDS (Etapa V1), de ninfas a los 17DDS (Etapa V2) y de pupas, a los 32 DDS. El número de muestras tomadas fue de diez folíolos centrales por estrato y por cuadrante. Adicionalmente, para cada cultivo, se hizo un muestreo total que consistió en colectar cuatro plantas completas para registrar los diferentes estados de desarrollo del insecto en cada folíolo de la planta.
Para el análisis estadístico, los datos de recuentos de insectos obtenidos en campo se transformaron a log10 (x + 1). Para hacer el análisis de varianza se tomó la planta como parcela principal y los estratos como subparcelas. Para conocer el sitio de mayor ocurrencia de cada estado del insecto, se hizo separación de medias por DMS al 5%. Se calculó el índice de dispersión del insecto por medio de dos métodos estadísticos; el primero, utilizando la ley de poder de Taylor que relaciona mediante una regresión el logaritmo de la varianza y el logaritmo de la media. Se interpreta el valor de b como índice de agregación así; si b > 1 el índice de dispersión es agregado, si b = 1, la dispersión es al azar, y si b < 1 los datos se ajustan a un patrón de dispersión uniforme. El segundo método utiliza el programa ENSTAT (Pedigo y Zeiss 1996) para estimar el ajuste de la distribución observada a diferentes tipos de distribución teórica tales como binomial negativa.
El tamaño de muestra se calculó para diferentes márgenes de error utilizando la fórmula citada por Bartels et al. (1997) en la cual se incluyen valores a y b obtenidos en la regresión lineal: n = (100/c)β tβ a xb-2. En esta fórmula, n es el número de muestras a tomar; c es el margen de error admitido y t es el valor aproximado de la distribución de t. El plan de muestreo secuencial se calculó utilizando el programa ENSTAT, al igual que los valores obtenidos en la regresión lineal y en el programa de índice de dispersión de ENSTAT.
En la segunda siembra de cada cultivo, se utilizó un tamaño de muestra obtenido con el 15% de margen de error calculado con la fórmula de Bartels et al. (1997). Semanalmente se registraron las poblaciones de ninfas en cuatro formas de muestreo; Z, X, zigzag y M; se calculó la variación relativa (VR), la precisión relativa neta (PRN), el coeficiente de variación (CV) y el tiempo que demora en hacerse cada método de muestreo y así definir la mejor forma de muestreo.
Se hizo una tercera siembra de habichuela y fríjol para cuantificar el número promedio de individuos existentes por folíolo cuando se utiliza el umbral de acción determinado por CIAT para el control de la mosca blanca (Cardona et al. 1993). Para esto se dispusieron cuatro tratamientos en un diseño de cuadrado latino 4 x 4. En todos los ensayos, cuando cada tratamiento llegó a un nivel de infestación predeterminado, se hizo control químico con imidacloprid a la dosis de 0.21 kg i.a. por ha. Los niveles de infestación, a los cuales se hicieron aplicaciones foliares, fueron:
Aplicación de control cuando la mitad de los folíolos presentaran entre el 15% al 30% del área foliar ocupada por ninfas de primer ínstar
Aplicación de control cuando la mitad de los folíolos presentaran entre el 30% al 50% del área foliar ocupada por ninfas de primer ínstar
Aplicación de control cuando la mitad de los folíolos presentaran más del 60% del área foliar por ninfas de primer ínstar
Testigo absoluto (no control)
Para ejercer un control oportuno en cada uno de los niveles de infestación predeterminados, se hicieron recuentos de población desde la aparición de las hojas primarias hasta cosecha, con intervalos de tres días. Las ninfas de T. vaporariorum se contaron en el estereoscopio en un área de 2.25 cmβ en cada folíolo de estratos inferiores y medios de doce plantas escogidas al azar por parcela, con el fin poder cuantificar el número de ninfas que correspondía a cada porcentaje de infestación. Este porcentaje se determinó así: si la mitad de los folíolos presentaba un porcentaje igual o superior al tratamiento asignado, se hacía una aplicación. Se tomaron los rendimientos (estimados en kilogramos por hectárea) en los surcos centrales de cada parcela y éstos se sometieron a análisis de varianza con el programa SAS (1989). Se registraron los gastos incurridos, para obtener los costos de producción de la habichuela y el fríjol, así como su precio por kilogramo. Se hizo la regresión lineal del tipo y = a + bx entre niveles de infestación y rendimientos con el programa Statistix (1998), lo que permitió calcular la función de daño y las pérdidas a cada nivel en términos de producción y porcentaje por los métodos de Schwartz y Klassen (1981).
Con estos valores se procedió a calcular el umbral de acción (U.A.) empleando la fórmula sugerida por Mumford y Norton (1987):
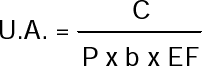
donde:
C = Costo total del control del insecto ($/ha)
P = Precio de la habichuela ($/kg)
b = Función de daño [pérdida en rendimientos (kg/ha/ninfa/2.25cmβ/folíolo), asociada con la densidad de población del insecto]
EF= Eficiencia del control (%)
Resultados y discusión
Fluctuación de poblaciones.
En habichuela el recuento de poblaciones de adultos y huevos se inició a los 7 DDS (etapa V2). A los 29 DDS ocurrió la mayor incidencia de adultos (Fig. 1). Esto se reflejó en un alto volumen de oviposición, posiblemente porque la precipitación disminuyó. Siete días después (a los 14DDS) de la colonización de los adultos, se registraron las ninfas de primer instar provenientes de la primera oviposición. A los 21 DDS (etapa V3), cuando la primera y segunda hoja trifoliada estaban totalmente abiertas y apareció la tercera hoja, la población de ninfas comenzó a incrementarse, sobre todo en las hojas cotiledonales. Las altas precipitaciones en los primeros 23 DDS pudieron ocasionar una disminución en la tasa de colonización de adultos y en consecuencia, reducción en la oviposición y en la emergencia de ninfas (Fig. 1).
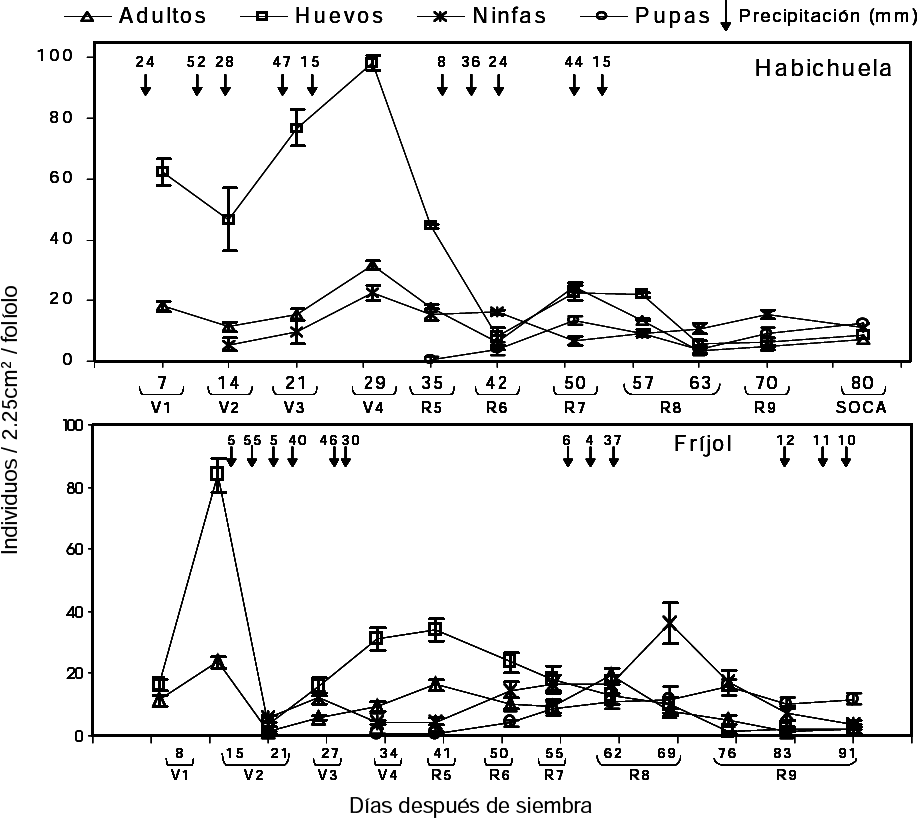
Fluctuación de poblaciones (media + error estándar) de adultos, huevos, ninfas y pupas de Trialeurodes vaporariorum registradas durante el ciclo de cultivo en habichuela (arriba) y fríjol (abajo). V1, V2, R6, etc. se refieren a los diferentes estados de desarrollo de la planta en sus fases vegetativa (V) y reproductiva (R).
En fríjol, el primer recuento de poblaciones se hizo a los 8 DDS (etapa V2). En la Figura 1 se observa que a los 15 DDS se registró la mayor incidencia de adultos de T. vaporariorum, los cuales ovipositaron el mayor número de huevos. La fuerte lluvia disminuyó el número de adultos encontrados en campo y pudo posiblemente desprender gran cantidad de huevos, lo que ocasionó un descenso en la futura población, confirmando que el clima, en especial las precipitaciones, juegan un papel importante en las tasas de natalidad y mortalidad de las poblaciones de mosca blanca. El recuento de ninfas empezó a los 21 DDS, cuando los trifolios primarios estaban totalmente abiertos (Fig. 1). En los dos cultivos ocurrió un descenso en las poblaciones al finalizar el ciclo del cultivo a causa de la migración de los adultos a nuevos hospederos.
Distribución en la planta.
Para conocer la distribución de los diferentes estados de desarrollo de la mosca blanca en los cuatro estratos de la planta, se hizo un análisis de separación de medias, encontrándose que en habichuela, las poblaciones de adultos fueron significativamente mayores en las hojas de los estratos superiores a través del ciclo del cultivo (Tabla 1). Se observó que los huevos se hallaron en el mismo sitio que los adultos, como consecuencia del patrón de colonización y de oviposición de éstos sobre tejidos jóvenes para asegurar el alimento a su descendencia. La población de ninfas en cambio, fue significativamente mayor en los estratos inferiores (1 y 2) (Tabla 1) hasta los 50 DDS, etapa R7. En las etapas siguientes, las ninfas aparecen con mayor frecuencia en el estrato tres a los 57 DDS y en el estrato cuatro a los 63 DDS. Esto se explica por el incremento previo en la oviposición en esta parte de la planta. El número de pupas registradas fue significativamente mayor en el estrato 1 a través del ciclo de desarrollo de la planta hasta los 70 DDS época para la cual la población fue uniforme a través de los cuatro estratos (Tabla 1).
Análisis de separación de medias de poblaciones de adultos, huevos, ninfas y pupas de Trialeurodes vaporariorum encontradas en diferentes estratos de la planta de habichuela a través del ciclo de cultivo. El estrato 1 corresponde al nivel inferior; estrato 2, nivel inferiormedio; estrato 3, nivel superior-medio; estrato 4, nivel superior.
Para cada estado, los promedios seguidos por la misma letra dentro de cada columna no difieren significativamente al nivel del 5% (pruebas de DMS).
En fríjol, los datos obtenidos en campo se analizaron por el método estadístico de prueba de t para detectar diferencias entre los dos estratos en que fue posible dividir la planta. Las poblaciones de adultos y huevos fueron significativamente mayores en los estratos superiores (Tabla 2) a través del ciclo del cultivo. Estas poblaciones disminuyeron a partir de los 69 DDS a causa del daño presentado en las hojas por la fumagina y la madurez fisiológica de la planta. La población de ninfas en el estrato inferior fue significativamente mayor, pues estas hojas fueron preferidas por el insecto para ovipositar en las primeras edades del cultivo. Después de los 55 DDS (inicio de llenado de vainas), las ninfas se encontraron en mayor número en el estrato 2, sitio en el cual ocurrieron las últimas oviposiciones. A los 34 DDS (apertura de la tercera hoja trifoliada), aparecieron las primeras poblaciones de pupas, que fueron significativamente mayores en el estrato inferior hasta los 69 DDS, fecha en la cual la población de pupas fue significativamente mayor en el estrato superior hasta los 91 DDS (cosecha).
Análisis de separación de medias de poblaciones de adultos, huevos, ninfas y pupas de Trialeurodes vaporariorum encontradas en diferentes estratos de la planta de fríjol a través del ciclo de cultivo. El estrato 1 corresponde al nivel inferior; el estrato 2 al nivel superior
Para cada estado, los promedios seguidos por la misma letra dentro de cada columna no difieren significativamente al nivel del 5% (pruebas de t).
Distribución espacial.
Para conocer cómo se encontraban distribuidos espacialmente los diferentes estados de desarrollo del insecto, se utilizaron dos métodos estadísticos: la Ley de Poder de Taylor y la prueba de bondad de ajuste a una distribución Binomial Negativa. En las regresiones obtenidas por el primer método, se observó que la pendiente b en la mayoría de las evaluaciones fue mayor que 1, lo que permite decir que todos los estados de desarrollo del insecto estuvieron distribuidos de forma agregada durante casi todo el período de desarrollo del cultivo. Para esta sección solo detallamos los resultados de adultos y ninfas en habichuela y fríjol. En la Tabla 3 se observa que en el cultivo de habichuela los adultos en campo se distribuyen de forma agrupada (b >1). Al interpretar el cálculo de las frecuencias, se confirma lo encontrado, ya que éstas se ajustan a una distribución binomial negativa, es decir, a un patrón de agregación. Para evaluar la población de ninfas, se cálculo la regresión entre el logaritmo de la media y el logaritmo de la varianza y se encontró que los datos tomados en campo mostraron una distribución agregada debido a que los valores de la pendiente (b) fueron mayores que 1. Al confirmar la información con la ayuda del programa ENSTAT, se encontró que en el cultivo de habichuela, las poblaciones de ninfas de T. vaporariorum se ajustaron a una frecuencia de distribución binomial negativa o agregada (Tabla 4).
Índices de dispersión obtenidos a través de regresión lineal y pruebas de bondad de ajuste a una distribución binomial negativa para poblaciones de adultos de Trialeurodes vaporariorum en épocas críticas de crecimiento del cultivo de habichuela. b = pendiente; X2 = chi cuadrado calculado; Gl = grados de libertad
1 Épocas críticas del cultivo: 14-21 días, desarrollo vegetativo; 35 días, floración; 50-57 días, formación y llenado de vainas; 70 días, final de cosecha. NS, no significativo; *, significativo al 5%; **, significativo al 1%.
Índices de dispersión obtenidos a través de regresión lineal y pruebas de bondad de ajuste a una distribución binomial negativa para poblaciones de ninfas de Trialeurodes vaporariorum en épocas críticas de crecimiento del cultivo de habichuela. b = pendiente; X2 = chi cuadrado calculado; Gl = grados de libertad
1 Épocas críticas del cultivo: 14-21 días, desarrollo vegetativo; 35 días, floración; 50-57 días, formación y llenado de vainas; 70 días, final de cosecha NS, no significativo; *, significativo al 5%; **, significativo al 1%.
En fríjol, como en habichuela, los adultos se distribuyeron en forma agregada (Tabla 5). En esta regresión, se encontró que todos los valores de la pendiente fueron mayores que 1. Cuando se interpretaron los patrones de dispersión por medio del cálculo de frecuencias con la ayuda del programa ENSTAT; se observó que los datos se ajustaron a una distribución binomial negativa, lo cual indica la agregación de los adultos dentro de cada estrato de la planta de fríjol. Las ninfas mostraron similar dispersión que los estadíos anteriores. Cuando se confirmó la información con la interpretación de los patrones de dispersión por medio de las frecuencias, se observó que la población de ninfas se ajustó a una frecuencia de distribución binomial negativa (Tabla 6). A los 62 DDS, algunas de las frecuencias no se ajustaron a esta distribución. Esto posiblemente se debió a una oviposición no agregada que condujo al desarrollo de una población dispersa de ninfas.
Índices de dispersión obtenidos a través de regresión lineal y pruebas de bondad de ajuste a una distribución binomial negativa para poblaciones de adultos de Trialeurodes vaporariorum en épocas críticas de crecimiento del cultivo de fríjol. b = pendiente; x2 = chi cuadrado calculado; Gl = grados de libertad
1 Épocas críticas del cultivo: 15-21 días, desarrollo vegetativo; 34 días, floración; 41-55 días, formación de vainas; 62-76 días, llenado de vainas; 91 días, cosecha. NS, no significativo; *, significativo al 5%; **, significativo al 1%.
Índices de dispersión obtenidos a través de regresión lineal y pruebas de bondad de ajuste a una distribución binomial negativa para poblaciones de ninfas de Trialeurodes vaporariorum en épocas críticas de crecimiento del cultivo de fríjol. b = pendiente; x2 = chi cuadrado calculado; Gl = grados de libertad.
1 Épocas críticas del cultivo: 21-27 días, desarrollo vegetativo; 34 días, floración; 41-55 días, formación de vainas; 62-76 días, llenado de vainas; 91 días, cosecha. NS, no significativo; *, significativo al 5%; **, significativo al 1%.
Refinamiento del umbral de acción.
Para poder desarrollar sistemas de muestreo secuencial en los dos cultivos, primero se cuantificó el umbral de acción desarrollado por Cardona et al. (1993). A través de dos ensayos en habichuela, se encontraron las equivalencias promedio entre la estimación visual del porcentaje de área cubierta por ninfas de primer instar y el número promedio de ninfas de primer instar presentes a través del ciclo del cultivo (Tabla 7).
Equivalencia entre la estimación visual del porcentaje de área ocupada y el número de ninfas de primer ínstar (N1) presentes en un área de 2.25 cm2 de folíolo (promedios de dos ensayos)
En los dos ensayos, la relación entre infestación y rendimiento fue inversa (Fig. 2). Se calcularon las regresiones (Y = 18264 – 158.4X; r = -0.81) y (Y = 23827 –258.1X; r=-0.70). Al comparar estadísticamente las pendientes (b) de las regresiones, se encontró que no hubo diferencia significativa entre ellas (P = 0.29). El umbral de acción (población a la cual hay que ejercer control de la mosca blanca) calculado mediante la fórmula que involucra el valor b de las regresiones, se mantuvo alrededor de 12 ninfas/2.25cmβ/folíolo, a pesar de que la función de daño (b) aumentó como consecuencia en el incremento de la población, el costo de la habichuela varió y el costo para ejercer control sobre el insecto se incrementó. Este número de 12 ninfas/2.25cmβ/folíolo equivale a tener la mitad de las muestras tomadas con el 15% al 30% del área foliar ocupada con ninfas de primer ínstar (Tabla 7).
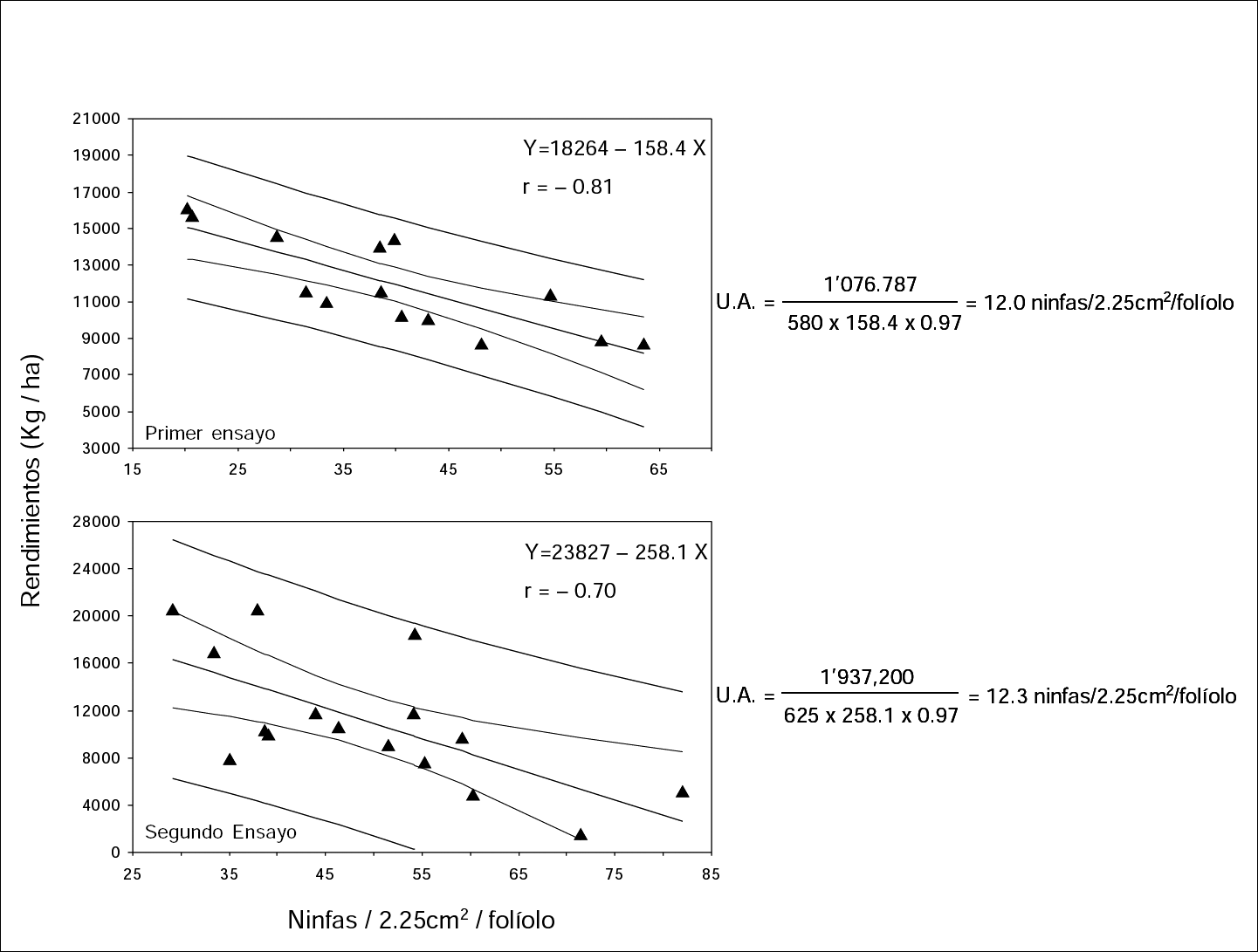
Relación entre los niveles de infestación de ninfas de primer instar de Trialeurodes vaporariorum y los rendimientos de habichuela. Intervalos de confianza al 95%. Cálculo del umbral de acción en dos ensayos.
En cuanto a fríjol, se intentó cuantificar el promedio de individuos por folíolos e incorporar el umbral a un sistema de muestreo secuencial para tomar medidas de control de una manera más fácil. A diferencia de lo observado en habichuela, los rendimientos obtenidos en cada ensayo no guardaron proporción con los diferentes niveles de infestación resultantes del control ejercido a los niveles preestablecidos para cada tratamiento. Ninguna de las dos regresiones fue significativa (P>0.2 y P>0.3). En consecuencia, se puede decir que a diferentes niveles de infestación, el insecto no causó pérdidas económicas significativas. Por esta razón, no se pudo calcular una clara función de daño y por consiguiente, no se debe calcular un umbral de acción. Con este resultado, no se puede desarrollar un plan de muestreo secuencial, debido a que para este cálculo, una condición es tener un valor límite, que para nuestro caso sería el umbral de acción.
Tamaño de muestra y muestreo secuencial.
Se determinó un tamaño de muestra para ninfas, tanto para habichuela como para fríjol, de 50 folíolos en promedio por ha, calculado a partir de los parámetros a y b obtenidos en la regresión cuando se utilizó la Ley de Poder de Taylor para determinar los índices de dispersión de ninfas. La Figura 3 se interpreta de la siguiente forma: si en habichuela se desea encontrar 12 ninfas por folíolo, es necesario tomar 100 muestras con un margen de error del 10%, 50 con un margen de error del 15% y 20 con un margen del 20%. En fríjol, si se desea encontrar 45 ninfas, es necesario tomar 90 muestras con un margen de error del 10%, 40 con un margen de error del 15% ó 20 con un error permitido del 20% (Fig. 3).
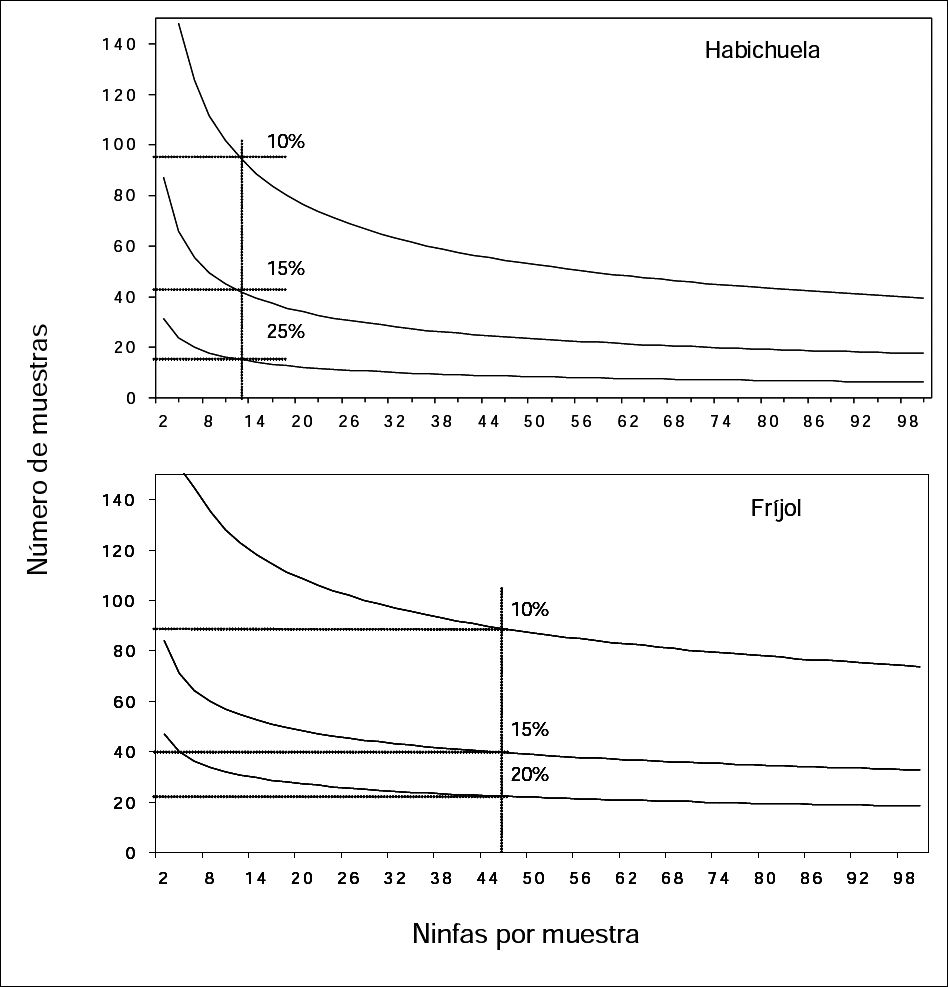
Tamaños de muestra para ninfas de Trialeurodes vaporariorum a diferentes márgenes de error (10, 15 y 20%) en habichuela y fríjol.
Se desarrolló un plan de muestreo secuencial en habichuela. Se ilustra para dos etapas críticas del cultivo (Fig. 4). El sistema permite clasificar las poblaciones de ninfas de T. vaporariorum con respecto al umbral de acción y determinar así si causan daño y por lo tanto, se hace necesario o no realizar un tratamiento de control. En la Figura 4 las “líneas de decisión” que dividen las dos categorías no son muy anchas y si las poblaciones caen en esta zona, es necesario continuar muestreando hasta que se puedan ubicar las poblaciones de ninfas en una de las dos categorías. Por ejemplo (Fig. 4), si en 34 muestras colectadas el número acumulado de ninfas es de 40, el cultivo no necesita ser aplicado. Por el contrario, si con el mismo numero de muestras, el acumulado es de 120 ninfas, se debe ejercer una medida de control de inmediato.
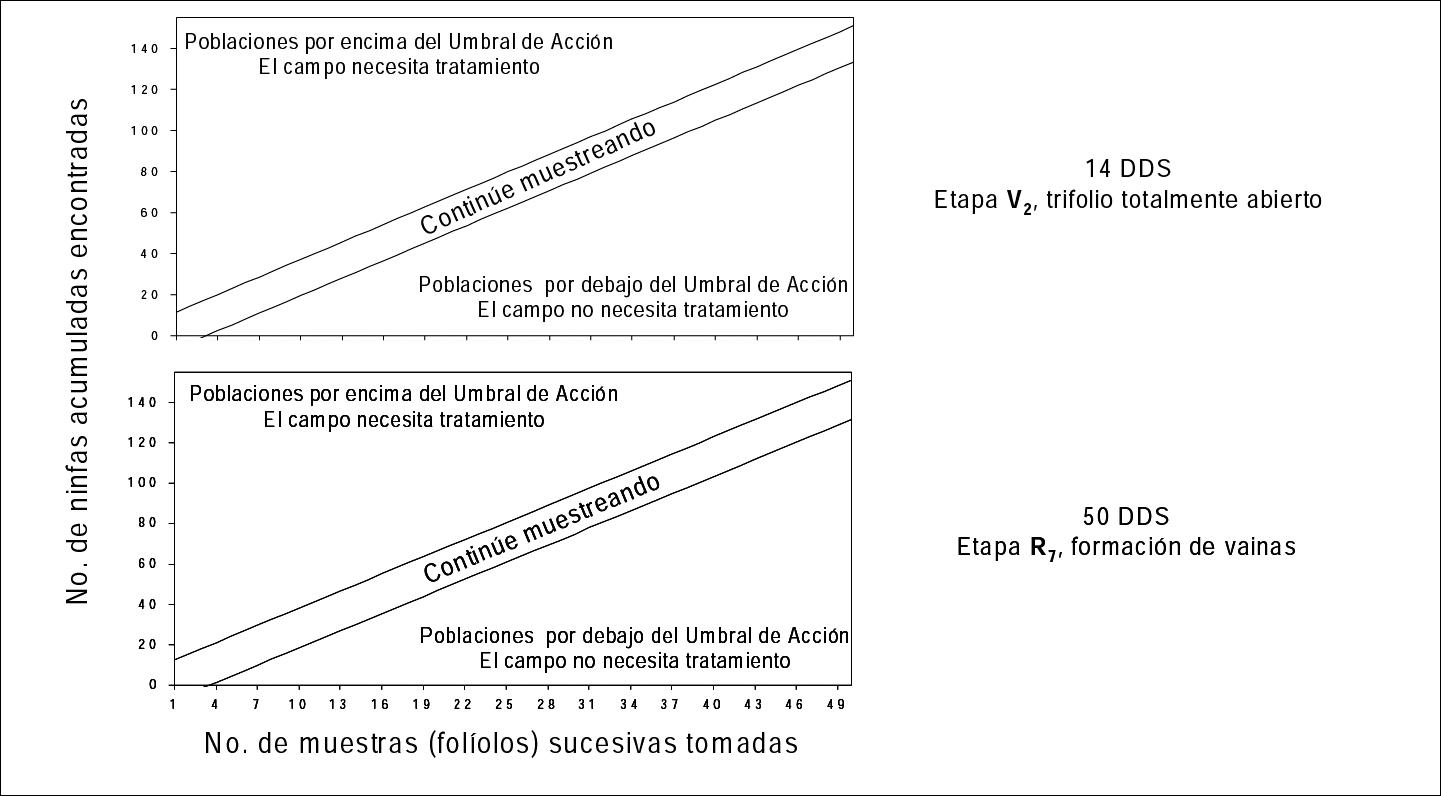
Muestreo secuencial para ninfas de Trialeurodes vaporariorum para dos etapas del cultivo de habichuela.
No se encontraron diferencias entre formas de muestreo en ninguno de los parámetros estadísticos en los dos cultivos (Tabla 8). Como la mayoría de insectos plaga, T. vaporariorum coloniza por los bordes y luego se dispersa a todo el lote; se recomienda recorrer el lote en forma de M al inicio del cultivo y en forma de X después de 19 DDS, para tener un registro confiable de las poblaciones.
Tabla 8. Parámetros estadísticos para seleccionar un método de muestreo para monitorear ninfas de Trialeurodes vaporariorum en habichuela y fríjol
El daño ocasionado en habichuela y fríjol es de tipo mecánico. Además, ocasiona daños indirectos como la fumagina, este daño se proyecta a las vainas, disminuyendo su calidad para el comercio. En ambos cultivos, los adultos y los huevos fueron encontrados en los estratos superiores mientras que las ninfas y pupas se concentran en los estratos inferiores de la planta. Nuestros resultados muestran que, tanto en habichuela como en fríjol, todos los estados de desarrollo del insecto se distribuyen de forma agregada a través del periodo del cultivo. Esto influye en la determinación de los tamaños de muestra y en la escogencia de métodos recomendados para el recuento de poblaciones.