
Abstract
This research was conducted on the farm of the “Universidad Technological del Chocó”, municipality of Lloro, 90 elev. and 24°C. Surveys were conducted over twelve months by periodically sampling each of the plant species selected; insects were directly captured from plant structures. The diversity of hymenopterans between forest and agricultural components was compared; in addition, the trophic groups for hymenopterans from each parcel were established. Forty species were identified, distributed in 8 families, with 57,5% of the species belonging to the family Formicidae; followed by Apidae with 17,5%, Ichneumonidae with 10% and Vespidae with 5%; the remaining 10% belonged to Argidae, Braconidae, Mymaridae and Pompilidae. Borojoa patino had the highest diversity of Hymenoptera and Apeiba aspera the lowest. About 44,4% of the insect species were common to all four plant components. Among the phytophagous insects, nectar and pollen feeders were the most representative with 17,5% each. Among the entomophagous insects, predators predominated with 32,5%. Abundance of the insects was associated with the phenology of the plant species in the parcel. B. patinoi, Cedrella odorata and Inga spectabilis shared the same periods of regrowth and flowering between April and May, when the abundance of Hymenoptera and their natural enemies was highest. The most abundant species were Eulaena meriana, Lestrimelitta limao (first reports for Chocó) and Sericomyrmex sp. (first report of the genus for Chocó). One economically important species, Atta cephalotes, was identified because of its damage to B. patinoi fruits.
Introducción
El hombre del trópico, en su búsqueda por aumentar la producción agropecuaria, ha modificado su entorno, originando con ello ciertos cambios tales como, disminución en la biodiversidad por la deforestación, eliminación de la fauna autóctona, extinción de corrientes de agua, disminución de la pro ductividad agrícola, erosión de los suelos y migraciones indeseadas. Estas actividades antrópicas afectan la dinámica de los bosques naturales tropicales; razón por la cual hoy en día, para mitigar todo el daño que se ha venido haciendo a los ecosistemas naturales, los científicos han desarrollado y enriquecido un conjunto de prácticas y manejo de sistemas mixtos, donde de manera programada se mezclan especies leñosas de uso múltiple y se asocian en un mismo terreno de forma secuencia! o simultánea, cultivos agrícolas y animales, conocido este tipo de prácticas como sistemas agroforestales (Montanini 1992).
Los sistemas agroforestales surgen como una alternativa para las regiones tropicales de América Latina, que presentan limitaciones para el uso intensivo de la tierra con sistemas avanzados mecanizados, puesto que gran parte de la población está ubicada en zonas de ladera, las cuales presentan susceptibilidad a la erosión y degradación de los recursos si se emplean masivamente en sistemas de monocultivo o de ganadería. Situación similar ocurre en las tierras bajas tropicales, en donde domina una densa y exuberante cара boscosa, la cual es muy vulnerable y también presenta severas limitaciones para el establecimiento intensivo de sistemas tradicionales de producción agropecuaria. Los sistemas agroforestales tienen su origen en épocas muy remotas, pues antes de la llegada de los españoles a América, los indígenas desarrollaron este tipo de prácticas basadas en la tumba y quema del bosque y en el posterior establecimiento en una misma parcela de amplia variedad de cultivos anuales y árboles permanentes. Varias de estas asociaciones agroforestales prehispánicas han llegado hasta nuestros días siendo heredados por las comunidades indígenas actuales, que las conservan como su modo de producción (Vélez 1995).
En Colombia, estos sistemas productivos o agroforestales, son comunes en las selvas de la Amazonia, en áreas de bosque de la Orinoquia, en la región Pacífica, en algunas áreas boscosas interandinas y la Sierra Nevada de Santa Marta; actualmente, se vienen desarrollando en la costa Atlántica como alternativa para mitigar el impacto sobre los ecosistemas naturales de bosque seco tropical.
Entre los insectos, los himenópteros constituyen un grupo importante en la dinámica de estos agroecosistemas, pues con una historia evolutiva muy larga y la capacidad de adaptación a diferentes hábitats, se ubican como uno de los órdenes hiperdiversos en la clase hexápoda. En la actualidad se conocen entre 120.000 y 200.000 especies de himenópteros, pero recientes estimaciones apuntan a que su diversidad real es probablemente mucho mayor (del orden de 250.000 a 500.000 especies) rivalizando con la de coleópteros el puesto de grupo más diverso de insectos (Gastón 1991; La Salle y Gauld 1993; Gauld y Hanson 1995a; Gastón et al. 1996; Fernández 2000). Esta suposición se apoya en el hecho de que alguna de las superfamilias con mayor número de especies, integrada por los parasitoides de tamaño muy pequeño, están aún muy deficientemente estudiadas, especialmente en las zonas tropicales donde se cree que su diversidad potencial es muy alta (Fernández 2000).
Los himenópteros no sólo se destacan por su gran riqueza de especies, sino también por su extraordinaria diversidad de modos de vida que van desde la fitofagia a la depredación y el parasitismo o la inducción de agallas y desde el comportamiento solitario hasta el mutualismo o la formación de sociedades complejas. En cuanto al aspecto económico su importancia para el hombre supera la de cualquier otro grupo de insectos. Algunas facetas de interés en este sentido, son la polinización de plantas cultivadas, el control biológico por parte de los himenópteros parasitoides de plagas agrícolas y forestales y la producción de bienes comerciales como la miel y la cera (LaSalle y Gauld 1993), por lo cual, este grupo es fundamental en la dinámica de estos sistemas productivos del pacífico colombiano. Los objetivos del estudio consistieron en identificar la fauna de himenópteros asociada a la parcela agroforestal, conocer las relaciones entre ellos y el componente vegetal y establecer la variación mensual de los himenópteros en la parcela.
Materiales y Métodos
La granja de la Universidad Tecnológica del Chocó se encuentra ubicada en el municipio de Lloró-Chocó (5°30,52′57″N, 76°33.33′15″O); sobre los 90 msnm, con una humedad relativa del 86%, una precipitación anual de 10.851 mm y una temperatura de 24°C; se ubica en la zona de vida de bosque pluvial tropical “bp-T” (Espinal 1977); es considerada como la región más lluviosa de la porción central de la llanura del Pacífico (Hans 1988).
La parcela está compuesta por 4 especies vegetales con un total de 202 individuos, distribuidos de la siguiente manera: 77,77% de Borojoa patinoi Cuatrecasas, 6,90% de Cedrela odorata L., 6,13% de Apeiba aspera Aubl y 9,20% de Inga spectabilis Willd, con dos estratos bien diferenciados, uno inferior dominado por B. patinoi y uno superior ocupado por las 3 especies de estructura arbórea. Por tales características, esta asociación corresponde a un sistema simultáneo, de tecnología agrosilvícola y el tipo de arreglo presente es de árboles permanentes sobre cultivos permanentes, con una extensión de 7.946,35 mβ.
Una vez realizado el inventario de las especies vegetales en la asociación, se seleccionaron al azar 14 individuos y se marcaron. Estos árboles se muestrearon cada 15 días durante los meses de noviembre de 1988 a octubre de 1999. Las capturas de los himenópteros se llevaron a cabo en cada una de las estructuras, tanto vegetativas como reproductivas de las plantas, a través de recorridos y colección, mediante método manual con el fin de establecer la relación planta-insecto, predador-presa, parasitoide-hospedero. El muestreo se complementó con anotaciones sobre el comportamiento de algunos himenópteros.
Las especies de himenópteros se separaron de acuerdo con la especie vegetal, el individuo y el muestreo realizado, así como con los datos de captura, con el fin de estandarizar los datos para su análisis estadístico posterior. Los individuos adultos se conservaron en alcohol al 70% y los estados inmaduros se colocaron en cajas de Petri sobre el sustrato en el cual fueron capturados, con el fin de obtener los adultos para su identificación. Éstos se transportaron al laboratorio de zoología de la Universidad Tecnológica del Chocó, donde se les realizó el seguimiento.
El montaje y rotulación del material se realizó de acuerdo con Borror et al. (1981) у el método usado en el Museo Entomológico “Francisco Luis Gallego” de la Universidad Nacional de Colombia- sede Medellín y el Museo de Historia Natural Colección de Entomología de la Universidad Tecnológica del Chocó “Diego Luis Córdoba”.
Para la identificación taxonómica de los himenópteros se utilizaron claves desarrolladas por los diferentes especialistas de cada grupo como Fernández (1991, 1993, 2002), Gauld y Hanson (1995a, 1995b, 1995c, 1995d), Goulet y Huber (1993), Hölldobler y Wilson (1990), Hanson y Gauld (1995), Mackay (1993a), Mackay y Mackay (1986, 1989), Masner y Dessart (1967), Palacio (1999), Richards (1977), Sharkey y Wahl (1992), Serna (1999). Así mismo, se contó con la colaboración de especialistas nacionales e internacionales como Francisco Javier Serna, Alan Henry Smith, Paul Hanson y Carolina Godoy.
Para el análisis estadístico se consideró la frecuencia de captura de los himenópteros y no el número absoluto de individuos colectados, debido a que este número varía según la especie (Hölldobler y Wilson 1990).
Con el fin de comparar la diversidad de himenópteros en cada una de las especies vegetales en la parcela y entre los individuos de una misma especie se utilizó el índice de diversidad de Shanon-Weaver (Ludwing y Reinolds 1988).
El tratamiento de la medida de diversidad se realizó mediante un análisis de varianza, donde la variable de respuesta está en los diferentes componentes vegetales presentes en la parcela objeto de estudio. Se utilizó una prueba de comparación múltiple Tukey (a=0,05), para observar si hay diferencias significativas entre las especies de plantas asociadas (Steel y Torrie 1980).
Para conocer la similitud de los himenópteros asociados a los componentes, se utilizó el índice de Jaccard, el cual está basado en la presencia de especies, representando éste el valor porcentual de las ocurrencias simultáneas respecto al total de especies y su oscilación respecto a un límite superior (1) e inferior (0), a partir del cual se obtuvo una matriz y posteriormente un dendrograma de afinidad (Ramírez 1999).
En cada mes se tomó la frecuencia de ocurrencia, es decir, la presencia de las especies de himenópteros por muestreo en cada una de las especies e individuos de las plantas asociadas, obteniendo un registro de 12 muestras correspondientes al 100% de la frecuencia. Para determinar si existían diferencias entre los meses se realizó una prueba de Chi Cuadrado.
Resultados
Análisis de la fauna de himenópteros en la parcela agroforestal
Se identificaron 40 especies, distribuidas en 8 familias, así: familia Formicidae representada por el 57,5%, seguida de Apidae con 17,5%, Ichneumonidae con 10%, Vespidae con 5% y el 10% restante compartido por las familias Argidae, Braconidae, Mymaridae y Pompilidae con una especie cada uno (Tabla 1).
Himenópteros asociados a la parcela agroforestal
En la familia Formicidae, se identificaron 23 especies, agrupadas en 6 subfamilias y 15 géneros. El 30,43% de los géneros pertenece a Myrmicinae, seguida de Ponerinae con 26,08% y el 43,49% restante compartido por Ectitoninae, Dolichoderinae, Formicinae y Pseudomyrmecinae (Fig. 1).
La subfamilia Myrmicinae presentó el número mayor de géneros con un total de 6, Acromyrmex, Atta, Cephalotes, Crematogaster, Solenopsis y Sericomyrmex, siendo Atta el género con la mayor cantidad de especies (28,57%), mientras que los demás estuvieron representados por una sola especie. De la subfamilia Ponerinae, la segunda mejor representada en la parcela, se encontraron 4 géneros: Ectatommа, Pachycondyla, Odontomachus y Paraponera, siendo Ectatomma y Pachycondyla los de la mayor riqueza con 33,33% de especies cada uno.
En la familia Apidae se identificaron 7 especies de 2 subfamilias, ubicándose como la familia con el segundo porcentaje de representación en la parcela. El 57,14% de las especies pertenecen a la tribu Meliponini y el 42,86% a Euglossini; en los meliponinos el género Trigona presentó tres especies y Lestrimelitta una especie, mientras en los euglosinos, Eulaema dos especies y Euglossa una.
En la familia Ichneumonidae, la subfamilia Orthocentrinae con 50% de las especies fue la más representativa, mientras Cryptinae e Ichneumoninae fueron las menos abundantes y poco diversas.
La familia Vespidae estuvo representada sólo por la subfamilia Polistinae con 2 géneros, Polybia y Polistes con una especie cada uno; las demás familias Argidae, Braconidae, Mymaridae y Pompilidae con una especie cada una.
El género Sericomyrmex Mayr, y las especies Eulaema meriana Oliver y Lestrimelitta limao Smith son nuevos registros para el departamento del Chocó. Las especies Camponotus sp., Dolichoderus sp., Euglossa sp., Eulaema cingulata F. y Eciton hamatum F. son nuevos registros para la cuenca fluvial del Atrato (Fernández 1995).
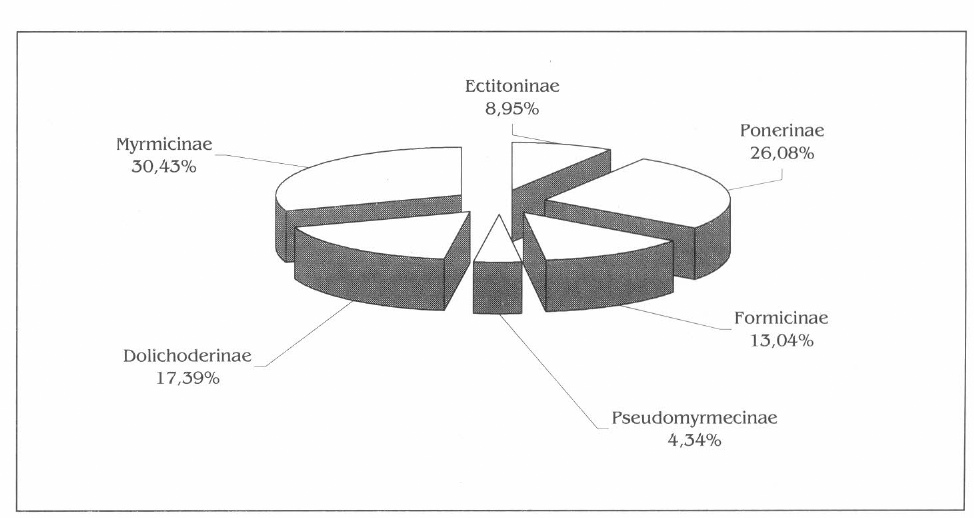
Composición de la familia Formicidae en la parcela agroforestal.
Comparación de los himenópteros asociados a las especies vegetales en la parcela agroforestal
Borojoa patinoi presentó la mayor diversidad y riqueza de insectos en la parcela, seguida por Inga spectabilis y la menor la tuvo Apeiba aspera. Habiéndose calculado el índice de diversidad de Shannon-Weaver y una vez confirmado la distribución normal de la diversidad y la homogeneidad de las variables, se realizó una ANOVA de doble vía (Aldana y Chacón 1999), donde se encontró diferencia significativa en las especies vegetales de la parcela (F= 0,05, p = 0,95). El análisis de comparación múltiple (Prueba de Tukey), indicó que sólo las especies Inga spectabilis y Cedrela odorata no muestran entre sí diferencia significativa (Tabla 2).
Diversidad de Shannon-Weaver para los himenópteros asociados al componente vegetal en la asociación agroforestal
En cuanto a la afinidad de los himenópteros asociados a las especies vegetales, se encontró que C. odorata e I. spectabilis comparten el 77% de los himenópteros, seguida por A. aspera y B. patinoi con 65,8%. La afinidad más baja se presentó entre las especies A. aspera y C. odorata con 33,33% e I. spectabilis y A. aspera con 34,3%; tan sólo el 44,46% de los himenópteros se encuentran asociados a los cuatro componentes vegetales asociados en la parcela (Fig. 2).
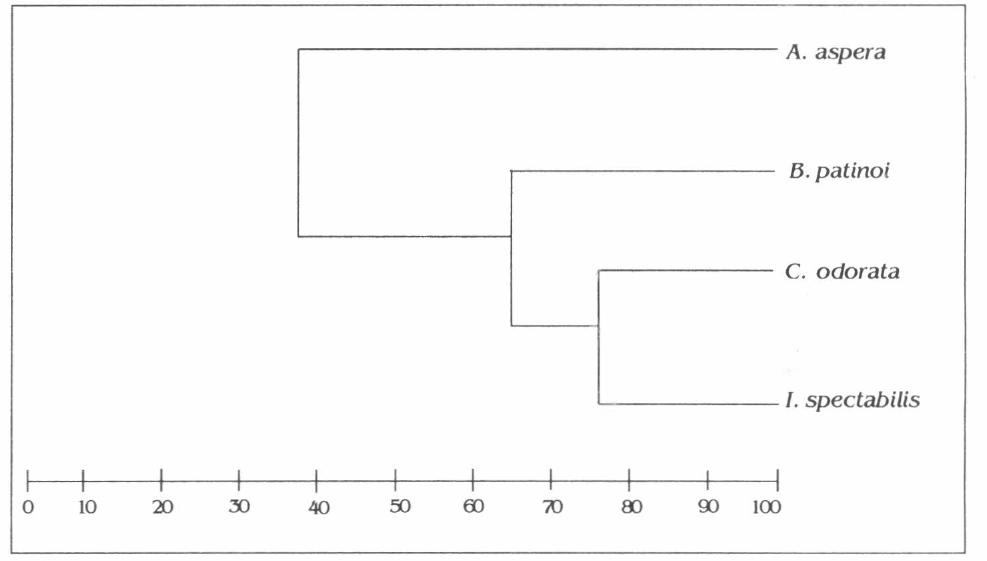
Dendrograma de afinidad de los himenópteros en las especies vegetales presentes en la asociación.
En relación con la composición trófica de la fauna de himenópteros, los entomófagos y fitófagos tuvieron igual representividad, ambos con 45%. En los entomófagos, los depredadores fue el grupo mejor representado, seguido por los parasitoides; mientras que en los fitófagos, los lamedores de néctar y palinófagos fueron los más abundantes, seguidos por los cortadores de hojas. Otros grupos de importancia en la parcela fueron los saprófagos, micófagos, necrófagos y parásitos (Fig. 3).
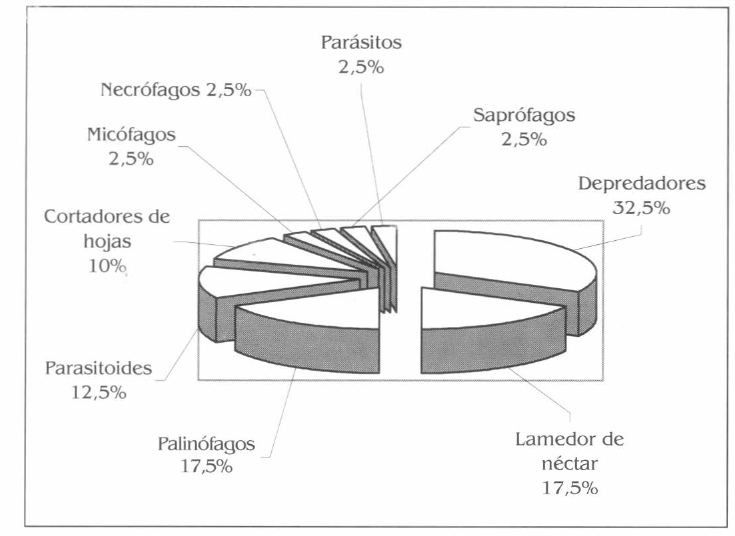
Diversidad trófica de los himenópteros asociados a la parcela agroforestal.
Variación de la fauna de himenópteros en los meses de muestreo en la parcela agroforestal
La frecuencia de himenópteros en la parcela agroforestal varió en cada una de las especies vegetales asociadas. Se presentaron algunas variaciones en la ocurrencia de los himenópteros, con diferencia significativa en la especie C. odorata X2=30,38 (DF=11; X0,992>P(X2=30,38)>X2=0,90); y la no existencia de tal diferencia en los otros componentes; es decir, I. spectabilis X2=15,07 (DF=11; X0,992<P(X2=15,07)<X2=0,90, A. aspera X2=4,17 (DF=11; X0,992<P(X2=4,17)<X2=0,90 y B. patinoi X2=21,64 (DF=11; X0,992<P(X2=21,64)<X2=0,975).
La mayor frecuencia de himenópteros corresponde a los meses de marzo a junio, época en la cual las especies vegetales muestran rebrotes de hojas, producción de frutos y flores, en especial C. odorata, A. aspera y B. patinoi, mientras que I. spectabilis aunque no presenta floración ni fructificación coincide con el rebrote foliar; siendo estos meses los de mayor precipitación. En este período la mayor frecuencia de los himenópteros está representada por las especies de hormigas como Atta colombica, Dolichoderus complejo bispinosus, Odontomachus sp., Pachycondyla sp., Pachycondyla sp., Paraponera clavata, Solenopsis sp., Sericomyrmex sp., Azteca sp., de igual forma las abejas Eulaema cingulata, Eulaema meriana, Euglossa sp, Trigona sp., Trigona sp., Trigona sp. y las avispas Polybia rejecta y Polistes carnifex carnifex y parasitoides de las familias Ichneumonidae, Mymaridae y Braconidae (Fig. 4a).
En B. patinoi, las especies más frecuentes fueron Dolichoderus complejo bispinosus, Paraponera clavata, comunes en todos los mes de muestreo; existen especies exclusivas de éste como Eulaema cingulata, Eulaema meriana, Euglossa sp.1, Trigona sp., Trigona sp., Trigona sp., y Atta cephalotes, Camponotus sp., Dolichoderus sp., Dolichoderus sp., Sericomyrmex sp. y otros como Polybia rejecta y Polistes carnifex carnifex y una especie de icneumónidos y bracónidos, respectivamente (Fig. 4b).
En I. spectabilis, las especies más frecuentes fueron Cephalotes atratus, Dolichoderus complejo bispinosus, colectándose en todos los muestreos, poseyendo a su vez especies muy exclusivas como Cephalotes atratus y Pepsis sp. (Fig. 4c).
En C. odorata, las especies más frecuentes fueron Ectatomma tuberculatum, Paraponera clavata, Dolichoderus complejo bispinosus, las cuales se estuvieron durante todo el periodo de estudio (Fig. 4d).
En A. aspera, de las 8 especies de himenópteros asociadas, la mayor frecuencia la presentó Azteca sp., la cual fue común durante todo el muestreo (Fig. 4e).
De las especies de himenópteros asociados a la parcela agroforestal, la especie más dañina fue Atta cephalotes, por cortar el epicarpio y el mesocarpio de B. patinoi.
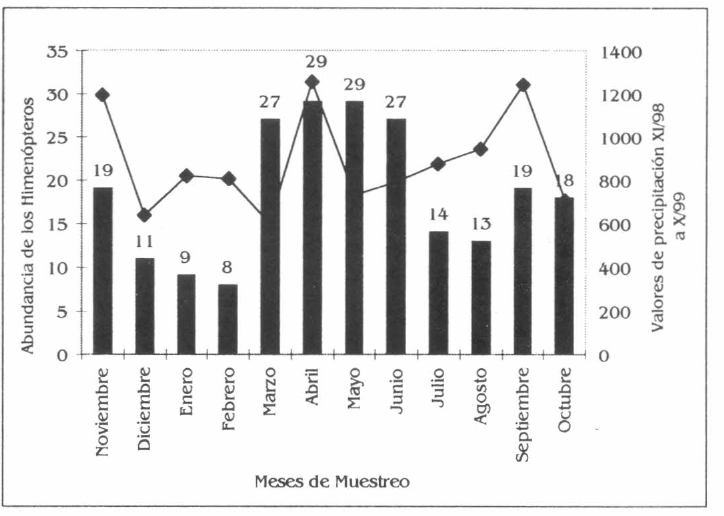
Frecuencia de los himenópteros en la parcela agroforestal de Borojoa patinoi C., Cedrela odorata L., Apeiba aspera A. e Inga spectabilis W.
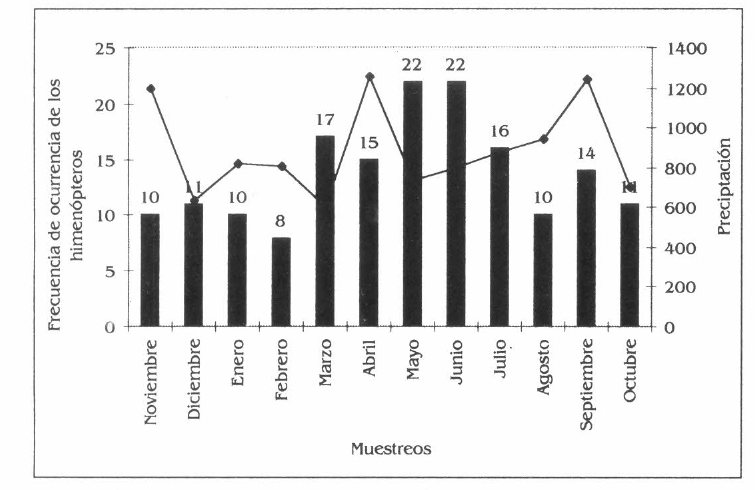
Frecuencia de los himenópteros en Borojoa patinoi.
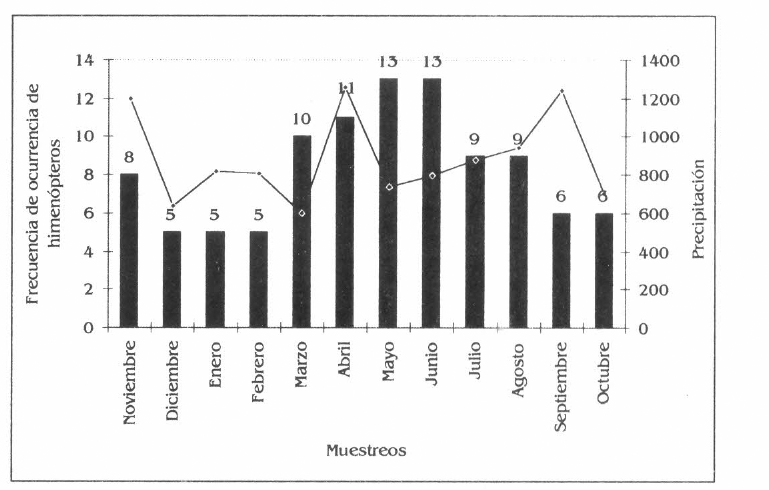
Frecuencia de los himenópteros en Inga spectabilis.
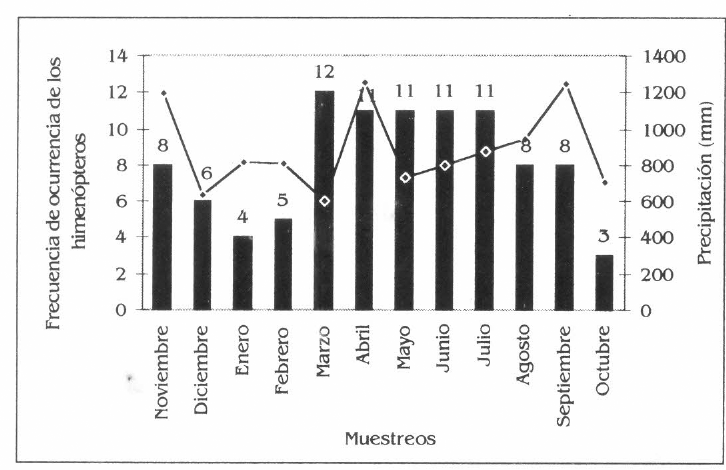
Frecuencia de himenópteros en Cedrela odorata.
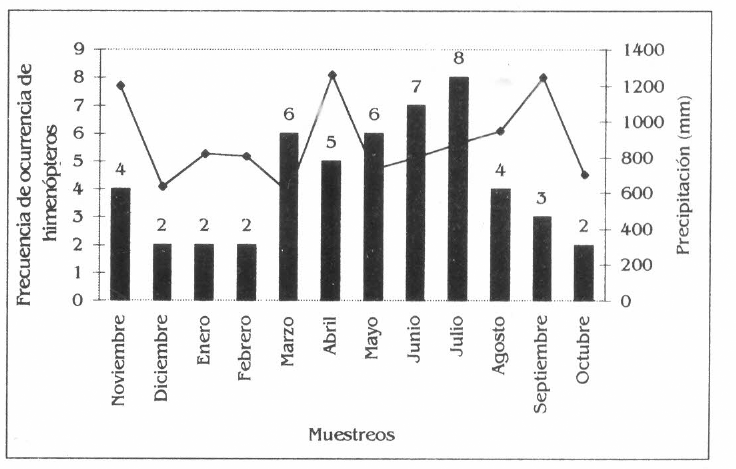
Frecuencia de himenópteros en Apeiba aspera.
Discusión
De los himenópteros asociados a la parcela agroforestal, las hormigas (Formicidae) fueron las más abundantes y con mayor número de especies. Se ha estimado que en los trópicos estos organismos representan la mitad o una tercera parte de la biomasa de insectos (Way y Khoo 1992). Debido a esta abundancia, estabilidad en el espacio y en el tiempo, sumado a sus hábitos alimentarios, generalmente omnívoros, las hormigas tienen una importante influencia en muchos hábitats (Hölldobler y Wilson 1990); es por ello la importancia en la dinámica de los agroecosistemas tropicales del pacífico Colombiano.
Se colectaron 6 de las 8 subfamilias de hormigas registradas para el trópico, esto se debe a la presencia de una gran diversidad de nichos, originados por la estratificación en los policultivos de estructura arbórea presente en esta asociación, siendo similar en muchos aspectos ecológicos a los ecosistemas naturales.
Otra familia de importancia en la parcela fue Apidae, la cual estuvo representada por 7 especies, siendo algunas de ellas muy exclusivas de algunos componentes vegetales en la parcela.
La abundancia de Ichneumonidae en la parcela obedeció principalmente a la abundante presencia de hospederos para sus estados inmaduros, los cuales se encontraban asociados a partes reproductivas y vegetativas de los componentes vegetales de la parcela.
B. patinoi, a pesar de ser el único componente agrícola en la parcela, presentó la mayor diversidad y riqueza de himenópteros, debido a la presencia de estas especies en los estratos medio e inferior de la parcela, los cuales presentan variaciones microclimáticas que favorecen una mayor presencia de nichos y por ende la mayor abundancia de insectos.
Otro aspecto fundamental fue la presencia en el tiempo y el espacio de los órganos tanto reproductivos como vegetativos de esta planta, lo que contribuye a mejorar las actividades antes mencionadas, así la exclusividad de algunos himenópteros como: Euglossa sp., Eulaema cingulata y Eulaema mariana, los cuales se encuentran asociados a la flor macho y hembra de Borojoa patinoi, como también las especies pertenecientes al género Trigona asociadas a receptáculos y cáliz de la flor macho de esta misma planta.
Por otro lado, la baja diversidad de A. aspera obedece a la poca apetencia de los fitófagos por esta especie, salvo Atta colombica que consume sus hojas durante el rebrote, no encontrándose otro tipo de relación fitófagica con otros himenópteros, pero, sin embargo, sirve como hospedero de otros insectos que son depredados y parasitados por algunas especies de la himenópterofauna, aunque poco abundante y diversa.
La estratificación vertical de la parcela permitió identificar la presencia de 3 estratos, los que a su vez, dejaron que los himenópteros se distribuyeran en ellos, dada la variabilidad de microclimas, aproximación física y mayor cantidad de nichos. Por estas razones, la similitud en la parcela es baja, siendo más alta en las especies de porte arbóreo que comparten el estrato superior de la parcela como C. odorata e I. spectabilis, que no presentan diferencia significativa en cuanto a su diversidad. De otro lado, la menor afinidad de A. aspera con otros componentes vegetales asociados se debe posiblemente a que ella se encontró ubicada en la parte noreste de la parcela actuando como cortina rompeviento, sitio en el que tuvo una mayor aproximación física con el componente agrícola B. patinoi, situación que puede explicar su afinidad alta con esta especie y su afinidad baja por la poca dispersión de manera fortuita en el área con relación a las otras especies forestales, a pesar de ser ella de porte arbóreo.
En la diversidad trófica de la himenópteros en la parcela, los entomófagos fueron los más representativos sobresaliendo los depredadores conformados especialmente por hormigas pertenecientes a los géneros Pseudomyrmex, Pachycondyla, Odontomachus, Ectatomma, Eciton y Paraponera y las avispas Polybia, Polistes y Pepsis; en el caso de los primeros, es decir, las hormigas su depredación se dio sobre larvas de Pyralidae (Lepidoptera) y Curculionidae (Coleoptera), que se encontraban barrenado el fuste del I. spectabilis. Otra presa la constituyó Nasitutermes sp. (Isoptera: Termitidae), la cual construye su nido en la corteza de A. aspera; el segundo caso corresponde a las avispas, pues su actividad entomofágica se dio sobre larvas de Pyralidae, comedoras de hojas de B. patinoi y estados inmaduros de Cicadellidae (Heteroptera) en fuste de A. aspera y arañas en bifurcaciones del componente arbóreo.
En los parasitoides se encontraron especies de bracónidos, icneumónidos y mimáridos parasitando las larvas de Noctuidae encontradas en galerías de Nasitutermes sp., así como de algunos Pyralidae barrenadores de frutos de A. aspera.
En el caso de los fitófagos, el grupo de los lamedores de néctar y palinófagos fueron los más representativos siendo abundantes para el primer caso, hormigas del género Camponotus, Dolichoderus, Solenopsis debido a la constante presencia en diferentes estados de desarrollo del fruto de B. patinoi, el cual concentra gran cantidad de néctar en su parte inferior (el receptáculo), por espacio de 7 meses aproximadamente; el segundo grupo estuvo conformado por los géneros Eulaema, Euglossa y Trigona los cuales se observaron alimentándose en flores masculinas y femeninas de B. patinoi.
Las especies cortadoras de hojas tuvieron su mayor representación en el género Atta con dos especies, mientras que Acromyrmex y Sericomyrmex tuvieron una especie cada una. Se encontraron preferencias por las estructuras vegetativas y reproductivas de C. odorata, A. aspera e I. spectabilis, así Atta colombica y A. cephalotes forrajearon los frutos de B. patinoi y hojas en diferentes estados de desarrollo en algunas plantas aisladas, las cuales coincidieron con poco sombrío por parte del componente arbóreo. En el caso de Sericomyrmex su forrajeo fue exclusivo en pétalos y cáliz de B. patinoi.
Otro grupo importante lo constituyen los Arguidos, Azteca sp. y Dolichoderus complejo bispinosus, debido a sus hábitos saprófagos, micófagos y necrófagos, respectivamente. Ellos contribuyen en la dinámica de estos agroecosistemas, principalmente en el ciclaje de nutrientes a través de la descomposición de animales y partes vegetativas y reproductivas de las especies vegetales. Este proceso se puede ver favorecido por las condiciones de alta humedad, precipitación y temperatura reinantes en la parcela agroforestal, característica biofísica importante de los sistemas agroforestales (Montanini 1992).
La diferencia significativa que presentó C. odorata, en cuanto a la frecuencia de especies de himenópteros, se debe principalmente a que esta especie, por su carácter caducifolio, permanece seis de los doce meses del año sin follaje, presentando en esta etapa fenológica muy poca presencia de los insectos y aumentando de una manera considerable en los meses donde existen los periodos de rebrote, floración, fructificación y dispersión de semillas. Estas actividades fisiológicas de las plantas se dieron desde abril a septiembre, coincidiendo con las mayores precipitaciones registradas durante el muestreo.
Para las otras especies vegetales, aunque sus etapas fenológicas se presentaron desde finales de marzo hasta octubre, los cambios en dichas actividades no fueron tan notorios como en el caso de C. odorata, ya que las estructuras vegetativas y reproductivas de estas especies permanecieron en el tiempo y los cambios fueron muy paulatinos, razón por la cual no existieron diferencias entre los meses de muestreo, en cuanto a la frecuencia de himenópteros, ya que había disponibilidad de alimento durante el año.
La ocurrencia de insectos fitófagos en la parcela mostró una fuerte relación con aquellos de hábitos entomófagos, sobre todo con los depredadores. De esta manera, la especie Nasitutermes sp. era depredada por las especies de hormiga Ectatomma ruidum, Pachycondyla sp., Pachycondyla sp. y Odontomachus sp.; mientras que las larvas de Pyralidae, barrenadores del fuste de I. spectabilis, eran depredadas por la avispa Polistes carnifex carnifex, y las hormigas Pachycondyla sp. y Odontomachus sp., siendo esta época la de mayor precipitación.
De otro lado, la frecuencia de ocurrencia de las especies depredadoras Eciton burchelli y Eciton hamatum estuvo relacionada con la presencia de estados inmaduros en colonias de otras hormigas como Atta cephalotes, Camponotus sp., Solenopsis sp. y Odontomachus sp.
Por otra parte, entre los meses de abril y mayo, la especie I. spectabilis presentaba rebrotes de hojas, las cuales eran forrajeadas por Atta colombica y A. cephalotes. Durante este evento la especie Paraponera clavata se observó sacrificando las obreras que forrajeaban este árbol, asumido esto como un comportamiento defensivo y no como una depredación propiamente dicha, ya que no eran consumidas por éstas.
Otros himenópteros como las abejas, cuya frecuencia fue mayor en los meses de abril a julio, obedece a que en esta época la especie B. patinoi presenta tanto flores masculinas como femeninas, a las cuales se encuentran asociadas estas especies por encontrar mayor cantidad de néctar y polen en las estructuras reproductivas.
Conclusiones
La riqueza y diversidad de los himenópteros asociados a la parcela da soporte a una de las características más importantes de los sistemas agroforestales, como es la alta diversidad organísmica que albergan. Debido a esto presentan gran similitud con los ecosistemas naturales, por ello hoy en día se intensifica su aplicabilidad en el trópico húmedo como una alternativa para mitigar el impacto ecológico ocasionado por los monocultivos en estas zonas.
El conocimiento de la diversidad trófica de los himenópteros como estudio básico en sistemas agroforestales es fundamental para el manejo adecuado de aquellos grupos de insectos fitófagos potencialmente importantes desde el punto de vista económico.
La afinidad de los himenópteros entre las especies vegetales de la parcela agroforestal, está dada por estratificación de los policultivos de estructura arbórea y la distribución horizontal del componente agrícola en la misma.
La presencia de los himenópteros en la parcela agroforestal está relacionada con los periodos fenológicos propios de cada especie vegetal, y no con la precipitación. Por tal razón, y debido a su carácter caducifolio, la especie C. odorata fue la única que presentó una diferencia estadísticamente significativa en la frecuencia de himenópteros durante el muestreo.
Footnotes
Agradecimientos
Los autores agradecen a Francisco Javier Serna por la colaboración en la identificación de las hormigas, Allan Henrry Smith en la identificación de las abejas, Paul Hanson en la identificación de Vespidae y Pompilidae, Carolina Godoy en la identificación de Cidadellidae, Jhon A. Quiroz en la identificación de algunos grupos de insectos. A Oscar E. Ortega por la asesoría en la parte ecológica. A los profesores Rodrigo Escobar Durán y Alicia Ríos H. de la Universidad Tecnológica del Chocó por su apoyo durante el desarrollo de la investigación. A Julia del C. Palacios, Nasser Cuesta, Rosalba Bonilla y María Lucila Rentería R. por su colaboración en el campo. Al grupo de Zoología-Universidad Tecnológica del Chocó MEFLG Universidad Nacional de Colombia-Sede Medellín por el apoyo logístico en el desarrollo de la investigación.