
Abstract
This research elucidated general aspects of the habits of oviposition and behavior of the Guatemalan potato tuber moth Tecia solanivora (Povolny) in stored potato. These aspects included determination of the pest's damage capacity, preferred oviposition sites in stows as well as the tuber, and influence of the state of cleanliness of the stored tubers on oviposition habits. The study was conducted at the Tibaitatá Research Center, in the IPM program of Corpoica and was carried out under controlled conditions of temperature, humidity and light. The experimental unit consisted of a wooden cage with muslin walls, in which a tray was introduced with 100 tubers, of 100 g average weight each. In the determination of the damage capacity of the pest it was found that 10 pairs of the Guatemalan potato tuber moth released on 100 tubers, produce over 35% of damage. Females T. solanivora prefer to oviposit mainly under the soil adhered to the tubers, around the area of the buds of the tuber and in the tubers located in the most protected places of the potato stow. As for the state of cleanliness of the tubers, it was found that the female prefers to oviposit in dirty tubers compared to clean tubers, therefore the total number of eggs placed on the same resource is higher in dirty potatoes.
Introducción
El cultivo de la papa es la actividad agrícola más sobresaliente en la zona fría en Colombia por diferentes razones: En lo económico se muestra que en 1996 el cultivo participó con cerca del 5,9% del total del valor de la producción agrícola, con 173.702 hectáreas sembradas, y para el año 2000 fueron sembradas unas 170.719 hectáreas (Fedepapa 2001). Es el cultivo de mayor demanda de plaguicidas químicos y el segundo después del café, en uso de fertilizantes, también es el producto agrícola que genera más transporte terrestre en Colombia. En lo social se encuentra que alrededor de 95.000 familias dependen directamente del cultivo, equivalente a unas 500.000 personas; al mismo tiempo, transportadores, comerciantes, lavadores, seleccionadores, procesadores, productores y otros agentes que intervienen en la cadena de la papa, dependen de manera indirecta de este cultivo (Porras 1999).
Una de las plagas de mayor importancia económica en el cultivo de la papa y que afecta de forma directa la calidad del tu bérculo es “la polilla guatemalteca de la papa”, Tecia solanivora (Povolny). Esta plaga es originaria de Centro América y fue introducida a Sur América en una importación de semilla hecha por Venezuela desde Costa Rica en 1983. Dicha semilla fue distribuida a los agricultores de la zona papera del Táchira, de donde fue introducida a Colombia (Araque y García 1999).
La polilla se estableció en la zona papera de Norte de Santander registrándose por primera vez en 1985 en el municipio de Chitagá (Soriano 1999). De allí inició su dispersión a todas las zonas productoras, conociéndose por primera vez en Boyacá y Cundinamarca en 1991, a Antioquia se propagó durante el año 1994 y en 1996 llegó a Nariño y a Ecuador (Espitia 1999).
La polilla guatemalteca ha cobrado gran importancia debido a la severidad de su ataque, debido a que ocasiona grandes pérdidas económicas en las áreas que invade. Así por ejemplo, en Norte de Santander las pérdidas en campo y almacenamiento superaron el 50% en 1986. En Boyacá y Cundinamarca las pérdidas iniciales alcanzaron hasta un 80% en campo en el año 1994. En el municipio de Ventaquemada en la vereda el Hato, la plaga arrasó con lotes en los cuales hasta el 90% de la papa de la cosecha de mitaca fue atacada. En 1996, se calculó que 540 toneladas de papa fueron consumidas por la plaga en sitios de almacenamiento y más de 2.000 toneladas en los lotes del cultivo para el altiplano cundiboyacense (Espitia 1999).
La producción de papa en Colombia reúne una serie de prácticas de cultivo que difieren de una región a otra en el país, siendo la poscosecha, o almacenamiento, una de las prácticas en donde existe probablemente la mayor variación a nivel nacional. La forma tradicional de almacenamiento en el altiplano cundiboyacense es en bultos, a granel o en cajas de madera guardadas en cuartos oscuros, pequeñas bodegas o en ramadas protegidas de la lluvia (Porras 1999).
Estos lugares generalmente son inapropiados para el almacenamiento porque las condiciones de humedad, temperatura y luminosidad favorecen la llegada y permanencia de la plaga en el sitio de almacenamiento. Esto, a su vez, genera una amplia gama de problemas que se deben abordar teniendo en cuenta el factor humano de cada región (Porras 1999).
Ante la importancia adquirida por la polilla guatemalteca como plaga del cultivo de la papa y el desconocimiento de sus hábitos y comportamiento en condiciones de almacenamiento, se realizó este trabajo en el Centro de Investigación “Tibaitatá” de Corpoica, el objetivo general fue conocer los hábitos y comportamiento de la polilla guatemalteca en condiciones de almacenamiento. Y los objetivos específicos consistieron en determinar la capacidad de daño de T. solanivora en papa almacenada y conocer los hábitos de oviposición de la plaga en esas condiciones.
Materiales y Métodos
El presente trabajo de investigación se llevó a cabo en el laboratorio de entomología del Programa Nacional de Manejo Integrado de plagas - MIP, en el Centro de Investigación “Tibaitatá” de Corpoica, localizado en el municipio de Mosquera (Cundinamarca), con una altitud de 2.550 msnm. Los experimentos se realizaron en condiciones ambientales controladas, con temperatura promedio de 18°C ± 3, humedad relativa del 60% ± 5. El material biológico de la plaga se obtuvo de las unidades de cría establecidas en el laboratorio de entomología, en donde se producen las cantidades que se requieren del insecto mediante la siguiente metodología general.
La cría de T. solanivora se inicia colocando posturas de esta especie sobre tubérculos de papa sanos dentro de cámaras de pupamiento y desarrollo larval (Fig. 1a) que son cubetas plásticas semitransparentes de 33 cm de largo x 26 cm de ancho x 10 cm de alto, cubiertas con su tapa a la cual se le realiza un orificio de 20 cm x 12 cm que se cubre con muselina, para facilitar el intercambio de aire. Dentro de estas cámaras transcurren los ínstares larvales de la plaga y el estado de pupa. Aproximadamente 20 días después de colocadas las posturas, comienza la formación de pupas dentro de las cámaras y pasados 48 días desde la colocación de los huevos hasta la emergencia de los primeros adultos.
Estos adultos son colectados y colocados en cámaras de oviposición (Fig. 1b), que son frascos de vidrio de 250 ml de capacidad tapados con muselina, donde son alimentados con una solución de miel al 10%, impregnada en un algodón. Las hembras ovipositan toda su carga de huevos en el transcurso de su vida sobre trozos de papel toalla que se colocan debajo de las cámaras. Las posturas se recogen diariamente y son utilizadas para experimentación o para continuar el ciclo de la cría.
Cría de T. solanivora. a. Cámara de desarrollo larval y pupamiento, b. Cámaras de oviposición.
Determinación de la capacidad de daño de una población conocida de Tecia solanivora en papa almacenada
Con el propósito de determinar la capacidad de daño de una población inicial conocida de adultos de T. solanivora sobre tubérculos almacenados, se llevaron a cabo dos experimentos bajo un diseño de bloques completos al azar con cinco tratamientos y tres repeticiones. Los experimentos se realizaron en el laboratorio de entomología bajo las condiciones de luz, temperatura y humedad descritas.
La unidad experimental utilizada consistió en una jaula de madera de 50 cm x 50 cm x 63 cm de alto, con tres paredes de muselina, la puerta y la parte superior en vidrio. Dentro de la jaula se introdujo una bandeja plástica de 40 x 30 cm, a la cual se le colocó una capa de arena de 5 mm de espesor. Sobre la arena se formó un cúmulo de 100 tubérculos con un peso de 80 a 125 g por tubérculo. La unidad experimental descrita se muestra en la figura 2.
En cada jaula se liberó el número correspondiente de adultos de T. solanivora de acuerdo con los tratamientos descritos en las tablas 1 y 2. Los adultos tenían un promedio de dos días de vida después de la emergencia como tales, periodo en el cual fueron alimentados con una solución de miel al 10%. Una vez liberados permanecieron en las jaulas hasta su muerte, esto ocurrió entre 10 y 12 días después de la liberación. De esta forma se daba la posibilidad a las hembras para que ovipositaran su carga de huevos sobre los tubérculos.
Cada experimento tuvo una duración de 40 días contados desde el día de la liberación de los adultos de la plaga. Tiempo necesario para que transcurriera el ciclo de vida del insecto, desde oviposición hasta que las larvas de cuarto ínstar abandonaran el tubérculo para pupar en la capa de arena colocada para este propósito.

Unidad Experimental. Jaula con bandeja y pila de 100 tubérculos de papa parda pastusa pareja.
En el primer experimento se usó una proporción sexual de 1,5♀:1♂, tomando como referencia para el tratamiento las hembras requeridas y el correspondiente número de machos, como se presenta en la tabla 1.
Tratamientos para determinar el daño causado por una población de T. solanivora en papa almacenada en el experimento con una proporción sexual 1.5♀:1♂
En el segundo experimento no se determinó el sexo de los adultos previamente. Se asumió una proporción sexual 1♀:1♂, que es la más ampliamente aceptada para la especie (Notz 1995; Herrera 1997; Trillos y Álvarez 1998; Araque y García 1999) y se liberaron parejas. Los tratamientos se describen en la tabla 2.
En cada uno de los experimentos y tratamientos se evaluaron las siguientes variables: peso inicial de los 100 tubérculos, número de tubérculos afectados, peso final de los tubérculos sanos y peso final de los tubérculos afectados.
Tratamientos para determinar el daño causado por una población de T. solanivora en papa almacenada
Determinación de los sitios de oviposición de Tecia solanivora en tubérculos almacenados
Para conocer los sitios de oviposición preferidos por la plaga, se realizó un experimento en el cual se utilizó la misma unidad experimental descrita. Se utilizaron dos jaulas y en cada una de ellas se liberaron diez parejas de T. solanivora de un día de vida.
La evaluación se realizó a los ocho días después de liberados los adultos de la polilla guatemalteca, tiempo en el cual se ha logrado el máximo de oviposición y las primeras posturas no han eclosionado. La inspección de los tubérculos se inició desde la parte superior hasta la base del montón, tomando cada uno de los tubérculos, revisándolos minuciosamente, incluso debajo de los terrones adheridos a la piel, para ubicar las posturas y los sitios de oviposición.
Determinación de la preferencia de oviposición de Tecia solanivora en cuanto al estado de limpieza de los tubérculos
Teniendo en cuenta que la comercialización de papa para consumo se realiza con papa sucia (como llega del campo) y excepcionalmente con papa lavada principalmente en cadenas de supermercados, y además que la papa semilla también es almacenada sucia, se realizó un experimento para comparar el comportamiento de oviposición de la plaga, sobre tubérculos limpios y tubérculos sucios.
Se utilizó la misma unidad experimental antes descrita. Se emplearon cuatro jaulas, en dos de ellas se pusieron tubérculos limpios, en las otras dos tubérculos sucios, en cada una de las jaulas se colocaron 100 tubérculos y se liberó una población de diez parejas de T. solanivora de un día de vida.
La evaluación se realizó dividiendo horizontalmente cada uno de los montones en cuatro estratos, superior, alto, medio y bajo. Se observaron los tubérculos de cada uno de los estratos siguiendo la metodología de evaluación descrita en el experimento de sitios de oviposición. Se hizo un registro del número de tubérculos por estrato sobre los cuales la plaga ovipositó.
Resultados y Discusión
Determinación de la capacidad de daño de una población conocida de Tecia solanivora en papa almacenada
Los resultados obtenidos en el primer experimento con una población conocida de la plaga como lo indican los tratamientos que se listan en la tabla 1, proporción sexual de 1,5♀:1♂, arrojaron los promedios que se presentan en la tabla 3 para cada una de las variables evaluadas.
Para la variable número de tubérculos afectados, los datos se ajustaron a la distribución normal (Shapiro - Wilk = 0,96) y se sometieron a un análisis de varianza donde la prueba de F presentó diferencias altamente significativas entre tratamientos (F = 30,72; g.l.: 4,8; P<0,01) pero no entre bloques.
Se hizo una prueba de comparación entre tratamientos a través de un análisis de regresión, donde el mejor ajuste se presentó en el modelo polinomial (F = 183,4; g.l.: 3,10; P<0,01) con un R2 = 98% (P<0,01). Para este modelo se obtuvo la siguiente ecuación:
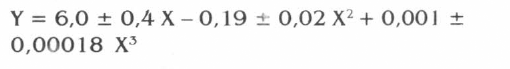
Y = Número de tubérculos afectados
X = Hembras de Tecia solanivora
Con esta ecuación se realizó la gráfica que se presenta en la figura 3, en la cual la variable, número de tubérculos afectados frente al aumento en la densidad de la plaga (número de hembras) presenta una curva sigmoidea con dos puntos de inflexión.
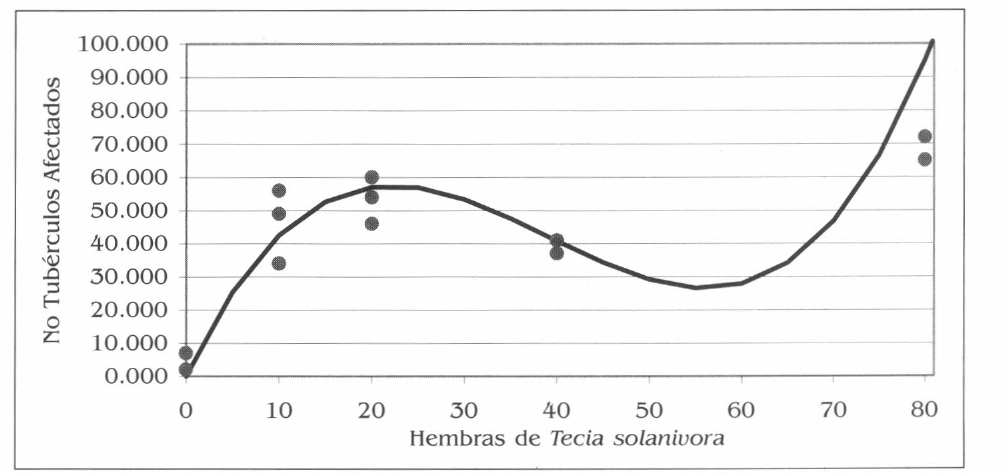
Efecto de diferentes densidades de la plaga en el número de tubérculos afectados. La curva representa la función polinomial y los puntos corresponden a los valores reales de las repeticiones.
Promedios de las variables peso y número de tubérculos afectados en el experimento con una proporción sexual de 1,5♀: 1♂
Promedios seguidos por la misma letra no presentan diferencias significativas, letras diferentes presentan diferencias.
DMT = 3,49, α = 0,05;
DMT = 11,68, α = 0,01.
Para hallar los puntos de inflexión se derivó la función y posteriormente se determinaron los puntos en los cuales la curva sufre una variación. El primer punto de inflexión se presenta con 22 hembras liberadas, y el segundo punto de inflexión con 56 hembras liberadas.
Se observa que con 22 hembras de la plaga se produce un daño en 57,5 tubérculos. De este punto en adelante, aunque se incremente el número de hembras liberadas, se presenta una reducción en el número de tubérculos afectados. Sin embargo, en el punto que corresponde a 56 hembras se presenta un daño en 26,5 tubérculos, a partir de allí, se inicia un nuevo incremento en el número de tubérculos afectados de manera creciente, fenómeno que será analizado y discutido más adelante.
Los datos obtenidos para las variables peso inicial y peso final siguen una distribución normal (Shapiro - Wilk= 0,97). Se efectuó la misma prueba de comparación entre tratamientos. El análisis estadístico de covarianza indicó que hubo diferencias significativas entre tratamientos (F = 4,45; g.l.: 4,7; P<0,05) más no entre repeticiones.
Se ejecutó la prueba de comparación entre tratamientos a través de un análisis de regresión, la respuesta obtenida fue un modelo lineal con un R2 del 94% (P<0,01) y la siguiente ecuación:
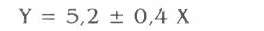
Y = Peso Perdido (Peso inicial — Peso Final)
X = Hembras de Tecia solanivora
Con esta ecuación se conformó la gráfica de la figura 4 sin intercepto, en la cual se observa el comportamiento en pérdida de peso frente al número de hembras liberadas, o cambios en la densidad de la plaga.
Para ajustar la respuesta de la variable peso, se hizo una corrección de todos los datos por el valor promedio de peso perdido del tratamiento testigo, T1, para eliminar el efecto de la pérdida de peso debida a la deshidratación natural que sufren los tubérculos durante el almacenamiento.
Para esta variable por cada hembra de T. solanivora liberada se reduce la masa inicial de los tubérculos en 5,22 g. Se observa en la figura 4 cómo la respuesta es directamente proporcional; es decir, a mayor número de hembras liberadas mayor peso perdido.
Para el segundo experimento se asumió una proporción sexual de la plaga de 1♀:1♂ que es la más ampliamente encontrada en los estudios biológicos de la especie. Se liberó el número de parejas que se indica en la tabla 2, sin determinar el sexo previamente. Los promedios de los resultados obtenidos se presentan en la tabla 4.
Promedios de las variables peso y número de tubérculos afectados en el experimento con una proporción sexual 1♀:1♂
Promedios seguidos por la misma letra no presentan diferencias significativas, letras diferentes presentan diferencias. * DMT = 2,36, α = 0,01; ** DMT = 13,15, α = 0,01.
Los resultados logrados para la variable número de tubérculos afectados con datos que cumplen una distribución normal (Shapiro - Wilk= 0,96) se sometieron a un análisis de varianza donde la prueba de F presentó diferencias altamente significativas entre tratamientos (F= 52,30; g.l.: 4,8; P<0,01) pero no entre bloques.
Se hizo una prueba de comparación entre tratamientos a través de un análisis de regresión en la respuesta de esta variable, donde el mejor ajuste se presentó en el modelo polinomial (F=287,9; g.l.: 3,12; P<0,01) con un R2 = 98% (P<0,01). Para este modelo se obtuvo la siguiente ecuación:
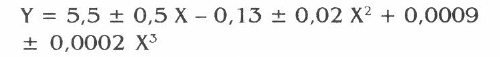
Y = Número de tubérculos afectados
X = Parejas de T. solanivora
Con esta ecuación se realizó la gráfica que se presenta en la figura 5, en la cual se muestra la respuesta de la variable número de tubérculos afectados frente a un incremento en el número de parejas de la plaga liberadas; al igual que en el primer experimento, se presenta una curva sigmoidea con dos puntos de inflexión.
Los puntos de inflexión se encontraron al derivar la ecuación, el primer punto de inflexión se presentó con la densidad de 33 parejas y el segundo punto con la densidad de 60 parejas.
Con la liberación de 33 parejas de la plaga se obtienen 74 tubérculos afectados y de este punto en adelante al incrementar el número de parejas se reduce levemente el número de tubérculos afectados. Sin embargo, a partir de 60 parejas, con un daño en 63,7 tubérculos, se inicia un incremento en el número de tubérculos afectados de manera creciente.
Los datos obtenidos para las variables peso inicial y peso final también se ajustaron a una distribución normal (Shapiro-Wilk = 0,96) y se sometieron a la misma prueba de comparación entre tratamientos. El análisis estadístico de covarianza indicó que hubo diferencias significativas entre tratamientos (F= 16,11; g.l.: 4,7; P<0,01) y no entre bloques.
Se realizó un análisis de regresión para observar el ajuste de los resultados a los diferentes modelos y se obtuvo el mejor resultado con el modelo lineal (F=122,21, g.l.: 1,14; P<0,01) que presentó un R2= 90% (P<0,01), y la siguiente ecuación:
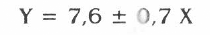
Y = Peso Perdido (Peso Inicial - Peso Final)
X = Parejas de T. solanivora
La gráfica de la figura 6 sin intercepto, expresa la respuesta lineal de la variable peso al aumento en el número de parejas liberadas, en este experimento en el que se asumió una relación de sexos 1♀:1♂.
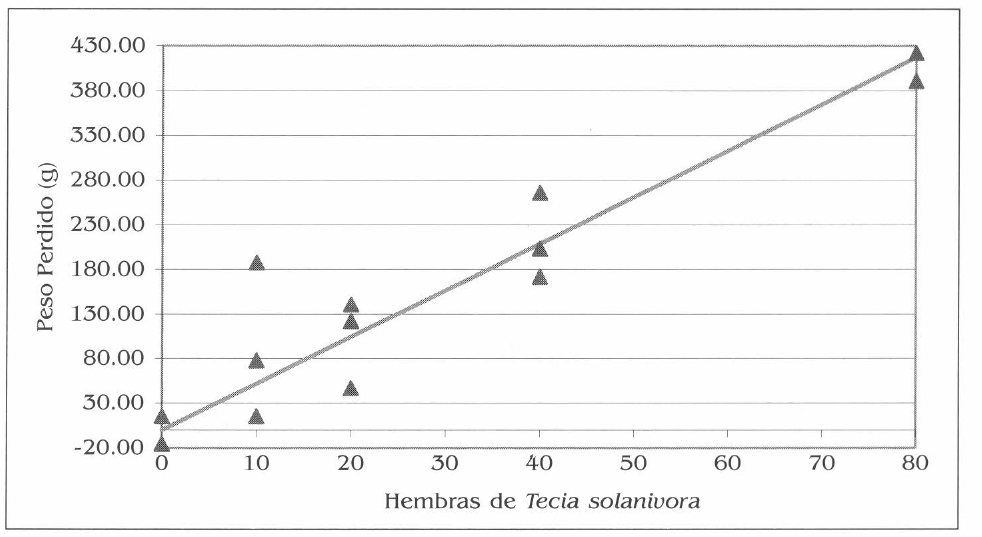
Efecto de diferentes densidades de la plaga en el peso perdido en los tubérculos. La línea continua representa la función lineal y los puntos corresponden a los valores en cada repetición.
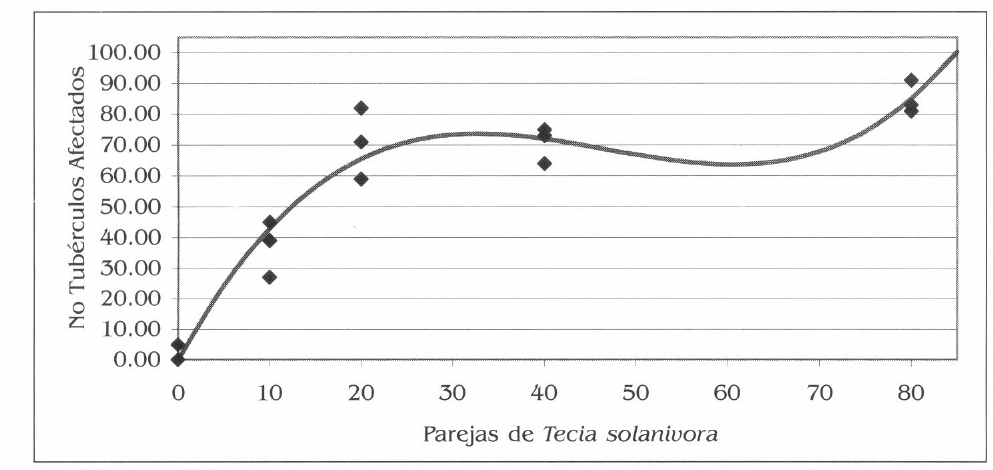
Efecto de diferentes densidades de la plaga en el número de tubérculos afectados. La curva representa la función polinomial y los puntos corresponden a los valores reales de las repeticiones.
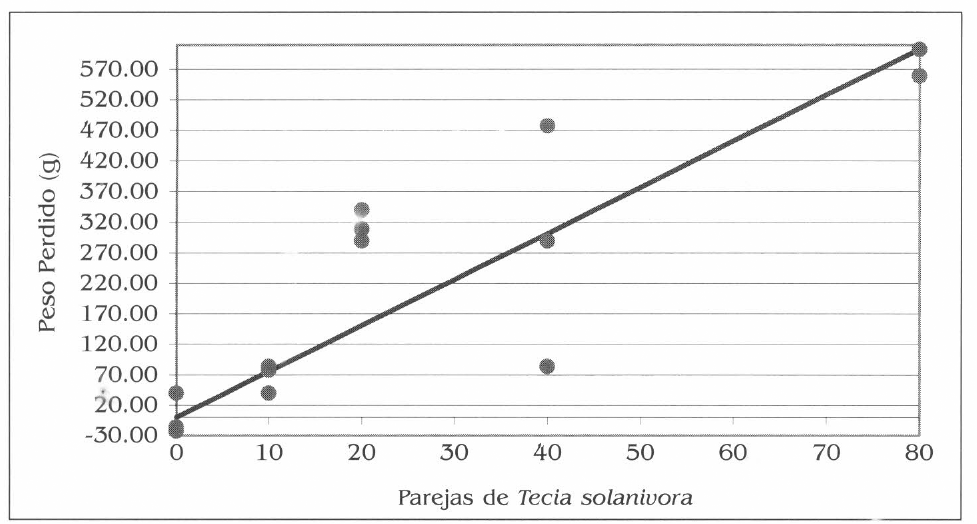
Efecto de diferentes densidades de la plaga en el peso perdido en tubérculos. La línea continua representa la función lineal y los puntos corresponden a los valores en cada repetición.
Igual que en el primer experimento, para ajustar la repuesta de la variable peso, se realizó una corrección de todos los datos por el valor promedio de peso perdido del tratamiento testigo, T1, para eliminar el efecto de la pérdida de peso debida a la deshidratación natural que sufren los tubérculos durante el almacenamiento.
Para esta variable por cada pareja de T. solanivora liberada se reduce la masa inicial de los tubérculos en 7,6 g. Se observa como la respuesta es directamente proporcional; es decir, a mayor número de parejas liberadas mayor peso perdido.
Para los dos experimentos se realizó una correlación entre las variables número de tubérculos afectados y peso, se observó que existe una correlación positiva con un valor de R=0,78 (P<0,01). Esta correlación indica que las variables están asociadas de manera directamente proporcional; es decir, que al aumentar el número de tubérculos afectados se da una mayor diferencia de peso para el mismo número de parejas o hembras de T. solanivora liberadas.
Para ambos experimentos se obtuvo una respuesta polinomial en la variable número de tubérculos afectados. La relación que existe entre las diferentes densidades de hembras o parejas de T. solanivora y el sustrato disponible, se puede explicar a partir de los modelos de competencia intraespecífica.
La tendencia creciente que se observa en las curvas de las figuras 3 y 5, se da hasta llegar a las densidades de 22 hembras y 33 parejas, en este lapso es probable que los individuos expresen su potencial reproductivo, como ocurre en una población que está creciendo sin llegar a poner en riesgo la capacidad de carga del medio. Sin embargo, al incrementar las densidades de la plaga a partir de estos puntos, se observa que disminuye el número de tubérculos afectados. Es en este punto donde puede manifestarse un efecto de la competencia intraespecífica, que puede ser por alimento, espacio o sitios de oviposición. Como mecanismo de regulación de la población la plaga puede ejercer un control de la natalidad, o utilizar mecanismos como la marcación de los sitios de oviposición para evitar que otras hembras ovipositen en tubérculos con posturas, marcación de los tubérculos a través de las heces para evitar que otras larvas penetren en los túneles o la migración de larvas de primer ínstar; todo esto para evitar la pérdida de la posición de equilibrio dentro del sistema, y no sobre explotar el recurso. Este comportamiento se mantiene hasta una densidad de 56 hembras en el primer experimento y 60 parejas en el segundo.
A partir de allí empieza de nuevo un incremento acelerado en el número de tubérculos afectados. Este aumento se puede explicar por fenómenos de competencia, que puede ser competencia de lucha o competencia de concurso. La competencia de lucha en condiciones ideales involucra la división exacta de los recursos y a partir de allí una división igual de los efectos de la competencia, esto puede ser manifestado por un cambio repentino cuando aumenta la densidad de la población desde una completa supervivencia hasta un 100% de mortalidad. Alternativamente la competencia de lucha puede manifestarse por cambios en la masa corporal o en el número de huevos ovipositados por hembra antes que por mortalidad. La competencia de concurso difiere de la competencia de lucha en que el recurso se reparte de manera desigual; algunos individuos consiguen todo lo que requieren, mientras otros tienen insuficientes recursos para sobrevivir o reproducirse (Hassell 1980).
Por otra parte, el modelo matemático empleado para la variable número de tubérculos presenta un ajuste del 98%; sin embargo, a partir de la densidad de 40 hembras o parejas de la plaga se muestra una amplia variación de la curva hasta la densidad de 80 hembras o parejas, esta variación pudo deberse más al ajuste del modelo polinomial que al comportamiento biológico de la especie, dado que no se probaron más tratamientos dentro del rango de densidades de 40 a 80 hembras o parejas de la plaga, aunque es evidente que la densidad de 80 hembras o parejas presenta el mayor número de tubérculos afectados.
La curva además sugiere, que se da un incremento infinito en el número de tubérculos afectados al aumentar la densidad de hembras o parejas liberadas, este comportamiento puede no tener una lógica dentro de un sistema trófico, lo que indica que es posible que cuando se agote el recurso parte de la población muera, o migre y esto se vería representado en la curva por variaciones, ya que podría estabilizarse, decrecer, ascender u oscilar.
Para ambos experimentos se obtuvo una respuesta lineal directamente proporcional para la variable peso. Los resultados muestran como a mayor densidad de T. solanivora el peso perdido es mayor porque hay un mayor número de larvas capaces de causar daño sobre la masa expuesta.
Los modelos que explican la respuesta de la variable peso perdido con una confiabilidad entre 90% y 94% muestran una línea con un crecimiento constante, que se presentaría en caso que el sustrato estuviera disponible indefinidamente; sin embargo, en el experimento el sustrato fue constante (100 tubérculos) y por lo tanto debe existir una variación en la respuesta lineal que podría estabilizarse y descender cuando el recurso se agote.
Vale la pena resaltar que al momento de realizar las evaluaciones de ambos experimentos se observó que, en el estrato superior de la pila, los tubérculos afectados mostraron un daño leve, en el estrato medio el daño fue moderado y en el estrato inferior el daño fue severo principalmente en los tubérculos del centro del montón. En general, para todos los tratamientos el daño siguió este patrón.
Finalmente, en los dos experimentos realizados el tratamiento donde se llevaron a cabo liberaciones de diez hembras y siete machos y diez parejas de T. solanivora, se presentó un alto número de tubérculos afectados, en el primer experimento el promedio fue de 46,33 tubérculos afectados y en el segundo experimento el promedio fue de 37 tubérculos afectados.
Determinación de los sitios de oviposición de Tecia solanivora en tubérculos almacenados
Las observaciones permitieron concluir que las hembras de T. solanivora colocan sus posturas debajo de la tierra adherida a los tubérculos (Fig. 7a) y alrededor de la zona de las yemas de la papa (Fig. 7b) y en la parte baja y central del montón, donde se presentó el mayor número de tubérculos con posturas. Este experimento permitió concluir que la polilla prefiere ovipositar en los lugares más protegidos tanto en la pila de papa como en el mismo tubérculo.
Determinación de la preferencia de oviposición de Tecia solanivora en cuanto al estado de limpieza de los tubérculos
Los resultados mostraron que el comportamiento de oviposición de T. solanivora en cuanto a la ubicación de las posturas en el montón fue el mismo tanto en papa sucia como en papa limpia, al analizar el número de posturas se encontró que las polillas prefieren los sitios del estrato medio y bajo para ubicarlas. Los resultados se presentan en la tabla 5.
Resultados de oviposición de T. solanivora en papa sucia y en papa limpia
En promedio el número total de tubérculos con posturas para papa sucia fue similar al encontrado en papa lavada (P> 0,05); sin embargo, fue definitivamente mayor el número de huevos por postura en las papas sucias en donde se hallaron grupos de hasta 13 huevos, mientras que en la papa lavada se localizó un menor número de huevos por postura, entre dos y tres huevos y en menos tubérculos.
Con estas observaciones se puede corroborar no sólo que la polilla prefiere los tubérculos más protegidos en la pila para ubicar sus posturas, sino que en el tubérculo busca los lugares con mayor protección.
Las hembras de T. solanivora pueden estar obteniendo estímulos de la tierra adherida a los tubérculos para ubicar sus posturas en mayor número en papa sucia comparado con papa limpia, estos estímulos pueden ser la textura y olores específicos del suelo, y la protección de enemigos naturales que puede tener al ubicar las posturas debajo de los terrones adheridos al tubérculo.
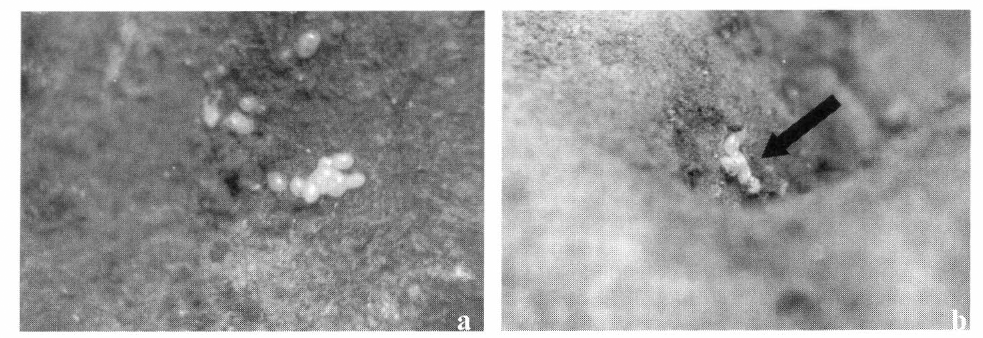
Sitios de oviposición de la polilla guatemalteca de la papa. a. Posturas observadas al retirar el terrón, b. Posturas alrededor de las yemas del tubérculo.
Conclusiones y Recomendaciones
En los experimentos de capacidad de daño de la plaga se presentó una respuesta polinomial para número de tubérculos afectados debido posiblemente a la competencia intraespecífica cuyo efecto se hizo visible a partir de 22 hembras y 33 parejas de Tecia solanivora liberadas en montones de papa. El peso perdido tuvo una relación directamente proporcional entre la densidad de la plaga y el peso perdido, determinándose que por cada hembra de T. solanivora liberada se da una pérdida de peso entre 5 y 7 g en el sustrato expuesto.
Existe un patrón en el daño causado por la plaga dentro del arrume, en el estrato superior el daño en los tubérculos fue leve, en el estrato medio el daño fue moderado y en el estrato inferior el daño fue severo. Lo que sugiere que las medidas tomadas para proteger los tubérculos deben concentrarse en este lugar.
La capacidad de daño de las larvas producidas por diez hembras o parejas de Tecia solanivora supera el 30% de tubérculos afectados, considerándose un porcentaje de daño alto en los experimentos realizados.
Se determinó que la hembra prefiere ubicar sus posturas en el sitio más protegido, es decir, en la parte baja y central del montón y en el tubérculo alrededor de la zona de las yemas y debajo de la tierra adherida a la piel. En cuanto al estado de limpieza de los tubérculos, la hembra coloca posturas en un número similar de tubérculos en la papa sucia comparada con la papa limpia; sin embargo, el número de huevos por postura en papa sucia es superior al encontrado en papa limpia, por lo tanto se considera conveniente lavar los tubérculos antes de almacenarlos. Analizando este comportamiento se concluye que es probable que la hembra de la polilla guatemalteca busca los sitios más escondidos y protegidos dentro de la pila de tubérculos para ovipositar.