
Abstract
The effect of the entomopathogenic fungus Beauveria bassiana (strain UdeA13) was evaluated on fifth instar nymphs of Rhodnius prolixus under laboratory conditions. The strain was isolated from R. pallescens in the region of San Onofre (Sucre) and cultivated on Sabouraud Dextrosa Agar (SDA). The insect colonies were propagated and reared under controlled conditions of temperature (25-27°C) and relative humidity (80%) in darkness for growth and reproduction. The insects were feed weekly for 1 h with hen's blood (Gallus gallus). The treatments consisted of conidial suspensions of the entomopathogenic fungus B. bassiana at concentrations of 3x105, 1x107, 3x108 y 3x109 conidia/ml which were sprayed on the insects using the Pulverization Tower emulator system. Control insects were sprayed with an aqueous solution of Tween 80 (0,01%). Effects were compared among fungus treatments and the controls. Significative differences (P<0.01) were found among all treatments, regardless of whether the control group was included in the analysis. The concentrations of 3x108 y 3x109 conidia/ml produced 90 to 100% of mortality between 7 and 9 days after treatment. The results of this research indicate that B. bassiana (UdeA13) can be considered as a possible agent for the biological control of the vector R. prolixus.
Introducción
Rhodnius prolixus (Stahl) es un insecto hematófago, vector del protozoario flagelado Trypanosoma cruzi, causante de la enfermedad de Chagas, una parasitosis crónica que constituye un grave problema de salud pública en Latinoamérica (Botero y Restrepo 1998; Días y Schofield 1999; Lecuona et al. 2001).
En Colombia, la enfermedad de Chagas constituye una de las mayores causas de incapacidad en la población económicamente activa; afecta el 7% de la población y cerca del 23% se encuentra en riesgo de adquirir la infección (Guhl y Nicholls 2001). Se calcula que debido a esta enfermedad en el país se pueden estar perdiendo alrededor de 150 millones de dólares anuales (Schofield 1997). La transmisión vectorial, el medio más importante para adquirir la infección, se da principalmente por insectos de los géneros Rhodnius y Triatoma. La especie R. prolixus es la más importante debido a su carácter domiciliar, capacidad vectorial, amplia distribución en el país y a que se encuentra adaptada a ambientes silvestres y peridomésticos. También es un vector importante en Centroamérica y parte de Sudamérica (Lent y Wygodzinsky 1979; Moreno 1997; Ministerio de Salud 1998; Calle 2000; Guhl y Nicholls 2001; OMS 2002).
El insecto posee cinco estadios ninfales, todos hematófagos estrictos. Estudios realizados en laboratorio, basados en el índice de excreción de todos los estadios, mostraron que el quinto es el vector más eficiente del parásito, seguido por hembras adultas y ninfas de cuarto estadio (Miwako y Edmanb 2001). Las poblaciones de insectos domiciliados tienden a ser genéticamente homogéneas, lo que sugiere que la respuesta a los diferentes métodos de control es la misma para todos los insectos (Moreno 1997; Días y Schofield 1999; Schofield 2000).
Organismos como ácaros, nemátodos, microhimenópteros y hongos se han utilizado en pruebas de laboratorio con el fin de observar su efecto en el control de poblaciones de insectos plaga (Ferron et al. 1991; CIDEIM 1994; Uribe et al. 1997). ΕΙ hongo entomopatógeno Beauveria bassiana (Bals.) Vuill ha sido considerado como agente potencial en el control de triatomineos por su amplia distribución y diversidad de hospederos (Romaña 1992; Arroyave 1995; Luz et al. 1998b; Fargues y Luz 2000; Lecuona et al. 2001, Pineda et al. 2002, 2004).
Romaña y Fargues (1992) evaluaron la susceptibilidad de diferentes estadios del insecto vector R. prolixus a la acción del hongo B. bassiana, encontrando excelentes resultados y considerándolo como un agente promisorio en el control biológico. Fargues y Luz (1998) demostraron el potencial epizoótico de B. bassiana para el control de R. prolixus considerando los factores abióticos que afectaban la conidiación sobre los insectos, obteniendo que la humedad relativa era el factor más importante. Pineda et al. (2002) probaron la susceptibilidad de R. ecuadoriensis (Lent y León) a la acción de la cера B. bassiana UdeA13 obteniendo altas tasas de mortalidad en estos insectos. Por esta razón en este estudio se consideró que el hongo B. bassiana UdeA13 tiene un efecto significativo a diferentes concentraciones en la disminución de la supervivencia de R. prolixus de V estado de desarrollo y en la conidiación del hongo sobre los cadáveres.
El objetivo de este trabajo consistió en evaluar la acción del hongo B. bassiana UdeA13 sobre ninfas de quinto estado de desarrollo de R. prolixus bajo condiciones de laboratorio.
Materiales y Métodos
Origen de los insectos
La población de R. prolixus utilizada se obtuvo de una colonia procedente de San Onofre (Sucre) y mantenida en el laboratorio de Micología, del Instituto de Biología de la Universidad de Antioquia. Se criaron dentro de cristalizadores plásticos de 288 cm³ con tapa plástica y un agujero central cubierto de muselina, al cual se le introdujo una lámina de cartón paja para ampliar el área de desplazamiento de los insectos. Los recipientes se mantuvieron en cámara climatizada (WTBbinder 78532 Tuttlingen/Germany), a una temperatura de 27°C y a una humedad relativa del 80%. Los insectos se alimentaron con sangre de gallina (Gallus gallus) dos veces por semana.
Hongo Entomopatógeno
La cepa del hongo B. bassiana UdeA13 utilizada en este estudio fue aislada de Rhodnius pallescens Barber infectado naturalmente, en la región de San Onofre, Sucre. Fue repicada en Agar Sabouraud Dextrosa (Oxoid Ltda., Basinastoke, Hampshire) cada 15 días a partir de insectos conidiados, e incubada a temperatura ambiente (25-27°C) en el laboratorio de Micología, Instituto de Biología, Universidad de Antioquia.
Bioensayo
Se preparó una suspensión conidial del hongo B. bassiana por raspado de la superficie del cultivo con una barra de vidrio estéril y 100 ml de solución acuosa estéril de Tween 80 (0,01%) (Sigma, St. Louis, MO, USA). La concentración fue determinada mediante el conteo de conidios en la cámara de Neubauer (Boeco-Germany). Para el bioensayo se probaron cuatro concentraciones de la cepa B. bassiana 3x105, 1x107, 3x108 y 3x109 conidios/ml. Se realizaron cuatro repeticiones con cada una de ellas y como testigo se utilizó una solución acuosa de Tween 80 (0,01%). Las dosis elegidas fueron seleccionadas con base en los resultados de estudios previos realizados por Romaña (1992), Fargues y Luz (2000) y Pineda et al. (2002).
Se probaron las diferentes concentraciones del hongo B. bassiana, asperjando 30 ml de las suspensiones conidiales sobre lotes de 20 insectos, siguiendo la metodología descrita para la utilización de la torre de pulverización (Burgerjon 1956). La presión de aire se ajustó a 6 lbs para todos los experimentos. Los lotes de insectos se mantuvieron en cristalizadores plásticos y se incubaron en cámara climatizada a una temperatura 27°C y a una humedad relativa de 80% (Pineda et al. 2002).
Supervivencia de los insectos
El análisis de supervivencia, tiempo letal y tiempo de conidiación se realizaron teniendo en cuenta la prueba generalizada de Gehan y el test de Kaplan Meier (Lee 1980) con un alpha de 0,01 obtenido mediante la corrección de Bonferroni. Se utilizó el programa Statistica 98 Statsoft Inc., Tulsa, USA.
Conidiación
Los bioensayos se observaron diariamente durante 25 días y los insectos muertos se retiraron, se colocaron en una cámara húmeda (caja de Petri con papel de filtro Wattman más agua destilada estéril). Se registró el tiempo de aparición y conidiación del hongo sobre los insectos para cada una de las dosis asperjadas. Los insectos conidiados se colorearon con azul de lactofenol, en placas, para observar las diferentes estructuras morfológicas del hongo y verificar su identidad (Pineda et al. 2002).
Resultados
De los 320 insectos sometidos a la acción del hongo entomopatógeno a diferentes concentraciones, 219 (68,44%) murieron y presentaron micelio y conidiación de B. bassiana. El resto de los insectos, 101 (31,56%), no murieron y mudaron hacia el estado adulto. Los testigos (100%) sobrevivieron durante todo el tiempo del ensayo (25 días); el TL50 varió entre 5 y 25 días (Tabla 1). La concentración de 3x109 conidios/ml produjo una mortalidad del 100% de R. prolixus al noveno día con un tiempo letal medio de 5 días en comparación con la concentración de 3x105 conidios/ ml que eliminó menos del 50% en el mismo tiempo. Esto permitió observar diferencias en las curvas de supervivencia según la concentración (Fig. 1).
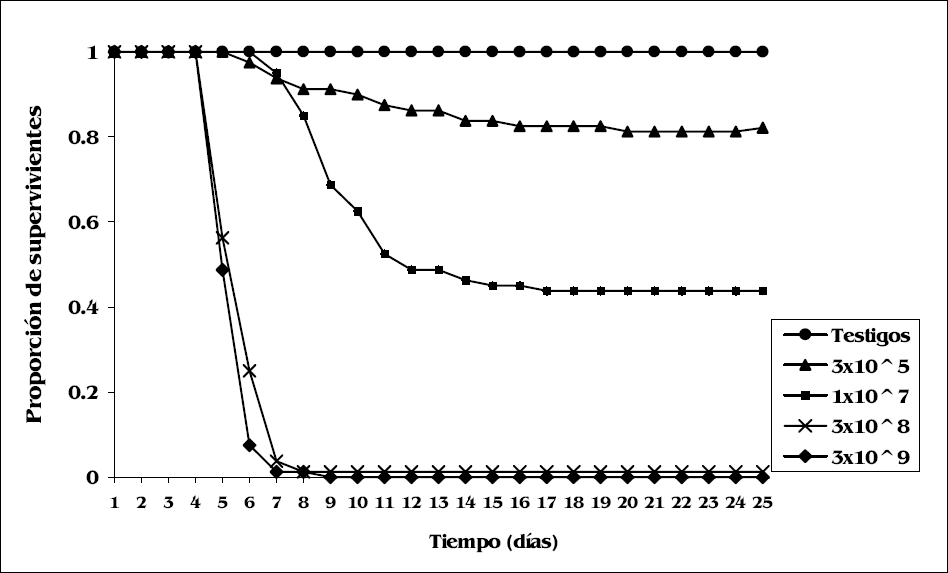
Supervivencia diaria de R. prolixus después del tratamiento con las dosis de B. bassiana UdeA13.
Sobrevivencia de R. prolixus a cuatro dosis de B. bassiana según el test estadístico de Kaplan-Meier
No se alcanzó el 50% de mortalidad.
Los análisis estadísticos mostraron diferencias significativas en mortalidad cuando se compararon los 4 tratamientos entre sí y el testigo. Las dosis 3x108 y 3x109 conidios/ ml no presentaron diferencias significativas entre sí (Z=-1,83, P= 0,067) (Tabla 2).
Comparación de las diferentes dosis evaluadas para supervivencia de R. prolixus y conidiación del hongo B. bassiana UdeA13 bajo el test estadístico de Kaplan- Meier con un alpha de 0,01
Dosis 0: control; 1: 3x105 conidios/ml; 2: 1x107 conidios/ml; 3: 3x108 conidios/ml; 4: 3x109 conidios/ml.
Los 219 R. prolixus que murieron, presentaron conidiación del hongo a partir del sexto día de su inoculación con las suspensiones conidiales. Se observó que a medida que aumenta la concentración se incrementa el porcentaje de insectos invadidos (Fig. 2).
Las concentraciones 3x105, 1x107, 3x108 y 3x109 conidios/ml presentaron diferencias significativas de conidiación (χβ=302, 63, P<0,00001), siendo mayor el número de insectos invadidos (100%) en la concentración 3x109 conidios/ml. Las concentraciones 3x109 у 3х108 conidios/ml no presentaron diferencias significativas entre sí (Z=-1,83, P=0,067) con tiempos medios de conidiación de 6 y 7 días, respectivamente (Tablas 2 y 3).
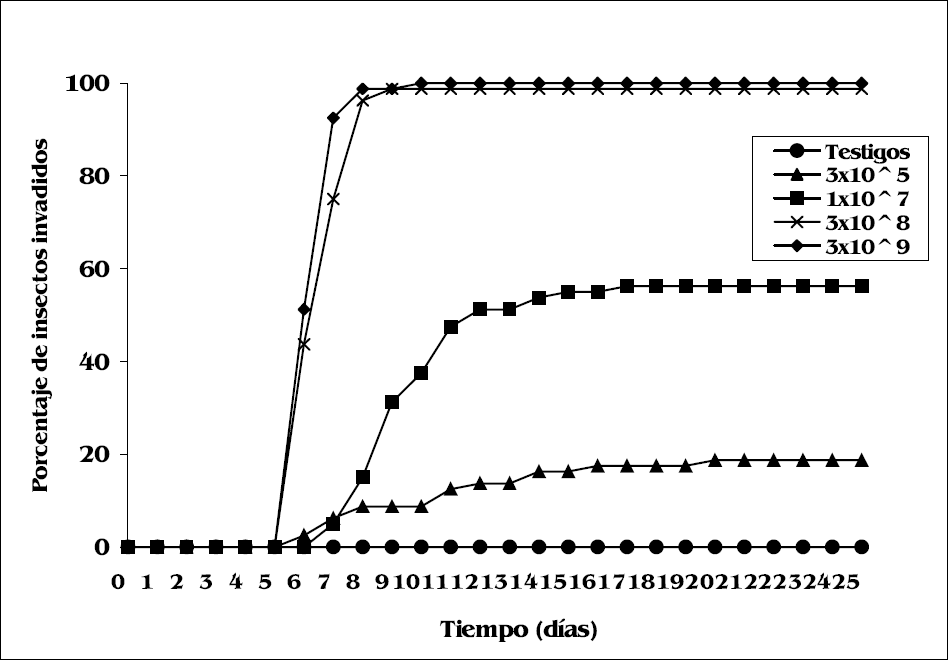
Cambio diario en el porcentaje de insectos con conidiación después del tratamiento con las diferentes concentraciones de B. bassiana UdeA13.
Tiempo de inicio de conidiación del hongo Beauveria bassiana UdeA13 sobre R. prolixus
No se alcanzó el 50% de insectos conidiados.
Discusión
La cepa B. bassiana UdeA13 utilizada en los ensayos causó una mortalidad alta en las ninfas de quinto estado de R. prolixus y presentó conidiación visible en los cadáveres.
La supervivencia de las ninfas de quinto estado de desarrollo de R. prolixus disminuyó a medida que se aumenta la concentración del inóculo del hongo B. bassiana UdeA13, resultados que concuerdan con trabajos realizados con hongos entomopatógenos sobre insectos plaga de importancia económica y algunos triatominos (Luz et al. 1998a; Lecuona et al. 2001; Gutiérrez 2003). Sin embargo, es importante resaltar que Pineda et al. (2002) mostraron que el hongo B. bassiana UdeA13 sobre R. ecuadoriensis de quinto estadio, bajo condiciones de humedad (80%) y temperatura (26±2°C) similares a las del presente estudio, produjo una mortalidad del 100% de insectos en las concentraciones de 3x105, 3x107 y 3x108 conidios/ml. Estas diferencias en la mortalidad de los insectos demuestran que especies de insectos relacionadas presentan grados de susceptibilidad diferentes a una misma cepa, como se menciona también en investigaciones realizadas por Arroyave (1995) y Luz et al. (1998b).
En estudios similares realizados por Romaña (1992), Luz y Fargues (1999), у Lecuona et al. (2001) con B. bassiana, el tiempo letal medio varió para Rhodnius prolixus y Triatoma infestans (Klug) entre 5,5 a 9,9 y 6,6 a 7,3 días, respectivamente. El tiempo letal medio varió dependiendo del estado ninfal y de las concentraciones del hongo. En la presente investigación los rangos variaron entre 5 y 11 días para todas las concentraciones, resultados que concuerdan con los estudios previos.
La conidiación se presentó de manera similar con todas las concentraciones de B. bassiana, iniciándose el día 1 después de haberse llevado los insectos a cámara húmeda, evolucionando de manera progresiva cada día hasta cubrir todo el cuerpo del insecto. La invasión se presentó principalmente en las áreas intersegmentales al igual que lo mencionan Tanada y Kaya (1993); iniciándose en las patas, proboscis, escutelo y finalmente en el tejido blando del abdomen, alcanzando el mayor grado de conidiación al día 5 (Fig. 3).
Existen pocas publicaciones relacionadas con cepas de hongos entomopatógenos aislados naturalmente de vectores triatominos en el campo. Parameswaran y Sankaran (1979) aislaron B. bassiana de Linshcosteus sp. (Luz et al. 2003) y Calle (2000) de R. pallescens. La ocurrencia natural de estos microorganismos es relevante para entender las relaciones del hongo con su hospedero en zonas infestadas y así poder seleccionar las cepas más patógenas y eficientes en el control de los triatominos.

A. Evolución de la conidiación del hongo B. bassiana sobre Rhodnius prolixus. a: Día 1. b: Día 2. c: Día 3. d: Día 4. e: Día 5. B. Zonas de conidiación en Rhodnius prolixus.
Conclusiones
Las ninfas de quinto estado de desarrollo de R. prolixus presentaron susceptibilidad al hongo B. bassiana (UdeA13), por lo cual se podría considerar como un buen agente de control para dicho vector.
El tiempo letal medio varió de acuerdo con las concentraciones de B. bassiana utilizadas mostrando que a medida que aumenta la concentración se presenta mayor mortalidad en menor tiempo.
La obtención del hongo B. bassiana (UdeA13) de los cadáveres de los insectos R. prolixus demostró su muerte por la acción del hongo, constituyéndose en fuente potencial de infección para otros insectos que habitan en el domicilio y peridomicilio, factor de gran importancia para un control natural en el campo.
Footnotes
Agradecimientos
Los autores agradecen a los profesores Abel Díaz, por la ayuda en el análisis estadístico, Fabio Pineda G., Jaime Calle O., Ana Isabel Gutiérrez G., Beatriz Cardona Y., Inés Elena Giraldo M. y Nadya Cardona B. por su apoyo técnico y logístico en la investigación. A la Corporación de Patologías Tropicales, al Instituto de Biología y al CODI de la Universidad de Antioquia por la financiación y el apoyo en esta investigación.