
Abstract
Summary
Knowledge about the foraging behaviour of parasitoids is essential to estimate the efficiency of biological control agents and to develop parasitoid release strategies. In this paper the behaviour of A. fuscipennis when searching for Trialeurodes vaporariorum nymphs on bean plants was studied under laboratory conditions. The residence time and searching behaviour of A. fuscipennis were determined on plants sprayed or unsprayed with a sulphur-based fungicide, and infested or uninfested with nymphs of T. vaporariorum. The residence time and walking activity of A. fuscipennis on infested, unsprayed plants were higher than that of uninfested or sprayed plants. The residence time of A. fuscipennis was prolonged by both, encounters with unparasitized hosts and by feeding from the plant. Results suggest that: 1) residence time was reduced on fungicide sprayed plants, indicating that the fungicide disturbed A. fuscipennis, when A. fuscipennis is used within an Integrated Pest Managment (IPM) system for A. fuscipennis, special care must be taken on that the release and migration of the parasite to the crop does not happen immediately after a sulfurated fungicide has been applied. 2) Despite being a pro-ovigenic parasitoid that emerges with their life-time egg load ready to oviposit, A. fuscipennis needs to feed on the plant probably because searching for hosts demands extra energy.
Introducción
Los parasitoides pueden contribuir al éxito de la agricultura sostenible regulando poblaciones plaga y disminuyendo la dependencia de plaguicidas químicos. Conocimientos básicos sobre la biología y ecología de un parasitoide son esenciales para evaluar su utilidad como agente de control biológico.
La selección de enemigos naturales, como los parasitoides, se ha realizado hasta hace poco basado en el método de error-ensayo. El proceso de selección se ha efectuado rápidamente y se ha liberado el enemigo natural sin haber conocido y probado ciertas características biológicas y ecológicas importantes. Posteriormente, cuando los problemas aparecen es difícil identificar las causas de las fallas y más aún, es difícil mejorar la situación. Como consecuencia, por un lado, agentes de control promisorios se han rechazado sin ni siquiera conocer las causas de la falla y por otro lado, agentes de control ineficientes se han liberado (van Lenteren y Woets 1988). Por lo tanto, es necesario desarrollar un entendimiento básico de cómo funciona el control biológico y poder hacer predicciones sobre el éxito de un enemigo natural antes de ser liberado en forma masiva. Los estudios de comportamiento son, por lo tanto, un complemento básico de ensayos biológicos que permiten entender de manera global qué tan efectivo puede ser un enemigo natural (van Lenteren y Woets 1988). El desempeño de un parasitoide en el campo o invernadero puede ser estimado al estudiar aquellos parámetros de comportamiento que indican su eficacia y potencialidad como agente de control (van Roermund 1995; Mattiacci et al. 1999).
En el presente estudio se examinó el comportamiento de búsqueda del hospedero de un parasitoide. El sistema tritrófico estudiado está conformado por fríjol (Phaseolus vulgaris L.), una de sus plagas clave, la mosca blanca de los invernaderos Trialeurodes vaporariorum (Westwood) (Homoptera: Aleyrodidae) y su parasitoide ninfal Amitus fuscipennis MacGown & Nebeker.
Fríjol es uno de los cultivos más importantes del Tercer Mundo porque es la fuente de proteína más barata que se puede conseguir y adicionalmente proporciona gran cantidad de calorías (Pachico 1993). En Colombia, el fríjol es atacado por la mosca blanca de los invernaderos T. vaporariorum que puede alcanzar el nivel de plaga clave (Prada et al. 1993). T. vaporariorum es además plaga importante en el ámbito mundial en muchos cultivos de invernadero (van Lenteren. y Martín 2000).
El control de T. vaporariorum se ha realizado en Colombia principalmente a través del uso de pesticidas químicos lo que ha originado problemas de resistencia y toxicidad al agricultor (Cardona et al. 2001; Rodríguez y Cardona 2001). Como una alternativa de control se implementó un programa de manejo con diferentes metodologías, excepto el control biológico, que combinaba el control cultural con la aplicación de insecticidas a un umbral económico previamente establecido (Prada et al. 1993). Al aplicar este programa en el campo con agricultores, se obtuvo una reducción del 54% en el uso de insecticidas y de un 18% en costos (Prada et al. 1993).
Como una adición importante a este programa, se está evaluando como componente biológico el parasitoide Amitus fuscipennis. Este enemigo natural está distribuido en Centroamérica, norte de Sudamérica y el Caribe (Mac Gown y Nebeker 1978). Ciertas características biológicas de A. fuscipennis soportan su potencial como agente de control de T. vaporariorum: a) Produce mayor cantidad de hembras que machos porque se reproduce por telitoquia posiblemente mediada por la rickettzia Wolbachia (Manzano et al. 2001a); b) La tasa intrínseca de crecimiento poblacional (rm) de A. fuscipennis es superior a la de T. vaporariorum a diferentes condiciones de humedad relativa y temperatura (Manzano et al. 2002a) y, c) A. fuscipennis prefiere ovipositar los primeros estadios ninfales de T. vaporariorum, camina rápidamente en el envés de la hoja para buscarlos (1,4 mm s-1) y los oviposita en un tiempo promedio más corto (53 s) (Manzano et al. 2002b) que otros enemigos naturales efectivos de T. vaporariorum como E. formosa (> 100 s) (van Lenteren et al. 1980).
Además de sus características biológicas, el éxito de un parasitoide como agente de control biológico depende en gran parte de su capacidad para buscar y encontrar al hospedero para parasitarlo. El estudio del comportamiento de búsqueda de un enemigo natural, es entonces un paso importante al desarrollar un programa de control biológico (van Roermund 1995; van Lenteren y Manzaroli 1999). Algunos aspectos del comportamiento de búsqueda y oviposición del hospedero de A. fuscipennis fueron descritos por Manzano et al. (2002b).
El proceso de búsqueda de hospedero por parte de un parasitoide se puede dividir en tres fases: localización del hábitat del hospedero, localización del hospedero dentro de ese hábitat y aceptación del hospedero (Vinson 1976). En el presente trabajo se estudian aspectos del proceso de búsqueda una vez que el parasitoide ya ha pasado la primera fase. A. fuscipennis está ubicado en el hábitat del hospedero (planta de fríjol), recorre las hojas de la planta y examina al hospedero para ovipositarlo o rechazarlo.
En este trabajo se probó la hipótesis de que la capacidad de búsqueda de A. fuscipennis estaba influenciada por la calidad de la planta hospedera. La calidad de la planta hospedera estaba determinada por la presencia o ausencia de ninfas de T. vaporariorum y por la presencia o ausencia de un fungicida sulfurado en las hojas trifoliadas del fríjol. En particular, se determinó el tiempo de permanencia de A. fuscipennis en la planta y su actividad de búsqueda.
Materiales y Métodos
Plantas
Las plantas de fríjol (Phaseolus vulgaris L) se sembraron y mantuvieron en condiciones de invernadero a 22 ± 0.4°C; 80.7 ± 19.2% HR y 12L:120. Las plantas no recibieron fertilización o tratamientos químicos previos. Cuando tenían de 15 a 20 días de sembradas se les podó para dejar un sólo trifolio (i.e., una hoja) por planta. Las plantas recibieron los siguientes tratamientos: 1) un grupo se conservó intacto; 2) otro grupo se asperjó con un fungicida sulfurado (3 × 10−3 ml), utilizando un aspersor manual y, 3) un tercer grupo se infestó con ninfas de Trialeurodes vaporariorum. Cada foliolo tenía un área de 34,3 cmβ ± 0.7 ES.
Mosca blanca
Los adultos de T. vaporariorum provinieron de una cría establecida en el Centro Internacional de Agricultura Tropical, CIAT, desde hace 5 años en fríjol (c.v. ICA-Pijao) con insectos colectados en Fusagasugá, Cundinamarca, Colombia. La cría se mantenía a 19°C, 80 ± 10% HR, y 12L:120. Para infestar la planta de fríjol con T. vaporariorum, se tomaron 20 adultos que se confinaron en una jaula pinza colocada en el envés de un foliolo por planta durante 4 horas. Después de este tiempo, los adultos se removieron y la cantidad de huevos presentes se manipuló para posteriormente obtener una densidad de 4 ninfas por foliolo.
Parasitoides
Las hembras de Amitus fuscipennis usadas en este experimento, provinieron de una cría establecida en 1996 en CIAT con individuos colectados en Tenerife, Valle del Cauca, Colombia. La cría se mantuvo utilizando ninfas de T. vaporariorum como hospedero y fríjol (c.v. ICA-Pijao) como planta hospedera. La cría de A. fuscipennis se mantuvo a 19°C, 80 ± 10% HR, y 12L:120.
Tratamientos
El comportamiento de búsqueda del parasitoide se estudió en plantas que tuvieron tres tipos de tratamientos: Tratamiento 1 (Control): plantas no asperjadas con el fungicida sulfurado ni infestadas con ninfas de T. vaporariorum; tratamiento 2: plantas asperjadas con el fungicida y no infestadas con ninfas de T. vaporariorum, y tratamiento 3: plantas no asperjadas con el fungicida e infestadas con ninfas de T. vaporariorum. Las plantas infestadas tenían 4 ninfas de primer o segundo ínstar de mosca blanca. Para cada tratamiento se utilizaron 20 plantas y 20 hembras diferentes de A. fuscipennis.
Observación del comportamiento
Las observaciones del comportamiento de búsqueda de A. fuscipennis se realizaron en un cuarto aclimatado a 19°C y 80 ± 10% HR en CIAT, simulando las condiciones climáticas de la zona de montaña donde frecuentemente es encontrado. La planta a observar se rodeaba de cuatro plantas intactas y de la misma edad para simular el ambiente del cultivo. La luz caía directamente sobre la planta con una intensidad de 2700 lux. Al comienzo de cada observación, una hembra de A. fuscipennis recién emergida y sin experiencia de oviposición, se introdujo en el envés de alguno de los tres foliolos utilizando una cápsula de gelatina (2,5 x 0,8 cm). Los parasitoides se depositaron siempre en el centro del envés del foliolo. En foliolos infestados los parasitoides se dejaron cerca de los hospederos.
La hembra de A. fuscipennis se observó a ojo inmediatamente después de ser dejada en el foliolo y su comportamiento se siguió hasta que voluntariamente abandonó la planta. Cuando el parasitoide permaneció 15 min quieto se consideró inactivo, se retiró y se cambió por un nuevo individuo. Cada hembra se utilizó y observó una sola vez. El comportamiento de búsqueda se registró en un microcomputador utilizando el programa para el estudio de comportamiento "The Observer" versión 3.0 para Windows (Noldus 1991).
Los siguientes parámetros de comportamiento de A. fuscipennis se diferenciaron y registraron: caminar, parar, limpiar el cuerpo, encontrar el hospedero, examinar el hospedero con las antenas y, tomar una postura de oviposición. La dirección de partida del parasitoide se examinó como hacia arriba, horizontal o hacia abajo. La localización del parasitoide en el foliolo se anotó como centro superior o inferior o borde superior o borde inferior. El borde de la hoja se definió con una anchura de 0,5 cm. La ubicación del parasitoide en la planta se señaló en hojas (trifolio), tallo (incluyendo el pecíolo) o en la estipela (estructuras semejantes a un pequeño foliolo ubicadas, dos en el pecíolo del foliolo central y otras dos. en cada pecíolo de los foliolos laterales).
Parámetros medidos
Se midió (1) tiempo de permanencia: que es el tiempo total que las hembras de A. fuscipennis permanecieron en las plantas; (2) actividad de búsqueda: fue expresada como el porcentaje de tiempo que el parasitoide caminó con base en el total de tiempo de permanencia en la planta.
Cuando se ofrecieron hospederos al parasitoide también se determinó su tiempo de permanencia en la planta sin incluir el tiempo de manipulación del hospedero (encuentro, examen con las antenas y oviposición del hospedero). Para plantas infestadas se midió también, (3) el parasitismo (%) al primer encuentro con hospederos no parasitados. También se midió, (4) el tiempo de partida: definido como el tiempo comprendido entre el encuentro con el último hospedero (encontrarlo, examinarlo con las antenas, ovipositarlo o no) y la partida de A. fuscipennis de la planta.
Análisis de datos
Los porcentajes de tiempo se transformaron a raíz cuadrada del arcoseno del porcentaje porque no tenían una distribución normal (Murdie 1972). Mediante análisis de varianza se compararon los diferentes valores transformados y luego se realizó la prueba múltiple de Student-Newman-Keuls.
Resultados
Tiempo de permanencia
El tiempo de permanencia de A. fuscipennis en las plantas visitadas se afectó por el tratamiento (Tabla 1). El tiempo de residencia fue más largo cuando el parasitoide encontró los hospederos (2,7 h) en plantas infestadas y sin fungicida. Las hembras de A. fuscipennis abandonaron las plantas asperjadas con el fungicida más rápidamente (0,9 h) que las plantas no infestadas ni asperjadas (1,7 h). El tiempo promedio de permanencia de A. fuscipennis fue significativamente diferente entre los tres tratamientos (Kruskal-Wallis, P < 0.05, Tabla 1).
La cantidad de tiempo (%) que A. fuscipennis caminó en la planta también se afectó por el tratamiento (Tabla 1). El tiempo invertido caminando (%) en plantas sin mosca y asperjadas (54,7%) fue similar al de plantas sin mosca y no asperjadas (62,1%), pero fue significativamente superior en hojas infestadas (78,3%) (Kruskal-Wallis, P<0.05, Tabla 1). En hojas infestadas la actividad de búsqueda de A. fuscipennis no aumentó ni disminuyó significativamente después de ovipositar.
En hojas infestadas el tiempo promedio invertido por una hembra de A. fuscipennis en la planta, sin contar el tiempo de manipulación del hospedero (encontrarlo, examinarlo con antenas y ovipositor, ovipositarlo) fue de 9.611,3 s ± 993 (2,7 h). ver Tabla 2. El tiempo invertido por las hembras de A. fuscipennis en manipular el hospedero fue de 2,8 s ± 0.5, equivalente al 3% del tiempo de permanencia en la planta (Tabla 2). El resto del tiempo A. fuscipennis lo invirtió caminando, limpiando su cuerpo de la cera que cubre a las ninfas de mosca blanca, comiendo o estando quieto en la hoja.
En hojas infestadas el tiempo de partida de A. fuscipennis de la planta, después del último encuentro con el hospedero, fue de 7.385,4 s (2,05 h). Este tiempo corresponde al 75% del tiempo de permanencia del parasitoide en la planta incluyendo el tiempo de manipulación del hospedero. En las plantas infestadas todos los parasitoides encontraron los hospederos. El porcentaje de oviposición al primer contacto con el hospedero fue del 80%. En el presente experimento el número de encuentros u oviposiciones no influyó en el tiempo de permanencia de A. fuscipennis en la planta hospedera.
Localización en la planta infestada
A. fuscipennis recorrió toda la planta buscando hospederos. Después de ser dejado en el envés de la hoja, A. fuscipennis caminó rápidamente y se movió entre los foliolos ya sea comunicándose a través de los pecíolos o pasando de un foliolo al otro cuando los bordes de estos se tocaban. Las cuatro partes principales visitadas por A. fuscipennis fueron las hojas, pecíolos, tallos y estipelas. Los parasitoides se detuvieron en las estipelas aparentemente a alimentarse (Fig. 1).
Considerando todos los tratamientos, los parasitoides invirtieron más tiempo buscando en las hojas que en otras partes de la planta: 95,3% del tiempo total de permanencia en plantas no asperjadas ni infestadas, 95,6% en plantas asperjadas y no infestadas y, 93,9% en plantas infestadas y no asperjadas. El tiempo invertido en pecíolos y tallos fue del 4,3% en plantas no asperjadas ni infestadas, del L7% en plantas asperjadas y no infestadas, y del 4,5% en plantas infestadas y no asperjadas. El tiempo invertido en la estipela fue del 0,4, 2,7 Y 1,6% respectivamente.
El número promedio de foliolos visitados fue significativamente inferior en plantas asperjadas (2.3, Tabla 3), porque el parasitoide abandonó la planta más rápidamente. Al buscar hospederos, A. fuscipennis caminó por todo el foliolo cambiando del haz al envés y viceversa. El tiempo invertido en buscar en el envés del foliolo fue significativamente mayor en plantas infestadas y no asperjadas (7.800,7 s) que en plantas no asperjadas ni infestadas (5.081,2 s) o no infestadas y asperjadas (2.644,6 s) (Tabla 3). El tiempo invertido en la parte central del envés fue significativamente mayor en foliolos infestados y no asperjados (6.443,2 s) comparado con los otros dos tratamientos (Tabla 3).
Tiempo promedio de permanencia (s) y tiempo promedio (%) invertido en caminar por Amitus fuscipennis en hojas no infestadas con ninfas de Trialeurodes vaporariorum y asperjadas con un fungicida sulfurado, no infestadas ni asperjadas, e infestadas y no asperjadas. Los promedios van acompañados del error estándar
Promedios con diferentes letras difieren significativamente (prueba de Student-NewmanKeuls, P < 0.05, después de ANOVA en una vía con Kruskal-Wallis, P < 0.0001).
Promedios con diferentes letras difieren significativamente (prueba de Student-NewmanKeuls, P < 0.05, después de ANOVAen una vía con Kruskal-Wallis, P < 0.0001).
Tabla 2. Tiempo promedio de permanencia (TP. excluyendo el tiempo de manipulación del hospedero); manipulación del hospedero (MN, % de tiempo de permanencia); número promedio de encuentros con el hospedero (EN) y posturas de oviposición (PO); proporción de hospederos ovipositados al primer encuentro (NO), y tiempo de partida (TP) de 20 hembras de A. fuscipennis después del último encuentro con el hospedero. Las medias son presentadas con el error estándar
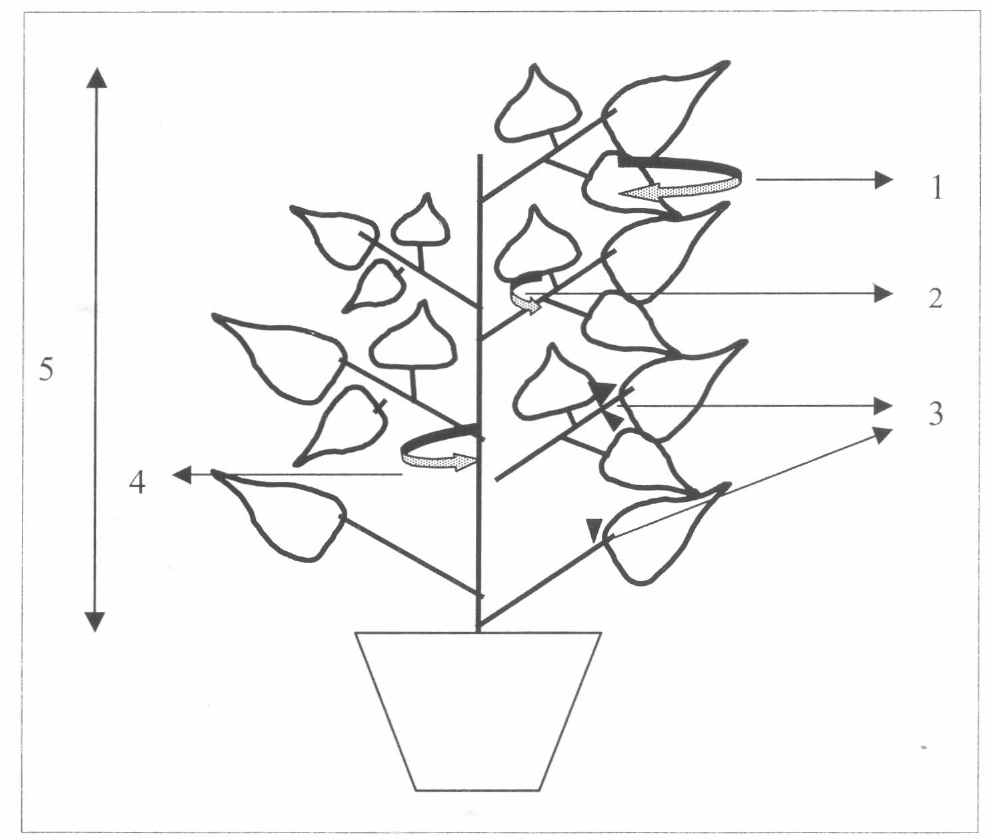
Desplazamiento y ubicación de A. fuscipennis en la planta de fríjol: 1. Haz-envés 2. Foliolo-pecíolo 3. Estipelas: aquí también ocurre alimentación 4. Pecíolo - tallo 5. A lo largo de todo el tallo.
Cuando había hospederos, los parasitoides invirtieron el 83% del tiempo de permanencia en el envés (excluyendo el tiempo de manipulación del hospedero) en su centro (Tabla 3), donde fueron introducidos y donde la mayoría de los hospederos estaban localizados. En foliolos no infestados y asperjados este porcentaje disminuyó a 51,3 y a 46,4% en plantas no infestadas ni asperjadas (Tabla 3). El tiempo invertido por el parasitoide en el borde del haz no fue significativamente diferente entre foliolos no infestados ni asperjados (673,9 s) y foliolos infestados y no asperjados (724,2 s) (Tabla 3). El tiempo invertido en el borde del haz fue significativamente más corto en foliolos asperjados y no infestados (389,9 s) comparado con los otros tratamientos (Tabla 3).
Las estipelas fueron más visitadas por A. fuscipennis en plantas infestadas (80%) que en plantas no infestadas ni asperjadas (50%) o no infestadas y asperjadas (40%). A. fuscipennis tomó alimento de las estipelas, especialmente en plantas infestadas (50% de los parasitoides) demorándose una hembra en promedio 85,0 s en la toma de alimento.
El sitio del foliolo de donde A. fuscipennis voló y abandonó la planta fue similar para todos los tratamientos. La mayoría de los parasitoides (85%) partieron del borde del haz; 8,3% partieron del tallo, 3,3% del borde del envés y 1,7% del centro del haz o del envés. La dirección del vuelo fue hacia arriba en la mayoría de los parasitoides (86%); hacia abajo para el 12% y horizontal para el 2% de los parasitoides.
Número de foliolos de fríjol visitados y tiempo promedio invertido (s) por Amitus fuscipennis en diferentes zonas del foliolo durante el proceso de búsqueda de hospedero. Los promedios van acompañados del error estándar
Excluyendo tiempo de manipulación del hospedero.
Promedios con diferentes letras difieren significativamente (prueba de Student-NewmanKeuls, P < 0.05, después de ANOVA en una vía con Kruskal-Wallis, P < 0.001).
Promedios con diferentes letras difieren significativamente (prueba de Student-NewmanKeuls, P < 0.05, después de ANOVA en una vía con Kruskal-Wallis, P < 0.0001).
Promedios con diferentes letras difieren significativamente (prueba de Student-NewmanKeuls, P < 0.05, después de ANOVA en una vía con Kruskal-Wallis, P < 0.0001).
Promedios con diferentes letras difieren significativamente (prueba de Student-NewmanKeuls, P < 0.05, después de ANOVA en una vía con Kruskal-Wallis, P = 0.044).
Discusión
Tiempo de permanencia
Los resultados obtenidos permitieron aceptar la hipótesis nula de que el comportamiento de búsqueda de A. fuscipennis es influenciado por la "calidad" de la planta donde busca hospederos. Al comparar plantas libres de mosca versus plantas con mosca blanca, se encontró que el tiempo de permanencia de A. fuscipennis fue mayor en plantas infestadas. Esto se explica porque cada encuentro y especialmente cada oviposición del parasitoide, prolongan su estadía en el foliolo y en la planta. Resultados similares fueron encontrados por van Roermund y van Lenteren (1995), quienes señalaron que el tiempo de permanencia de Encarsia formosa Gahan en foliolos de tomate sin ninfas de T. vaporariorum fue en promedio 20 min, tiempo que aumentó a 144 min cuando había 4 ninfas de T. vaporariorum por foliolo.
De otro lado, el tiempo de permanencia de A. fuscipennis fue menor a 1 h cuando las hojas no estaban infestadas con ninfas de T. vaporariorum y habían sido asperjadas con un fungicida sulfurado. Cuando el fungicida estaba ausente y tampoco había mosca blanca, el tiempo de permanencia del parasitoide aumentó a 1,7 h.
Agricultores de algunas zonas del Valle del Cauca utilizan fungicidas sulfurados para el control de ciertas enfermedades vegetales (CIAT 1999). Aunque este tipo de fungicida no mata a la mosca blanca (E. Valencia, comunicación personal), los presentes resultados muestran que interfiere con el control biológico reduciendo el tiempo de permanencia de A. fuscipennis, un enemigo natural promisorio de T. vaporariorum (Manzano 2000). Aparentemente las hembras de A. fuscipennis perciben la presencia del fungicida y abandonan las plantas asperjadas más rápidamente que las plantas no asperjadas (el tiempo de permanencia en plantas no asperjadas fue 0,7 h más largo que en plantas asperjadas). Adicionalmente, en plantas asperjadas el parasitoide fue menos activo (i.e. caminó menos tiempo). Si A. fuscipennis llega a ser utilizado en programas de Manejo Integrado de Plagas (MIP) para mosca blanca, las aplicaciones de fungicidas sulfurados deben ser cuidadosamente sincronizadas con la liberación o llegada del parasitoide al cultivo para evitar interferencias. La información existente sobre el efecto de fungicidas en el comportamiento de búsqueda de los parasitoides de mosca blanca y de enemigos naturales en general, es escasa. Es más comúnmente registrado el efecto mortal de los insecticidas y herbicidas en parasitoides de mosca blanca (Koppert http://www.kopert.nl 2002).
En plantas infestadas con mosca blanca, A. fuscipennis caminó más en busca de sus hospederos porque probablemente encuentros con hospederos y oviposiciones incrementaron su motivación de búsqueda. El tiempo de partida de la planta hospedera (7.385,4 s) correspondió al 75% del tiempo de permanencia de A. fuscipennis en la planta hospedera. Esto significa que el parasitoide encontró sus hospederos rápidamente (en el 25% del tiempo de permanencia) y el resto del tiempo lo invirtió caminando activamente en busca de más hospederos, limpiando su cuerpo y tomando alimento de las estipelas. Todos los parasitoides encontraron las ninfas rápidamente porque fueron depositados en el envés de los foliolos muy cerca de los hospederos.
Localización en la planta hospedera
La presencia de hospederos no parasitados en plantas no asperjadas estimularon al parasitoide a incrementar su tiempo de permanencia a 2,7 h. Sin embargo en el presente estudio, no se pudo probar estadísticamente que el tiempo de permanencia del parasitoide en la planta estuviera correlacionado con el número total de encuentros con el hospedero o con el número total de oviposiciones. El número de réplicas (20) fue tal vez muy bajo o quizás la densidad de infestación de mosca blanca fue baja. Van Roermund y van Lenteren (1995) encontraron que el parasitoide de T. vaporariorum, Encarsia formosa Gahan permaneció más tiempo en la planta hospedera después de encontrar y ovipositar hospederos no parasitados.
A. fuscipennis al buscar en la hoja permaneció más tiempo en el envés que en el haz del foliolo. En plantas no asperjadas e infestadas, el parasitoide invirtió más tiempo buscando en el envés de la hoja comparado con plantas no infestadas, lo que indica que encuentros con hospederos no parasitados producen un efecto de retención del parasitoide en la hoja al encontrar este tipo de hospederos. Considerando que las ninfas de mosca blanca se localizan casi exclusivamente en el envés de las hojas, mostrar preferencia por este lado de la hoja es una ventaja adaptativa para A. fuscipennis. Cuando los hospederos estaban presentes, A. fuscipennis prefirió el centro de la hoja para buscarlos, posiblemente debido a que el parasitoide tiene como estrategia buscar sus hospederos en un área restringida. Este comportamiento es una adaptación para encontrar hospederos que, como la mosca blanca, se distribuyen espacialmente de manera agregada (Godfray 1994).
Las hembras de A. fuscipennis se alimentaron principalmente de las estipelas en plantas infestadas con mosca blanca, lo que incrementó el tiempo de permanencia del parasitoide en la planta. No hay una evidencia morfológica de que existan secreciones en las estipelas de Phaseolus vulgaris como sí ocurre en otras especies de Phaseolus (D. Debouck, comunicación personal). Sin embargo se observó durante el presente estudio, que A. fuscipennis tomaba ciertas sustancias de las estipelas. Este aspecto merece más investigación. Un efecto importante del ofrecimiento de alimento por parte de la planta hospedera es la retención del enemigo natural (Cortesero et al. 2000). Los resultados del presente trabajo sugieren que A. fuscipennis se alimentó durante más tiempo en la planta hospedera porque la actividad de búsqueda del hospedero y su oviposición le exigió más requerimientos energéticos. A. fuscipennis es un parasitoide proovigénico que emerge con su carga de huevos lista para ser ovipositada (Gerling 1990), no se alimenta de su hospedero (Manzano et al. 2001b), pero toma alimento de la planta hospedera. Por lo tanto A. fuscipennis puede ser retenido en la planta como resultado de encuentros con hospederos y toma de alimento.
Conclusiones
En cuanto al comportamiento de búsqueda de A. fuscipennis se puede concluir lo siguiente:
El tiempo de permanencia del parasitoide fue mayor en plantas infestadas y no asperjadas con un fungicida sulfurado (2,7 h) comparado con plantas no infestadas ni asperjadas (1,7 h) o no infestadas y asperjadas (< 1h).
El parasitoide caminó significativamente más tiempo buscando hospederos en plantas infestadas y no asperjadas (78,3%) que en plantas no infestadas y no asperjadas (62,1%) o no infestadas y asperjadas (54,7%).
El parasitoide prefirió buscar sus hospederos en foliolos infestados, en el centro del envés.
El parasitoide ovipositó en el 80% de los hospederos no parasitados encontrados por primera vez.
Estos resultados señalan una eficiencia de búsqueda alta de A. fuscipennis, debido a que este parasitoide busca más intensamente en los sitios donde se espera encontrar hospederos y una vez encontrados permanece en el área aumentando la probabilidad de encontrar más hospederos. A. fuscipennis presentó también un porcentaje alto de parasitismo a densidades bajas del hospedero. Aunque en condiciones de campo las poblaciones de mosca blanca se encuentran en densidad alta, se espera que su condición de parasitoide pro-ovigénico le permita potencialmente reducir estas poblaciones altas de la plaga.
Agradecimientos
Se agradece a H. Morales (CIAT) su colaboración en las actividades de invernadero y a C. Stolk (Wageningen University) sus comentarios al manuscrito. Esta investigación fue financiada por la rundación Holandesa para el Fomento de la Investigación en los Trópicos WOTRO.