
Abstract
The composition of beetles from the family Staphylinidae was studied in ten dry forest fragments located in the departments of Valle del Cauca and Risaralda. Sampling was carried out with a transect system with stations spaced every 20 m. Three collection methods were used at each station: (1) Leaf litter sifting and staphylinid extraction with Winkler sacs, (2) Pitfall traps and (3) Visual searching. A total of 1.463 individuals, grouped in eight subfamilies and 78 morphospecies, were collected in 694 sampling stations. This number of morphospecies corresponds to 10,7% of the species reported in Colombia. Subfamilies Paederinae and Staphylininae are the most dominant in the studied forests. At the regional level, the estimators Chao2, Jacknife 2 and ICE indicated that 90,7, 83,0 and 87,7% respectively of the expected species richness was reported in this study. The Alejandría forest fragment showed the highest diversity with 51 species, followed in descending order by El Medio and El Hatico fragments with 36 staphylinid species each. Complementarity indices showed high and similar values, indicating that species composition among the different fragments is relatively different.
Introducción
Para contribuir con el manejo de los recursos bióticos, es necesario alcanzar un alto nivel de conocimiento de la diversidad biológica. Actualmente, los sistemáticos han descrito entre 1'400.000 y 1'750.000 especies de organismos en el mundo entero y según “Systematics Agenda 2000”, se calcula que faltan por describir entre 10 millones y más de 100 millones de especies (IAvH 1998). Las estimaciones de la fauna a nivel local o regional, basadas en los protocolos de muestreo de los ecólogos o inventarios de los taxónomos, han resultado ser una herramienta valiosa en la dispendiosa labor de evaluar la biodiversidad mundial. Sin embargo, la biodiversidad de Colombia no se conoce en un alto porcentaje y se destruye de manera acelerada (IAvH 1998). La falta de conocimiento dificulta una conservación eficiente y sostenible; de ahí que la necesidad de generar información para evaluar el impacto de la actividad humana en ecosistemas naturales sea uno de los aspectos prioritarios en términos de conservación.
Los artrópodos han sido usados como indicadores de cambios ambientales, debido a sus diversas características y requerimientos ecológicos (Wettstein y Schmid 1999). Entre ellos, los insectos constituyen una proporción sustancial de la biomasa y riqueza de especies terrestres y juegan un papel significativo en el funcionamiento de los ecosistemas; esto ha llevado al desarrollo de diversos estudios que incluyen el uso de especies, taxa superiores, ensamblajes y comunidades de, por ejemplo, libélulas, escarabajos, polillas, mariposas y hormigas en diferentes tipos de hábitat (MacGeoch y Chown 1998; MacGeoch et al. 2002).
El conocimiento actual que se tiene de los Coleópteros de la familia Staphylinidae, particularmente sobre sus preferencias de índole ecológico como tipos de hábitats, distribución y hábitos alimenticios, ha generado el desarrollo de varios estudios con el fin de encontrar métodos que permitan esclarecer la importancia de esta familia como posible bioindicadora, no obstante las dificultades asociadas a la taxonomía del grupo. Se han probado varias metodologías para la aplicación de la familia Staphylinidae como potencial bioindicadora de paisajes naturales y seminaturales (Bohac 1999; Anderson y Ashe 2000).
En Colombia son relativamente pocos los estudios sobre estafilínidos; se conocen algunas especies registradas por Blackwelder (1944), y recientes trabajos locales y de índole ecológico desarrollados por Bernal y Ervik (1996), Arcila (1997), García et al. (2001) y Gutiérrez et al. (2003). Εl estudio preliminar de García et al. (2001) se centró en el bosque seco tropical, uno de los ecosistemas más amenazados del neotrópico (Janzen 1988), y reconocido entre los tres más degradados, fragmentados y menos conocidos de Colombia (IAvH 1998). La región del valle geográfico del río Cauca, en particular, no se ha escapado de este preocupante panorama, pues sus tierras son muy apetecibles por su alta fertilidad y productividad (Alzate 1994). Teniendo en cuenta dos puntos importantes, la destrucción del bosque seco en Colombia y el poco conocimiento de los coleópteros de la familia Staphylinidae en el país, se planteó el presente estudio, que contribuirá al conocimiento del ensamblaje de estafilínidos y al inventario de especies del grupo destacando la importancia faunística de los bosques secos en los departamentos del Valle del Cauca y Risaralda, mediante el análisis de la composición de especies, durante dos años de estudio.
Materiales y Métodos
Área de estudio
El trabajo se realizó en la zona norte del valle geográfico del río Cauca al suroccidente colombiano entre los 4°51’ N, 75°52’ W y los 3°40’ N, 76°17′ W (Fig. 1), comprendiendo los departamentos del Valle y Risaralda, en diez fragmentos de bosque seco tropical (bs-T) no conectados entre sí: El Trapiche (0,64 ha), Córcega (0,79 ha), La Carmelita (1,83 ha), Miralindo II (5,85 ha), El Hatico (7 ha), Miralindo I (7,77 ha), Aguas Claras (9,18 ha), Las Pilas (12,2 ha), El Medio (12,7 ha) y Alejandría (13,44 ha). Los bosques se encuentran en altitudes entre los 970 a los 1.000 msnm, presentan temperatura media superior a los 24°C y precipitación promedia anual entre 1.000 - 2.000 mm (Espinal 1967). La vegetación arbórea en esta zona presenta un dosel alto de 25-30 m y las comunidades terrestres corresponden a fragmentos boscosos heterogéneos que en algunos casos contienen una especie dominante como mantecales (Laetia americana), guaduales (Guadua angustifolia), burilicales (Xylopia ligustrifolia), caracolies (Anacardium excelsum), y en otros son bosques heterogéneos sin una dominancia específica, difíciles de diferenciar y nominados como bosque natural (CVC 1990).
Ubicación de los fragmentos de bosque seco estudiados en los departamentos del Valle del Cauca y Risaralda.
Muestreo de estafilínidos
Se estableció un sistema de transectos lineales en los que se demarcaron estaciones de muestreo cada 20 m. La longitud de los transectos y el número de estaciones varió dependiendo del área total de cada bosque. En cada estación de muestreo se implementaron tres tipos de colecta siguiendo la metodología de García et al. (2001): 1. Escrutinio de cinco litros de hojarasca previamente cernidos en saco miniwinkler; 2. Una trampa de caída (50 ml) con agua jabón expuesta durante 12 h entre diurnas y nocturnas; y 3. Búsqueda directa de estafilínidos, durante 15 min, en vegetación de sotobosque, troncos descompuestos, nidos de hormigas y excremento de animales silvestres. Los estafilínidos capturados se conservaron en alcohol al 70%. En el laboratorio de Entomología de la Universidad del Valle, se separaron por morfoespecies y se determinaron a nivel de subfamilia y género utilizando las claves de Arnett (1963), Dowmie y Arnett (1996), Newton et al. (2001) y Navarrete et al. (2002). Algunos especimenes se determinaron a nivel de especie con la colaboración de A. Newton.
Análisis de datos
Para evaluar la diversidad local (diversidad alfa) en cada fragmento de bosque y la diversidad total o regional (diversidad gamma) en el conjunto de los diez fragmentos, se determinó la abundancia de estafilínidos con base en el número de individuos para cada especie y la riqueza como el número de especies en cada sitio de muestreo. Se calcularon los índices de diversidad de Shannon (H’) y el índice de equitabilidad de Pielou (J); y mediante el análisis de curvas de acumulación de especies, se evaluó la eficiencia de los muestreos usando el programa Estimates 5.1 (Colwell 1997). Para eliminar el efecto de las diferentes técnicas de colecta utilizadas en el estudio así como el orden en el cual se adicionaron las muestras, éstas se aleatorizaron 100 veces. Se emplearon estimadores no paramétricos basados en la incidencia (presencia-ausencia) como Chao 2, Jacknife de segundo orden (Jack 2) y el estimador de incidencia basado en la cobertura (ICE), recomendados para el análisis de grupos hiperdiversos y con distribuciones agrupadas (Longino 1994). Se evaluó la complementariedad de especies entre los fragmentos (diversidad beta) con el índice de complementariedad (IC) propuesto por Colwell y Coddington (1994), como una medida del recambio de especies entre los fragmentos de bosque o entre los métodos de colecta.
Mediante el coeficiente de correlación de Spearman (rs), se relacionó la riqueza de especies con el área de cada fragmento. Se utilizó el programa Statistica 6.0.
Resultados
Composición general de estafilínidos
En las 694 estaciones de muestreo se colectaron 1.470 individuos, agrupados en ocho subfamilias y 78 morfoespecies (Tabla 1). Las morfoespecies encontradas se determinaron a nivel de subfamilia en un 100%, a nivel de género en un 82% y tan sólo el 18% a nivel de especie. La cifra anterior es una consecuencia de la falta de estudios de este grupo en el país, sumado a la taxonomía difícil que caracteriza a la familia Staphylinidae.
Especies de la familia Staphylinidae coleccionadas en diez bosques secos del suroccidente colombiano. Tra: El Trapiche; Car: La Carmelita; Cor: Córcega; Mirl: Miralindo I; MirII: Miralindo II; AgCl: Aguas Claras; Hat: El Hatico; Pil: Las Pilas; Med: El Medio; Alej: Alejandría
Las subfamilias Staphylininae y Paederinae presentaron una amplia dominancia (20 especies), reducidas a casi la mitad en las subfamilias Aleocharinae (12 especies) y Osoriinae (9 especies), seguidas por Tachyporinae y Oxytelinae (6 especies); y con una pequeña representación se observaron Steninae (3 especies) y Piestinae (2 especies).
Entre las especies más abundantes, a nivel regional, se registraron Coproporus sp.1 con 260 individuos colectados, Aneocamp-tus excisicollis con 176 individuos, Anotylus sp.1 con 129 individuos, Aleo-charinae sp. 7 con 78 individuos, Copro-porus sp. 2 con 61 individuos y Xenopigus analis con 54 individuos. Igualmente, los bosques que aportaron un número mayor de individuos a la fauna total de estafilínidos de la región estudiada fueron Alejandría (456), El Medio (385), El Hatico (201) y Aguas Claras (154).
Riqueza de especies
Se encontró una correlación significativa entre la riqueza de especies de estafilínidos y el área de los fragmentos de bosque (rs = 0,847, g.l.= 10, P<0,005) (Fig. 2). En la tabla 2 se indica el número de especies observadas y los valores de los estimadores (Chao 2, ICE y Jack 2) para cada uno de los fragmentos y para el total de los fragmentos. A nivel local, el fragmento de Alejandría que tiene la mayor extensión (13,44 ha), presentó la riqueza más alta con 51 especies observadas y 67,2 especies estimadas (Chao 2). Le siguen los fragmentos de El Medio (12,7 ha) y El Hatico (7 ha), con 36 especies (Chao 2 = 41,1 y 38 especies, respectivamente). Este último fragmento sorprendió por presentar un número de especies superior al de otros bosques con áreas mayores. En contraste, en el fragmento de Las Pilas (12,2 ha), con un área casi similar al de El Medio, se observó una riqueza baja de especies (13). El número de especies más reducido se encontró en los bosques con áreas menores, pero sobresalió el bosque ubicado en la hacienda El Trapiche (de sólo 0,64 ha), por presentar un número de especies igual al de bosques de tamaño medio como Miralindo II y un número alto de especies raras (ocho). A nivel regional, la riqueza observada fue de 78 especies y la riqueza esperada, mediante el estimador Chao 2, predice que aproximadamente ocho especies adicionales deberían haber sido encontradas en el estudio (Chao 2 = 86). Otros estimadores, como ICE y Jack 2, presentan un comportamiento similar donde se predice que respectivamente 10 y 16 especies adicionales deberían haber sido encontradas (ICE = 88,9; Jack 2 = 94).
Valores observados y estimados de riqueza y diversidad de especies de estafilínidos en los bosques muestreados
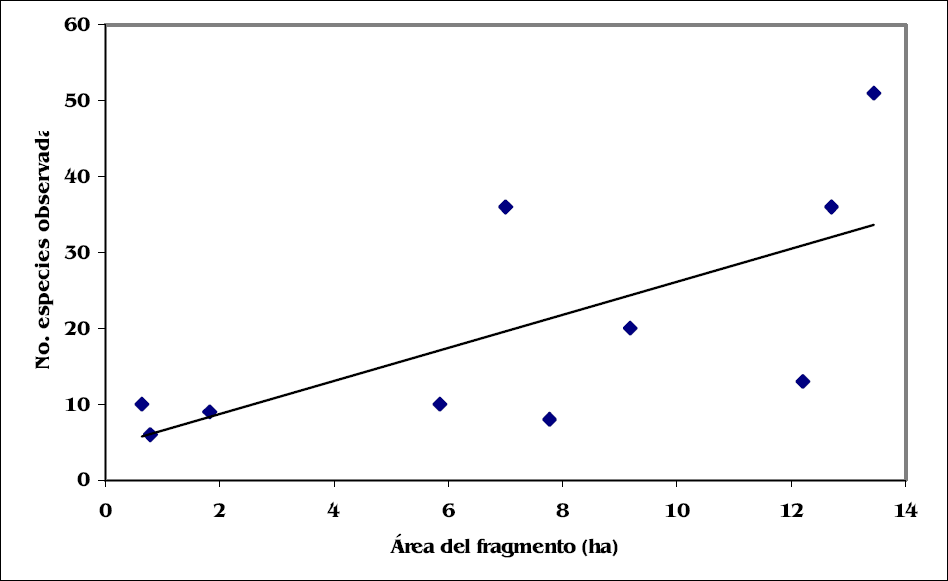
Relación entre la riqueza de especies de Staphylinidae y el área de los fragmentos de bosque seco.
Las curvas de acumulación de especies y el comportamiento de los estimadores de riqueza, para los diez fragmentos muestreados, se observan en la figura 3. A nivel local, el estimador Chao 2 arrojó un porcentaje de eficiencia que varió entre un mínimo de 62%, acompañado de un número alto de especies representadas por un sólo individuo, para el bosque de Las Pilas (Fig. 3H); y un máximo de 95% para el bosque de El Hatico (Fig. 3G). A nivel regional, con el mismo estimador Chao2, el 91% de la riqueza esperada fue registrada en el muestreo y aproximadamente el 37,2% de las especies pueden ser catalogadas como especies “raras” o poco abundantes (15 únicos y 14 duplicados) (Fig. 3K).
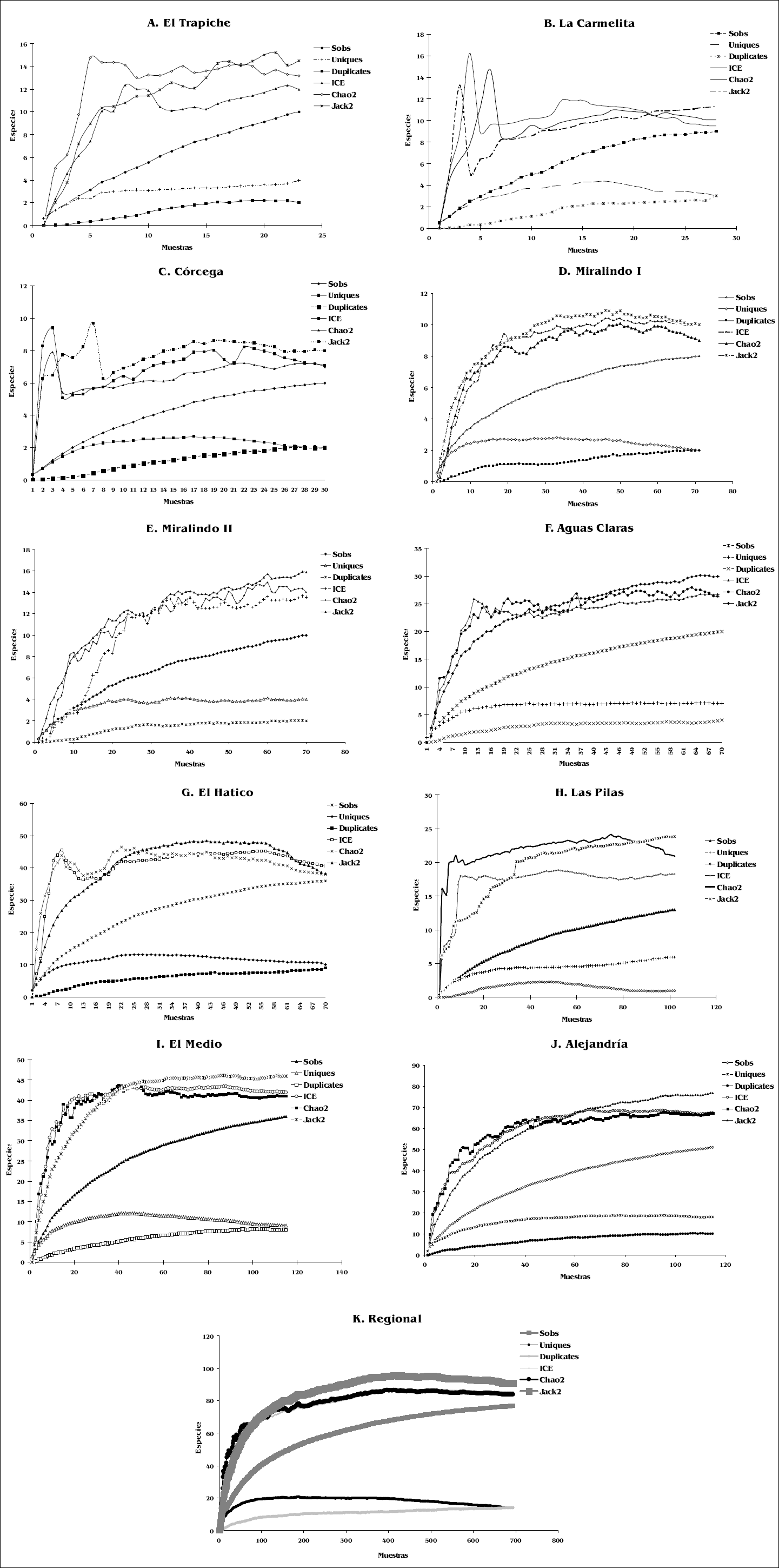
Curvas de acumulación de especies observadas y de los estimadores no paramétricos de riqueza para los fragmentos de bosque seco.
El índice de diversidad de Shannon presentó los valores más altos en los fragmentos de Alejandría (H'= 3,34), El Hatico (H’ = 3,27) y El Medio (H’ = 2,89). Los datos anteriores concuerdan con los obtenidos por los estimadores Chao2, ICE y Jack2, los cuales ubican estos mismos bosques como los de mayor riqueza, en orden de mayor a menor se presentan Alejandría, El Medio y El Hatico (Tabla 2).
Complementariedad entre los fragmentos
Los valores del índice de complementariedad o el grado con el cual cada fragmento de bosque aporta a la diversidad total o regional se observan en la tabla 3. En general los valores del IC al comparar entre pares de bosques presentan valores en su mayoría cercanos a uno (1); es decir que la composición de especies entre los bosques estudiados es relativamente diferente. Los valores más altos del IC se observan al comparar los bosques El Trapiche con Las Pilas (IC= 0,96), Miralindo II y La Carmelita (IC= 0,95) y los bosques El Trapiche y Córcega (IC= 0,93). Los valores más bajos se registraron al comparar Miralindo I y Miralindo II (IC= 0,57), El Hatico y El Medio (IC = 0,57) y El Hatico con Alejandría (IC= 0,58). Los bosques de Alejandría, El Medio y El Hatico comparten un número considerable de especies entre 22 y 26 especies de 78 totales, que representan aproximadamente el 30% de todas las especies registradas en la región. Los fragmentos más pequeños (El Trapiche, Córcega y La Carmelita), a pesar de estar ubicados muy cerca entre ellos, comparten un número reducido de especies, en parte debido a la diversidad baja encontrada en estos bosques, indicando que no existe una recolonización rápida que garantice un número mayor de especies compartidas entre bosques cercanos.
Complementariedad observada de estafilínidos entre pares de diez fragmentos de bosque seco. Tra: El Trapiche; Car: La Carmelita; Cor: Córcega; Mirl: Miralindo I; MirII: Miralindo II; AgCl: Aguas Claras; Hat: El Hatico; Pil: Las Pilas; Med: El Medio; Alej: Alejandría
Discusión
Los especímenes registrados en este estudio corresponden aproximadamente al 10,7% de las 732 especies de estafilínidos conocidas para Colombia (Blackwelder 1944, A. Newton com. pers.). Esta cifra es considerable si se tiene en cuenta que los fragmentos analizados cubren un área total de solo 45,56 ha y que el muestreo abarcó solamente el sotobosque. Se aprecia una dominancia de las subfamilias Staphylininae y Paederinae confirmando los resultados previos de García et al. (2001) y las descripciones de Newton et al. (2001). El género Coproporus resultó el más abundante; según Campbell (1975), este género se caracteriza por estar ampliamente distribuido en áreas tropicales de todas las regiones del mundo y se asocia a material vegetal en descomposición, tal como lo observado en los fragmentos de bosque seco, donde las mayores colectas de Coproporus se lograron con el cernido de la hojarasca y la búsqueda directa en troncos caídos.
La estimación de la riqueza de estafilínidos, mediante las curvas de acumulación de especies, reveló que a nivel regional, un porcentaje alto de especies se colectó en el estudio (entre 83 y 90%). El porcentaje de especies que aún falta por coleccionar, podría obtenerse al cubrir un área mayor de muestreo, no obstante el objetivo perseguido podría no compensar el esfuerzo de muestreo y costos del estudio invertidos. Lo anterior se apoya en la comparación del trabajo preliminar de García et al. (2001), quienes implementaron los mismos métodos de captura de estafilínidos en nueve de los diez bosques objeto del presente estudio. Obtuvieron 50 morfoespecies y 237 individuos extraídos de 245 muestras; respecto a estas cifras, en el presente estudio el esfuerzo de muestreo fue casi triplicado, la abundancia de individuos aumentó seis veces pero la lista de especies no alcanzó a duplicarse, aumentando en un 56% (28 morfoespecies).
La comparación entre el área de cada fragmento y el número de especies observadas arrojó una correlación significativa positiva, lo que concuerda con el trabajo de Kotze y Samways (1999), quienes encontraron pocas especies de estafilínidos en parches de bosques afromontanos pequeños (2,5-3,5 ha), y un número alto de especies en parches medianos (<6 ha). Estos resultados son muy importantes si se tiene en cuenta la teoría de biogeografía de islas (Morrone et al. 1996), pues una de las características de los fragmentos que se analizaron es que no están conectados, presumiéndose un aislamiento considerable entre ellos, además de presentar diferentes tipos de matrices circundantes. Los fragmentos más grandes, como Alejandría y El Medio, tuvieron un mayor número de especies mientras que los bosques pequeños se caracterizaron por presentar pocas especies.
Como una excepción a la relación significativa área-especies antes mencionada, sobresalen tres fragmentos: Las Pilas, El Hatico y El Trapiche. Las Pilas presentó un número pobre de especies con relación a su área (12,2 ha), resultado que puede estar afectado por las dificultades en el desarrollo de las técnicas de muestreo en este bosque invadido por chusquea, lo cual va acompañado de una producción baja de hojarasca y acceso difícil a diferentes sustratos para la búsqueda directa. En contraste, El Hatico (7 ha) y El Trapiche (0,64 ha) presentaron una riqueza alta de especies en comparación con otros fragmentos de área similar o mayor. Con relación al bosque El Hatico es importante considerar la heterogeneidad de la matriz circundante, constituida por potrero arbolado, caña de azúcar orgánica, frutales, guadua y zona en regeneración, lo cual podría estar favoreciendo un flujo de especies entre los diferentes elementos del paisaje. La mayor diversidad de estafilínidos encontrada en el Hatico, es consistente con los resultados obtenidos sobre riqueza (66 especies) y diversidad de hormigas (H’ = 3,52) para este mismo fragmento, en comparación con fragmentos de casi el doble de extensión como Las Pilas (50 especies; H'=3,27) (Armbrecht y Chacón de Ulloa 1997; Armbrecht y Ulloa-Chacón 1999). En el caso de los trabajos adelantados con hormigas, las mismas autoras intentan explicar sus resultados enfatizando en el manejo antropogénico de El Hatico, continuamente protegido por sus dueños. Con respecto al Trapiche, un fragmento muy pequeño (0,64 ha), es probable que sea afectado por efectos de borde que facilitan la entrada de individuos provenientes de la matriz (Ås 1999), dando como resultado un número relativamente alto de especies al interior del bosque.
A nivel regional, se observa una mayor riqueza de especies (diversidad gamma) en comparación con la riqueza de cada fragmento (diversidad alfa); igualmente, el índice de diversidad de Shannon tuvo un valor más alto a nivel regional que a nivel local; estos datos resaltan la importancia de las especies propias de cada fragmento como un aporte a la fauna de estafilínidos del valle geográfico del río Cauca. El número de especies raras, con uno o dos individuos, se mantuvo constante a medida que aumentaban las estaciones de muestreo, para la mayoría de los bosques. Considerando el total de los bosques, el 37,2% de las especies colectadas fueron catalogadas como raras; esta proporción es considerable si se tiene en cuenta la vulnerabilidad de estos fragmentos en un ambiente altamente antropogénico.
Los índices de complementariedad (diversidad beta) mostraron valores medios-altos, es decir que el número de especies compartidas entre pares de bosques es relativamente bajo, presentándose un mayor número de especies compartidas entre los fragmentos Miralindo I y Miralindo II que casualmente son los bosques más adyacentes; al igual que entre los bosques El Medio y El Hatico, dos de los tres fragmentos ubicados en el departamento del Valle del Cauca, los cuales comparten además un tipo similar de matriz circundante (caña de azúcar). Lo anterior destaca la importancia de desarrollar estudios con estafilínidos incluyendo distribuciones de especies en áreas agrícolas circundantes, o en otras áreas antrópicas que conforman el paisaje fragmentado.
Conclusiones
En los fragmentos de bosque seco localizados en los departamentos del Valle del Cauca y Risaralda, se registró aproximadamente el 10,7% de las especies de estafilínidos conocidas para Colombia.
En el ecosistema de bosque seco tropical se encontró una mejor estimación de la riqueza de especies de coleópteros estafilínidos a escala regional (conjunto de diez fragmentos de bosque) que a escala local (cada fragmento).
A pesar de las dificultades en la determinación de estafilínidos, se halló una correlación significativa entre la extensión de los fragmentos de bosque y la riqueza observada; se tuvo una proporción considerable de especies raras y un número bajo de especies compartidas entre los fragmentos, indicando la vulnerabilidad en las poblaciones de estafilínidos en los bosques estudiados.
Los resultados obtenidos en este estudio aportan una valiosa información taxonómica y ecológica sobre la familia Staphylinidae en un ecosistema fragmentado, como base para posteriores trabajos de planeamiento y manejo de la biodiversidad.
Se recomienda continuar desarrollando estudios sobre los estafilínidos de Colombia, orientados a resolver problemas en la taxonomía y a perfeccionar métodos de muestreo para su posterior uso como grupo bioindicador.
Footnotes
Agradecimientos
Las autoras agradecen al taxónomo A. F. Newton Jr. (Field Museum of Natural History, Chicago); a J. L. Navarrete-Heredia por la donación de material bibliográfico; a I. Armbrecht por su apoyo en la primera fase del trabajo, a los propietarios y administradores de los bosques por permitir el trabajo en sus predios, C. Avila, M. Botero, G. Franco, H. Hincapié, G. Jaramillo, P. Salazar, H. Sanint y P. Silverstone-Sopkin. A las entidades cofinanciadoras del estudio: Instituto Colombiano para el Desarrollo de la Ciencia y la Tecnología, COLCIENCIAS (proyecto PNCMАН 1106-13-197-96) y Programa de becas FES-IAvH.