
Abstract
The aim of this research was to construct the life table of the predator Delphastus pusillus (Le Conte) (Coleoptera: Coccinellidae), feeding on its prey, the greenhouse whitefly Trialeurodes vaporariorum (Westwood) (Hemiptera: Aleyrodidae). A group of 30 adults of known age were taken from among individuals reared in a colony maintained in the laboratory, and put into Petri dishes. Based on cohorts of 493 and 682 eggs of the predator, observations were made on the number of individuals alive at the beginning of each development stage, individuals that died in each stage, duration of each stage, female longevity, eggs laid per female, period of oviposition and sex ratio. The information collected was used to calculate the parameters of finite growth rate (λ), intrinsic rate of natural increase (rm), net reproductive rate (Ro), generation time (Tg), time to population doubling (Dt), time of female fecundity, oviposition rate, and the oviposition and survival curves of the predator. There was lower survival of D. pusillus in the early developmental stages, particularly the egg and larval stages. The estimated parameters obtained from the first and sixth generations were: Ro= 5.51 and 3.17, respectively, Dt = 23.0 and 20.5, λ = 1.03 for both generations and rm = 0.03 for both generations. It is suggested that first generation female predators make a greater contribution to female daughters compared to the contribution made by females of the sixth generation maintained in laboratory.
Introducción
Las moscas blancas (Hemiptera: Aleyrodidae) se conocen de tiempo atrás y han sido estudiadas por más de 250 años. De las cerca de 1200 especies de mosca blanca reconocidas e identificadas a nivel mundial, Trialeurodes vaporariorum (Westwood) y Bemisia tabaci (Gennadius) difieren de las otras especies por los graves daños económicos que causan a la agricultura en muchas regiones del mundo (Gerling 2000). En Colombia T. vaporariorum es la especie predominante en el trópico alto y valles interandinos atacando principalmente cultivos de fríjol, habichuela, tomate y papa (Quintero et al. 1999).
Como medidas de control de T. vaporariorum el uso de plaguicidas químicos es casi la única herramienta empleada. A partir de un diagnóstico con agricultores se determinó que el 46% de ellos han tenido que abandonar alguna vez el cultivo por mal control de la plaga, por lo que cuestionan el empleo de insecticidas. El número de ingredientes activos usados contra moscas blanca asciende a 32, siendo los grupos insecticidas más utilizados los organofosforados, carbamatos, piretroides y los de nueva generación (Rendón et al. 2000).
En la búsqueda de alternativas de manejo de la plaga, se han adelantado estudios con el insecto depredador Delphastus pusillus (Le Conte) (Coleoptera: Coccinellidae), el cual se alimenta de las dos especies de mosca blanca de mayor importancia económica T. vaporariorum y B. tabaci y se encuentra ampliamente distribuido en el país en los departamentos de Guajira, Cesar, Santander, Boyacá, Cundinamarca, Tolima, y Huila (López-Ávila et al. 2001).
Antes de liberar un enemigo natural en campo es necesario conocer algunos atributos biológicos y ecológicos que permitan identificar su efecto regulador. Uno de ellos es la capacidad de crecimiento de la población del depredador con respecto a su presa (Rodríguez del Boque 1994). Algunas características de la población en estudio pueden ser obtenidas con la construcción de tablas de vida. La tabla de vida es un componente importante en el entendimiento de la dinámica de población de una especie. Deevy (1947) citado por Southwood (1978), fue el primer investigador que enfocó su atención en la importancia de estas tablas. Las tablas de vida han sido usadas para determinar la expectativa de vida de un candidato biológico y la expectativa de vida de un estado específico del organismo. Ya que muchos insectos tienen generaciones discretas y sus poblaciones no son estacionarias, las tablas de vida “de tipo horizontal o específicas por edades” son más aplicables que las tablas de vida de “temporales o verticales”. Las primeras se basan en la mortalidad real de una cohorte en la que los individuos pertenecen a la misma generación, mientras que las tablas de vida temporales o verticales se basan en la muerte de una cohorte imaginaria encontrada por determinación de estructura de edad en un punto específico de desarrollo de la misma (Southwood 1978; Rabinovich 1980).
En la obtención de una tabla de vida se deben estimar tres parámetros de la población para que esta sea lo más completa posible. El primero es el número de individuos vivos que entran a cada estado del ciclo de vida del insecto; el segundo la mortalidad a partir de factores específicos en cada estado de vida y el tercero la fecundidad (Bellows et al. 1992). Con el presente trabajo se estimaron parámetros de la tabla de vida para Delphastus pusillus y se obtuvieron las curvas de sobrevivencia para el depredador criado en una primera y una sexta generación mantenidas en laboratorio.
Materiales y Métodos
Con el fin de determinar diferencias en los parámetros de la tabla de vida por efecto de las condiciones de cría en laboratorio, se establecieron diferencias para D. pusillus entre una generación reciente y una generación antigua, generaciones I y VI.
A partir de individuos procedentes de la colonia del depredador, se tomó un grupo de 30 adultos de edad conocida, los cuales fueron ubicados por parejas en cajas de Petri de vidrio de 5 cm de diámetro. Al interior de las que se ubicaron foliolos infestados con inmaduros de mosca blanca; los foliolos fueron renovados cada dos días, con el fin de retirar las posibles posturas así como para renovar el alimento al depredador. Las cajas de petri donde se ubicaron las posturas de D. pusillus, fueron debidamente identificadas con la fecha de oviposición y la pareja de adultos de la cual provenían. Con el fin de determinar el comportamiento biológico del depredador en condiciones de cría, se construyó la tabla de vida para una primera y una sexta generación; a partir de una cohorte de 493 y 682 huevos del depredador respectivamente. Se realizaron las observaciones en los diferentes estados por los que pasa el insecto. Se registró el número de individuos vivos al inicio de cada estado, individuos muertos en cada estado, tiempo de duración de cada estado, longevidad de la hembra, huevos puestos por hembra, tiempo en que la hembra fue fecunda y proporción de sexos.
Con estos registros se estimaron los siguientes parámetros de fecundidad del depredador: tiempo de fecundidad de la hembra definido por el tiempo que la hembra ovipositó dividido por su longevidad, tasa de crecimiento reproductivo (GRR), tasa de oviposición promedio día, tasa neta reproductiva (Ro), tasa intrínseca de crecimiento natural (rm) y tasa finita de crecimiento (λ). Posteriormente se obtuvieron parámetros adicionales no relacionados con la fecundidad de D. pusillus: tiempo generacional (Tg), tiempo de duplicación de la población inicial (Dt) y longevidad de la hembra. Finalmente se determinó el comportamiento de sobrevivencia y mortalidad de las poblaciones del depredador en estudio.
La información se analizó con ayuda del procedimiento propuesto por Maia et al. (2000) y el paquete estadístico SAS® (1999). La comparación de los parámetros estimados para las dos generaciones en estudio se realizó con la prueba t de una cola.
Resultados y discusión
La prueba t de una cola presentó diferencias significativas entre las generaciones uno y seis para las variables tiempo de fecundidad, tasa de crecimiento reproductivo (GRR), tasa neta reproductiva (Ro) y tasa de oviposición hembra/día (Tabla 1).
El tiempo de fecundidad, definido como el tiempo de vida en que la hembra ovipositó dividido por su longevidad, para hembras de D. pusillus en la primera generación fue de 69,3 días. En este tiempo, la hembra depredadora presentó una tasa de crecimiento reproductivo (GRR) de 185,3 huevos y una tasa de oviposición diaria de 0,84 (Tabla 1). Las hembras en la sexta generación mostraron un tiempo de fecundidad de 49,0 días, una GRR de 84,2 huevos y una tasa de oviposición hembra/día de 0,61 (Tabla 1).
Para la variable tasa neta reproductiva (Ro) se obtuvo un valor estimado de 5,51 hembras hijas/hembra madre para la primera generación; para la generación seis el aporte de hembras hijas/hembra madre fue de 3,17. Los resultados muestran que hembras del depredador en la primera generación hacen un mayor aporte de hembras hijas respecto al aporte hecho por hembras de una generación seis mantenida en laboratorio.
Parámetros de fecundidad de hembras de D. pusillus en las generaciones uno y seis mantenidas en laboratorio.
Valores significativamente diferentes al nivel del 5%. NS valores no significativos, según la prueba t de una cola
El significado del valor de Ro en ecología de poblaciones, señala el crecimiento, disminución o estabilidad de una población de generación en generación; valores de Ro mayores de la unidad indican que la población en estudio se encuentra en estado de crecimiento, caso observado para la población de D. pusillus, donde dichos valores son mayores que la unidad (Southwood 1978; Rabinovich 1980).
La cría de insectos en condiciones de laboratorio puede generar adaptaciones no genéticas temporales, o cambios genéticos permanentes como resultado de selección, deriva y endocría. Ambos tipos de cambios pueden influenciar el comportamiento del insecto benéfico al ser liberado en campo y puede afectar su desempeño en programas de control biológico. Por ello la implementación de un esquema de control de calidad es importante en un programa de manejo de plagas cuando se utilizan enemigos naturales producidos en laboratorio (De Clercq et al.1998). A partir de la información obtenida se hace necesario determinar a partir de que generación del depredador hay mayor reducción del valor Ro, para introducir poblaciones de D. pusillus de campo y mitigar la disminución de hembras hijas.
La prueba t no detectó diferencias significativas para las variables tasa intrínseca de crecimiento (rm) y tasa finita de crecimiento (λ), en las generaciones de D. pusillus evaluadas. La variable tasa intrínseca de crecimiento (rm), para las dos generaciones presentó un valor de 0,03 hembras/hembra/día; mientras que para la variable tasa finita de crecimiento (λ) el valor fue 1,03.
La igualdad en los parámetros rm y λ estimados para las dos generaciones en estudio se podría explicar por que éstos son considerados intrínsecos de la especie y están regidos por factores genéticos. Al respecto Rabinovich (1980), menciona que rm, como una tasa intrínseca de crecimiento natural, es un parámetro genéticamente determinado y refleja una capacidad potencial de multiplicación de la población.
En condiciones naturales los factores que afectan el equilibrio de una población están dados por el nacimiento, muerte, inmigración y emigración de los individuos. En condiciones controladas los dos últimos factores no son considerados. En condiciones controladas el parámetro rm está regido por la natalidad y la mortalidad de individuos de la población, las cuales para las condiciones de estudio fueron similares.
En términos demográficos el valor de λ está determinado por el ambiente y por el ciclo de vida de las especies. Este parámetro caracteriza la dinámica potencial de una población de una forma simplificada y sirve para discutir efectos de escenarios más complejos. Además sirve como punto de partida de manejo de poblaciones ya sea para reducir la diseminación de la plaga o para incrementar la dispersión de una especie introducida (Neubert y Caswell 2000).
En la mayoría de las hembras evaluadas se observó que la primera postura no fue viable, pero fue de importancia para ubicar sitios de oviposición seguros; algunos sitios usados por la hembra de D. pusillus para proteger sus posturas fueron las nervaduras de las hojas, debajo del cuerpo de adultos de mosca blanca muertos y en el interior de exuvias pupales de mosca blanca; al parecer, este comportamiento reduce la probabilidad de que los huevos sean atacados por otros enemigos naturales.
La curva de oviposición hembra/día (Fig. 1a), para hembras en la primera generación presentó dos picos. El primero a los 32 días y el segundo a los 48 días de vida de la hembra con un promedio de 15,6 y 16,4 huevos/hembra/día respectivamente. Después del día 48 disminuyó la cantidad de huevos puestos, hasta ser cero al último día de vida de la hembra (día 104) La curva de oviposición acumulada para hembras de la primera generación mostró una tendencia sigmoide. Desde el día cero hasta el día 24 de vida de la hembra, se observó un leve incremento de oviposición. Entre los días 30 y 68 de vida de la hembra se presentó un incremento acelerado de oviposición (Fig. 1b), determinado por los mayores picos de oviposición. Después del día 68 de emergida la hembra, se redujo la tasa de oviposición que concuerda con los períodos finales de vida de la hembra (Fig. 1b).
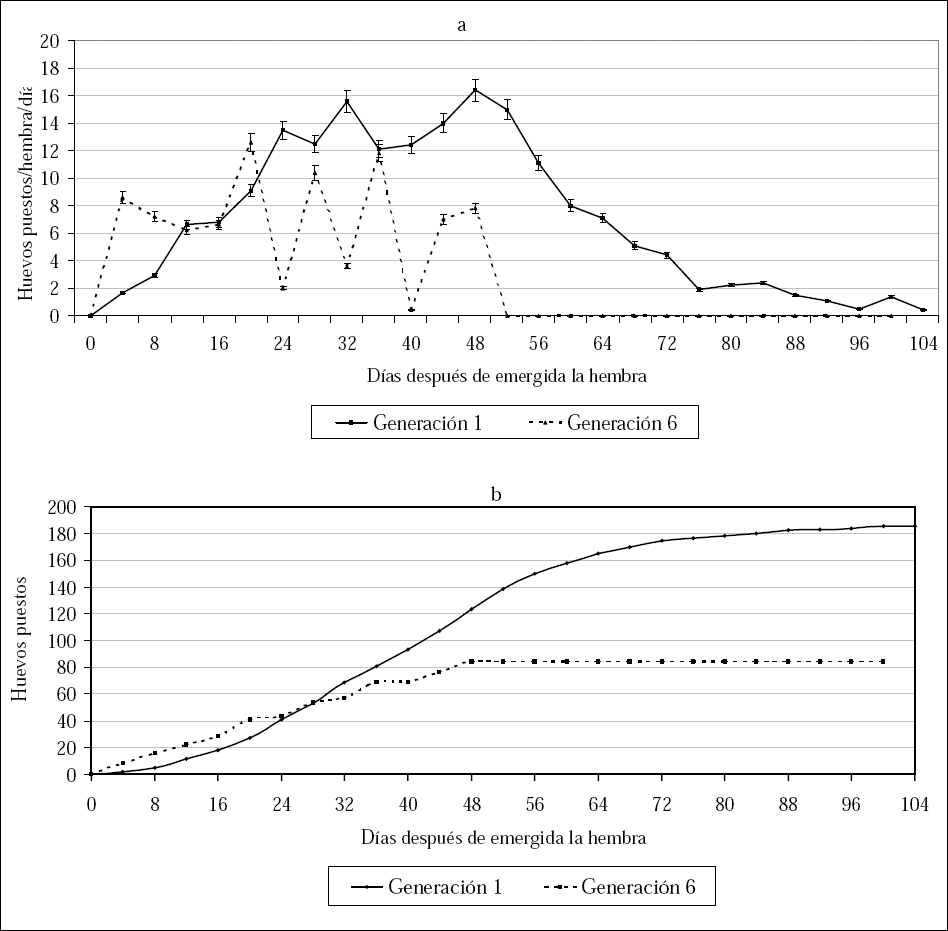
Fecundidad de la hembra de D. pusillus (Le Conte) observada para las generaciones 1 y 6 mantenidas en laboratorio a. Oviposición promedio/día b. Curva de oviposición acumulada.
La curva de huevos puestos/hembra/día para hembras de D. pusillus en una sexta generación mostró picos de alta oviposición seguidos por períodos de baja oviposición (Fig. 1a). Este comportamiento junto al hecho de que la hembra presentó un bajo tiempo de fecundidad respecto a hembras de la primera generación, redujo en gran medida la GRR para la sexta generación (Tabla 1). La curva de oviposición acumulada para hembras de la sexta generación mostró un incremento en la cantidad de huevos puestos desde el día 4 hasta el día 48 de vida de la hembra. Después de esta fecha no se observaron posturas, por lo que la curva se estabilizó hasta la muerte de la última hembra producida al día 100 (Fig. 1b).
A partir de las curvas de oviposición acumulada se evidencia la diferencia que existe en la fecundidad de hembras de la primera y la sexta generación de D. pusillus. La oviposición total acumulada para hembras de la primera y la sexta generación respectivamente fue de 185,3 y 84,2 con un promedio de 1,8 y 0,8 huevos puestos/hembra/día respectivamente. El período de finalización de la oviposición, representado por el tiempo en que la curva se estabiliza, se observó en los días 84 y 48 de emergida la hembra para las generaciones uno y seis respectivamente (Fig. 1b).
Algunos de los factores descritos como limitantes de la fecundidad de especies mantenidas en laboratorio son la calidad y cantidad del esperma producido por el macho (Jervis y Copland 1996). Otro factor limitante es la endocría, en la que algunas características biológicas de la especie como la fecundidad y la proporción de sexos se afectan negativamente, al cruzar individuos con algún parentesco (De Clercq et al. 1998).
El procedimiento seguido para calcular parámetros de la tabla de vida permitió obtener estimativos para otras variables diferentes a la fecundidad y que correspondieron a longevidad, tiempo generacional (Tg) y tiempo de duplicación de la población original (Dt).
La prueba t mostró diferencias significativas para el tiempo de duplicación de la población original, pero no evidenció diferencias para el tiempo generacional y longevidad de la hembra, en las generaciones evaluadas (Tabla 2). De acuerdo con los parámetros estimados D. pusillus en una primera generación requiere 23,0 días para duplicar su población inicial; mientras que el depredador en la generación seis, requiere 20,5 días (Tabla 2). Es probable que D. pusillus trate de remediar la reducción en la fecundidad con la disminución en el tiempo necesario para duplicar la población original. La variable longevidad en las hembras de D. pusillus fue de 82,5 días para hembras en la primera generación. Para hembras de la sexta generación la longevidad de la hembra fue de 79,8 días en promedio (Tabla 2).
Parámetros de la tabla de vida estimados para poblaciones del depredador D. pusillus mantenidas en laboratorio.
Valores significativamente diferentes al nivel del 5%. Ns valores no significativos según la prueba de t de Student.
De los parámetros estimados, el tiempo de duplicación de la población inicial (Dt) fue el único que se vio afectado por las crías sucesivas en laboratorio. Posiblemente existe una relación directa entre (Dt) con los parámetros de fecundidad tasa neta reproductiva (Ro), tiempo de fecundidad, tasa de crecimiento reproductivo (GRR) y tasa de oviposición hembra/día, los cuales fueron significativamente diferentes en las dos generaciones evaluadas (Tabla 1). Como lo muestran los datos, la cría sucesiva del depredador en laboratorio reduce algunos componentes de fecundidad por lo que se debe determinar la generación a partir de la cual se reduce de manera significativa la fecundidad para introducir poblaciones del depredador en campo.
Algunas investigaciones han mostrado la efectividad de la cría continuada de insectos en laboratorio. Cohen y colaboradores (1999), demostraron casos de crías continuadas de insectos en laboratorio por más de 25 años. Se cita el ejemplo del insecto depredador Geocorys punctipes (Hemiptera: Lygaeidae) depredador generalista con el cual se ha mantenido una cría continuada por más de 150 generaciones y la colonia se ha llevado por cerca de 15 años en laboratorio. Para que esta colonia se estabilizara en el tiempo, fue necesario realizar una selección de individuos que permitiera obtener aquellos con capacidades de sobrevivencia en laboratorio. Una vez superado este paso, la colonia del depredador se ha mantenido.
Al analizar la proporción de individuos vivos de D. pusillus durante su tiempo de vida, los resultados mostraron una reducción significativa desde los estados iniciales de desarrollo del insecto; la disminución en la sobrevivencia fue más acelerada en individuos de la sexta generación del depredador respecto a aquellos obtenidos en la primera generación (Fig. 2).
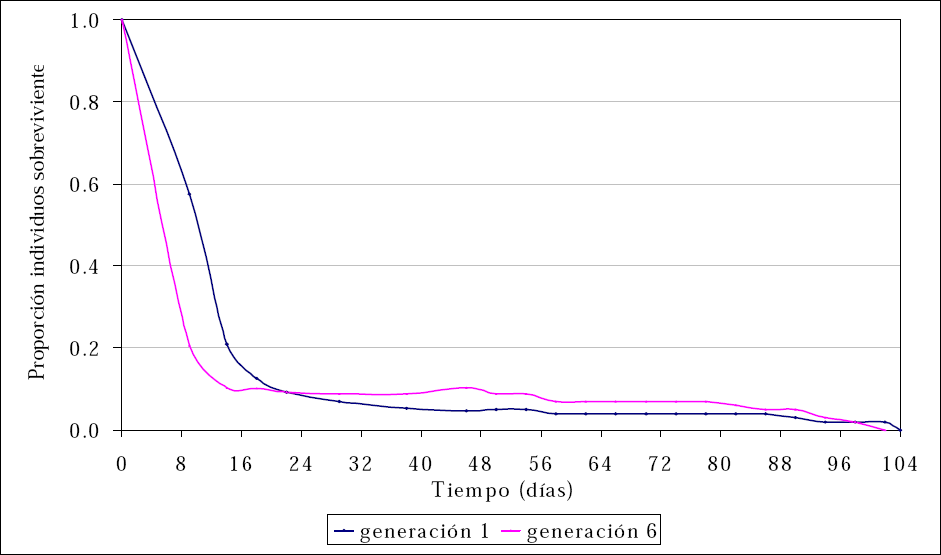
Curvas de sobrevivencia para dos generaciones de Delphastus pusillus (Le Conte) mantenidas en laboratorio.
En la primera generación se observó una reducción en la proporción de individuos vivos al pasar de 1,0 a 0,13 desde el día cero hasta el día 18 de puesto el huevo. Después del día 18 y hasta el día 104 la probabilidad de individuos vivos presentó disminuciones mínimas con 0,13 individuos vivos al día 18, con 0,09 individuos vivos al día 22, con 0,05 al día 54, con 0,04 al día 86 y con 0,02 sobrevivientes al día 102 (Fig. 2). Igual tendencia se observó con individuos de la generación seis, aunque la reducción en la probabilidad de sobrevivientes fue más acelerada al inicio del tiempo de vida de los individuos. En los primeros 14 días, la probabilidad de vida del depredador se redujo de 1,0 a 0,10. Después del día 14 y hasta el día 54, la probabilidad de vida del depredador se redujo de 0,1 a 0,09 individuos vivos. Desde el día 54 al día 78 la probabilidad de vida se redujo de 0,09 a 0,07. A partir de esta fecha la probabilidad de vida se redujo hasta ser cero al último día de vida del adulto.
Los resultados de sobrevivencia de los diferentes estados de D. pusillus, mostraron una reducción importante durante los primeros días de vida del depredador. Este período coincidió con los estados de huevo y larval. Individuos del depredador en una primera generación tienen mayor probabilidad de sobrevivir en los primeros estados de desarrollo del insecto comparado con aquellos de una sexta generación. Individuos de la primera generación tuvieron una probabilidad de vida de 0,21 al día 14; mientras que individuos de una sexta generación tuvieron una probabilidad de vida de 0,10.
La reducción de la sobrevivencia en los primeros días de vida de D. pusillus, se debió a la cantidad de huevos que no fueron fertilizados. Otro factor de mortalidad observado en el estado de huevo se debió en menor grado a huevos depredados por larvas del mismo depredador. Cuando ocurrieron oviposiciones en grupo, las larvas que primero emergieron tuvieron la posibilidad de consumir huevos de su especie.
En el estado de larval las causas de mortalidad observadas fueron intrínsecas y extrínsecas. Aquellas larvas que no tienen alimento en su vecindad salen en búsqueda de presas. Si en el transcurso de 20 a 30 minutos la larva no encuentra alimento, muere. En estos casos la característica de la hoja juega un factor esencial. En el caso de foliolos con gran cantidad de tricomas se interfiere el proceso de búsqueda y encuentro de presas por el depredador ya que se dificulta el contacto entre depredador y presa. Estudios para determinar el efecto de la cantidad de tricomas presentes en la superficie de la hoja sobre el comportamiento reproductivo y alimenticio de D. pusillus, han demostrado que el depredador presenta mayor fecundidad y consumo de ninfas de mosca blanca cuando la hoja posee baja cantidad de tricomas (Heinz et al. 1994).
Aquellos individuos que superaron los estados mencionados tuvieron mayor probabilidad de vida, lo que se evidenció con la estabilidad de las curvas de sobrevivencia (Fig. 2). Después del día 22 de vida y hasta la muerte del último adulto al día 104, individuos de la primera generación presentaron menor probabilidad de vida respecto a la probabilidad observada en individuos de la sexta generación. Este resultado se tradujo en una menor sobrevivencia de los estados de larva2, larva3, larva4, pupa y adulto para individuos de la primera generación.
De acuerdo con los principales tipos de curvas de sobrevivencia propuestos en la literatura D. pusillus asemeja una curva tipo IV, (Fig. 3). Una curva de sobrevivencia tipo IV se caracteriza por que la mortalidad natural afecta fundamentalmente a los individuos jóvenes de la población.
Cuando se han superado las etapas juveniles, la mortalidad natural se reduce en forma considerable y la sobrevivencia de los individuos de la población se estabiliza (Rabinovich 1980; Southwood 1978). En condiciones de cría se hace necesario suministrar suficiente alimento y de buena calidad nutricional al depredador en los primeros instares larvales y facilitar la consecución de presas sanas para la larva recién emergida con el fin de incrementar la probabilidad de sobrevivencia de estos estados.
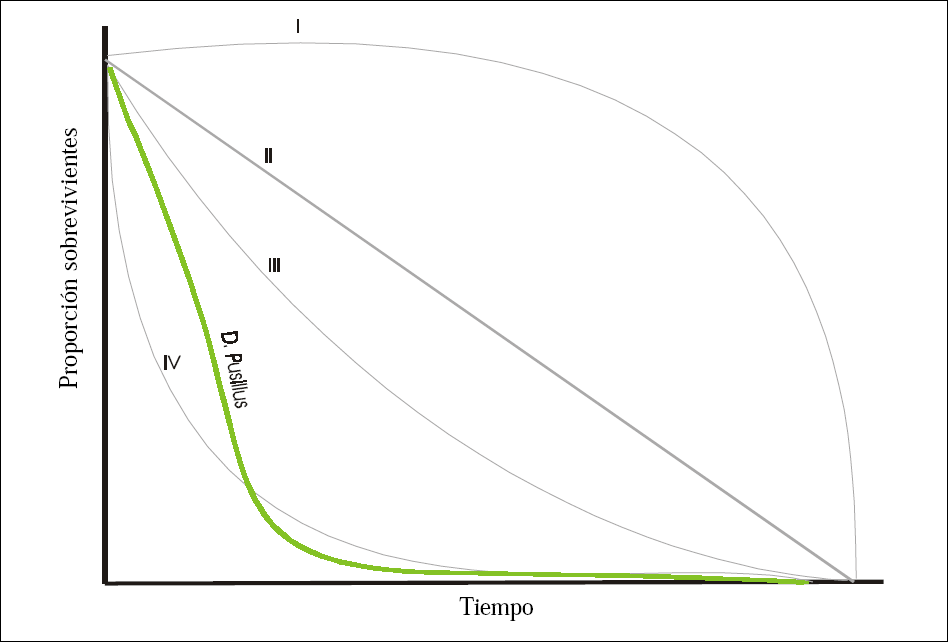
Curva de sobrevivencia obtenida para dos generaciones de Delphastus pusillus (Le Conte) y curvas de sobrevivencia propuestas por Rabinovich (1980) (I-IV).
Al comparar los parámetros estimados para el depredador D. pusillus con los registros obtenidos para otros insectos entomófagos se puede tener una idea del comportamiento del enemigo natural sobre la especie hospedera/presa. Se compararon los parámetros de población de D. pusillus con los registrados en la literatura para los insectos Orius laevigatus (Hemiptera: Anthocoridae) y Eretmocerus californicus (Hymenoptera: Aphelinidae), respectivamente un insecto depredador y un insecto parasitoide (Cocuzza et al. 1997; López y Botto 1997). Los parámetros tasa intrínseca de crecimiento rm, tasa finita de crecimiento λ y tiempo generacional Tg. fueron menores para el depredador en estudio D. pusillus respecto a los enemigos naturales mencionados. Para las variables longevidad, tasa reproductiva de crecimiento (GRR), tiempo de duplicación (Dt) y tasa neta reproductiva Ro, D. pusillus presentó valores mayores comparado con O. laevigatus (Tabla 3).
Como lo muestran los parámetros estimados y presentados en la Tabla 3, insectos depredadores como D. pusillus presentan valores de Ro, rm y λ bajos comparados con los observados por insectos parasitoides, caso específico E. californicus. Estos resultados se explican en el hecho que los parámetros mencionados se fundamentan en la producción de hembras hijas por unidad de tiempo o por generación. Los insectos depredadores requieren del consumo de una cierta cantidad de presas para poder alcanzar su estado adulto y así producir, de acuerdo a la proporción de sexos, una hembra (Hassell 1980).
Comparación de los parámetros estimados para el depredador D. pusillus y los estimados para otros insectos benéficos.
Tomado de Cocuzza et al. (1997)
Tomado de López y Botto (1997)
El hecho de requerir más de una presa para desarrollarse, incrementa el riesgo de mortalidad del depredador en los diferentes estados inmaduros. El efecto biocontrolador de los depredadores se fundamenta en el hecho que tanto larvas como adultos del depredador requieren consumir más de una presa para cumplir con sus funciones vitales. En el caso de la larva para completar su ciclo de vida, mientras que el adulto lo utiliza para funciones reproductivas. Caso contrario ocurre en los parasitoides, donde la relación parasitoide/ huesped es por lo menos 1:1, es decir por cada huesped parasitado se asegura el alimento al inmaduro y la formación de mínimo un adulto (Hassell 1980).
Conclusiones
De los resultados obtenidos en los diferentes experimentos se puede concluir que:
– Los primeros estados de desarrollo de Delphastus pusillus (Le Conte) son los más sensibles a los factores de mortalidad natural. Cuando se superan dichos estados la mortalidad de la población tiende a equilibrarse.
– Características intrínsecas del depredador reflejadas por los valores similares de rm y λ en las dos generaciones evaluadas permiten proponer la cría continuada en laboratorio del depredador.
– Hembras de una primera generación respecto a hembras de una sexta generación hacen un mayor aporte a la población de hembras hijas.
– La ubicación de posturas por la hembra de D. pusillus en áreas con disponibilidad de presas sanas, incrementa la probabilidad de sobrevivencia de las larvas recién emergidas.