
Abstract
Summary
The diamond back moth, Plutella xylostella L., (Lepidoptera: Plutellidae), a pest of significance in the production of cruciferas in the Antioquean east. Virulence of 17 native isolations and three commercial products of B. bassiana on P. xylostella was evaluated. Three of them, wich showed more activity on the mentioned pest, were selected. The effect of these three isolations was also tested on larvae of Copitarsia sp., Peridroma sp. and Leptophobia aripa (Boisduval), which are pests that are also associated with cruciferas in that region. Pathogenicity of B. bassiana on P. xylostella was characterized by means of these variables: percentage of mortality, half-lethal time and intrinsic mortality. The effect in the field was evaluated in plots of 30 plants, in a randomized complete blocks design, with five treatments and four replications. Treatments involve two native isolations CLS 029 and CLS 003 (those with high biological activity on P. xylostella), the commercial product Bauveril and two controls. Half-lethal concentrations for these three treatments were 2.7x105, 2.1x106 and 2.4x106 c/ml, respectively. The three examined sources of B. bassiana caused mortality to P. xylostella under field conditions. The infection of CLS 029 isolation stands out on L. aripa larvae. Only one larva of Copitarsia sp. was infected with the CLS 003 isolation. None of the treatments affected Peridroma sp.
Introducción
Las hortalizas constituyen un renglón productivo para la economía campesina del oriente antioqueño colombiano. Dentro de ellas, las crucíferas juegan un papel importante con cerca de 1.900 ha sembradas en los municipios de Marinilla y El Santuario, principalmente. La producción de crucíferas en Antioquia se enfrenta a diversos problemas insectiles, siendo los de mayor importancia los masticadores de follaje que afectan el rendimiento y la calidad. Algunas de las especies plaga sobresalen porque afectan negativamente el desarrollo y la calidad de estos productos llegando incluso a causar pérdidas considerables. No obstante, sólo las especies Plutella xylostella, Copitarsia sp. y Peridroma sp., de las nueve registradas en cultivos de crucíferas en Antioquia, podrían considerarse limitantes ya que ocasionan daños en las etapas vegetativa y de formación de cabeza afectando así el crecimiento, la calidad y por tanto el ingreso económico de los agricultores (Gómez 1979; Gallego y Vélez 1992).
La polilla dorso de diamante P. xylostella es considerada el insecto plaga más importante de crucíferas a nivel mundial con un costo anual de control estimado en un billón de dólares, siendo especialmente problemática porque su control con insecticidas ha promovido la rápida evolución de resistencia en varias plagas de esta familia de plantas en el mundo (Talekar y Shelton 1993). Los programas de manejo integrado de esta especie en el ámbito mundial incluyen prácticas culturales como el riego aéreo para la interrupción de la oviposición y la destrucción de parches de plantas excesivamente infestadas, el control biológico con el uso de parasitoides y entomopatógenos, el control etológico con el uso de trampas cebadas con feromonas y el control químico con una amplia gama de insecticidas.
Como limitantes en el manejo del insecto se puede mencionar el desarrollo de la resistencia a muchos de los insecticidas comerciales disponibles, incluyendo algunos aislamientos de Bacillus thuringiensis Berliner. Esta evolución de resistencia rápida es un inconveniente significativo a las medidas químicas y microbiales actualmente usadas. Al igual que las prácticas de manejo integrado se combinan para tener un efecto final más contundente sobre la disminución poblacional de una plaga, se hace necesario rotar más ampliamente las medidas de control e incluso aumentar la variabilidad de los agentes de control biológico, con el propósito de aumentar la biodiversidad y disminuir los riesgos de la resistencia. Con esto se pretende bajar la presión de selección que ejerce un sólo método de control, aumentando la vulnerabilidad del insecto blanco por distintos frentes de ataque. Ante tal situación, se hace necesario evaluar otras alternativas de control de P. xylostella, factibles de ser incorporadas en programas de manejo integrado (Tabashnik y Mau 1986).
Los estudios adelantados por Vanderberg et al. (1998) demuestran que B. bassiana puede ser una herramienta importante en un programa de manejo de P. xylostella cuando es aplicado a plántulas en los semilleros. De este modo se pueden proveer transplantes libres de infestación, ofreciendo ventajas a los cultivadores.
La aplicación de B. bassiana induce epizootias en el campo que pueden reducir significativamente las poblaciones de la polilla dorso de diamante, afectando todos los estadios larvales, pupas y adultos (Fuentes y Carballo 1995); no obstante, la efectividad del patógeno varía entre zonas, debido principalmente a las condiciones climáticas (Lacayo 1994), por lo cual se requieren evaluaciones locales. Vanderberg et al. (1998) registran una reducción significativa de larvas de P. xylostella en L2, L3 y L4 bajo condiciones de invernadero y campo tratadas con la formulación comercial de B. bassiana denominado Mycotrol® wp, así como un efecto de persistencia con una sola aplicación hasta por dos semanas en larvas criadas en laboratorio. Es también notoria la persistencia del hongo en hojas de repollo tratadas en semillero.
Las larvas de P. xylostella infectadas con B. bassiana cesan su alimentación a las pocas horas después de expuestas, por lo cual es notoria una reducción del daño (Ibrahin y Yew 1994; Vandenberg et al. 1998). Con base en esto los dos primeros autores mencionados sugieren la posibilidad de usar Mycotrol para reforzar las prácticas actuales de manejo integrado de la polilla dorso de diamante en Malasia. Vandenberg et al. (1998) por su parte, consideran a B. bassiana como una nueva herramienta en el manejo de P. xylostella y recomiendan estudios adicionales para determinar la época más adecuada para la aplicación de este hongo, el efecto sobre otras plagas de crucíferas y sus enemigos naturales en el sistema.
Con el fin de ofrecer al agricultor alternativas biológicas enmarcadas en un programa sostenible de manejo de cultivos de crucíferas en el oriente antioqueño, se planteó este estudio cuyo objetivo fue evaluar la actividad biológica de aislamientos de B. bassiana sobre P. xylostella, tanto en condiciones de laboratorio como de campo y medir el efecto adicional de las aplicaciones del hongo sobre otros lepidópteros asociados a cultivos de coliflor.
Materiales y Métodos
La investigación se realizó en las instalaciones de CORPOICA en el Centro Administrativo "Tulio Ospina" y en el Centro de Investigación "La Selva" durante los años 2000 y 2001. Se utilizaron las facilidades de laboratorio, insectario y campo de los mencionados centros.
Manejo del material biológico
Para la cría del huésped principal, P. xylostella, se siguió la metodología de Liu y Sun (1984), modificada por Jaramillo (1999). Aunque la especie blanco en este trabajo experimental fue P. xylostella, también se evaluó el efecto de las aplicaciones de B. bassiana en campo sobre tres especies de lepidópteros más. La selección de las tres especies se realizó a partir de un estudio de campo, que incluyó la identificación de los lepidópteros asociados a crucíferas en el oriente antioqueño. Para ello se hicieron dos muestreos en el Centro de Investigación "La Selva" de CORPOICA; el primero consistió en la recolección semanal de cabezas de crucíferas (286 cabezas en total) para separar lepidópteros presentes en la etapa de cabeceo; mientras que el segundo muestreo permitió valorar la incidencia en campo de cada una de las especies, en un momento dado, sobre cultivos de repollo verde y morado (Brassica oleracea var. capitata), coliflor (Brassica oleracea var. botrytis) y brócoli (Brassica oleracea var. italica); para ello se escogieron 40 plantas al azar por material. La cría de las especies de lepidópteros se estableció en el insectario y partió inicialmente de la recolección de larvas en estados de desarrollo avanzados. Las larvas recolectadas se colocaron en bandejas plásticas y se alimentaron con hojas de repollo y coliflor de cultivos establecidos en el Centro Administrativo "Tulio Ospina", que garantizaron proveer un alimento libre de agroquímicos.
Reactivación, siembra y multiplicación de aislamientos nativos de B. bassiana
El hongo B. bassiana se obtuvo a través de las etapas de reactivación, siembra y multiplicación y correspondió a 16 aislamientos nativos de la colección de trabajo de entomopatógenos de Corpoica en Antioquia, identificados con los códigos CLS-001, 003, 005, 007, 008, 009, 010, 011, 013, 014, 015, 017, 018, 019, 020, 029 y el aislamiento de Cenicafé, Bb 9205; así mismo, los productos comerciales Bauveril, Bassiana y Bioveria.
Descripción de síntomas de P. xylostella afectado por B. bassiana
Diariamente se efectuaron observaciones de los síntomas que presentaron las larvas por los efectos del hongo tales como coloración, emisión de micelio y esporulación sobre el cuerpo. Esto permitió describir morfológicamente el proceso de micosis.
Selección de aislamientos patogénicos de B. bassiana sobre P. xylostella
La patogenicidad de los aislamientos de B. bassiana se determinó en larvas de tercer ínstar de la polilla dorso de diamante a una concentración previamente ajustada de 1x107 conidias/ml (c/ml). Para la prueba se seleccionaron larvas activas, sanas y de la misma edad para garantizar la homogeneidad del material biológico. Cada uno de los aislamientos constituyó un tratamiento. Se utilizó un control consistente de agua destilada estéril. Se inocularon 100 larvas por tratamiento para un total de 2.100. Cada tratamiento se inoculó por aspersión en volumen de 5 ml a la concentración descrita anteriormente. Las larvas se colocaron en el interior de una jaula de madera y tela tull, sobre hojas de repollo, dentro de un recipiente plástico cubierto con tela organza, ubicado en el fondo de la jaula. Los datos de mortalidad se tomaron diariamente después de 48 horas de la aplicación de la suspensión hasta el día décimo.
Las características de escogencia de un entomopatógeno deben ser varias y se deben conjugar de tal manera que permitan la comparación de varios aislamientos. De este modo se le da la oportunidad a cada uno de mostrar sus habilidades al colonizar el hospedero y al investigador para tener más éxito en la escogencia. La selección de los tres aislamientos más patogénicos contra P. xylostella se hizo considerando el porcentaje de mortalidad, el tiempo letal medio (TL50) y la mortalidad intrínseca, entendida esta última como la proporción de insectos muertos con el hongo B. bassiana con respecto a la mortalidad total. Con el fin de escoger los dos aislamientos nativos mejores, por su habilidad patogénica sobre L3 de P. xylostella, se hizo una comparación entre los cuatro más sobresalientes, asignando rangos entre ellos para cada una de las variables. Los rangos más bajos fueron asignados a las respuestas más pobres o no deseadas como porcentaje de mortalidad pequeños o nulos, proporción de muertos por B. bassiana pequeña o nula y tiempos letales medios muy tardíos.
Producción promedia de conidias por larva
Para cada uno de los aislamientos de B. bassiana se determinó la cantidad promedio de conidias producidas por una larva de P. xylostella infectada. De cada aislamiento se escogieron cinco larvas con signos de B. bassiana, las cuales se dejaron en cámara húmeda para su esporulación hasta el noveno día después de la aparición de los primeros síntomas de infección. Las cinco larvas se pesaron en una balanza analítica; posteriormente se agregaron 10 ml de agua destilada estéril y se adicionó el dispersante Agral 90 al 2 por mil. La solución así constituida se centrifugó durante 3 minutos a 1.000 rpm para desprender y dispersar adecuadamente las conidias. La concentración de conidias de dicha solución se calculó de una dilución 10−3 mediante conteos de células en la cámara de Neubauer.
Determinación de las concentraciones letales CL50 y CL90 para los aislamientos más activos contra P. xylostella
El bioensayo se llevó a cabo en insectario. Se evaluaron las concentraciones 103, 105, 107, 109 c/ml y un testigo con sólo agua destilada estéril. Cada concentración se inoculó directamente sobre 100 larvas ubicadas en hojas de repollo, dispuestas en bandejas plásticas. Se trabajó con los tres tratamientos que mostraron mayor actividad biológica sobre P. xylostella. Los datos de mortalidad se tomaron 48 horas después de la aplicación de la suspensión de esporas, hasta el octavo día; con ellos se calcularon las concentraciones letales media (CL50) y noventa (CL90) usando el análisis Probit.
Pruebas de campo
Se realizaron con dos aislamientos seleccionados a las siguientes concentraciones: CLS 029 a 1.6 x 107 c/ml: CLS 003 a 1.3 x 108 c/ml y, el producto comercial Bauveril a 1x108 c/ml, sobre las especies P. xylostella (especie blanco), Copitarsia sp., Peridroma sp. y Leptophobia aripa. Estas concentraciones se establecieron teniendo en cuenta las concentraciones letales calculadas en el paso anterior y la capacidad máxima del aislamiento para producir conidias en arroz, bajo las condiciones propias de este experimento. La variable medida fue la mortalidad intrínseca, es decir la causada por el microorganismo aplicado: B. bassiana.
El montaje del ensayo se realizó sobre cultivos comerciales de coliflor de la variedad Casablanca mejorada, ubicados en el Centro de Investigación "La Selva" en Rionegro, Antioquia. Se utilizó el diseño de bloques completos al azar con cinco tratamientos y cuatro repeticiones. Los bloques fueron eras de 1,2 m de ancho y el tamaño de las unidades experimentales estuvo dado por una era de 1,2 m de ancho por 5 de largo, para un área de 6 m2. Cada parcela se separó de la contigua con una barrera de maíz. Las plantas de cada parcela se infestaron una sola vez con larvas de los insectos P. xylostella, L. aripa, Copitarsia sp. y Peridroma sp.; dicha infestación se hizo en la semana seis del cultivo para L. aripa y al inicio de la formación de pella, cuando el florete tenía 1-5 cm de diámetro, para las tres especies restantes.
Se infestaron 5 plantas con larvas de las especies L. aripa y Peridroma sp. y 10 con larvas de P. xylostella y Copitarsia sp.; el número de larvas por especie colocadas por planta fue de 3, 1, 10 y 1, respectivamente y el total de cada especie mencionada por tratamiento fue de 400, 60, 40 y 20. La definición de la cantidad de larvas por especie insectil tuvo que ver con la capacidad de cría en el insectario, con la cantidad de plantas por parcela y con los hábitos de cada insecto, especialmente de Copitarsia el cual presenta canibalismo. La metodología de infestación consistió en colocar las larvas en el cogollo de la planta en la etapa de cabeceo, a excepción de L. aripa que se ubicó en el follaje. La inoculación de los tratamientos se hizo después que las larvas se desplazaron a los sitios preferidos de alimentación en la planta (1-2 horas después de infestación). Las plantas infestadas se señalaron con cintas de color para discriminar cada especie insectil. Cada aislamiento de B. bassiana constituyó un tratamiento el cual fue inoculado tres veces, dos de ellos antes de la formación de cabeza y uno en la etapa de cabeceo. Cada tratamiento se inoculó en todas las plantas de cada parcela, cubriendo con la aspersión la totalidad de la planta infestada.
Los tratamientos fueron los aislamientos CLS 029, CLS 003, el producto comercial Bauveril y dos testigos, uno dentro del lote contiguo a los tratamientos de B. bassiana y otro en un lote aparte, lejos de la influencia de los mismos. Con esto se pretendió observar la posibilidad de infestación del hongo por vecindad a los lotes tratados y sus facilidades de dispersión en el ambiente de prueba. Los tratamientos se aplicaron manualmente, empleando bombas de espalda. Las evaluaciones se hicieron 5 días después de inocular el hongo y al momento de la cosecha.
Las variables medidas durante la etapa de campo fueron: número de larvas afectadas por B. bassiana para cada especie de lepidóptero por parcela; número de larvas de cada especie de lepidóptero afectadas por otros factores bióticos de mortalidad; número de cabezas dañadas por la especie plaga o por la población mixta (dos o más especies) y número de cabezas sin daño de lepidópteros.
Resultados y Discusión
Las cuatro especies de lepidópteros de mayor incidencia en cultivos de crucíferas en el oriente antioqueño fueron P. xylostella, Leptophobia aripa, Copitarsia sp. y Peridroma sp. (Fig. 1). Las especies P. xylostella, Copitarsia sp. y Peridroma sp. causan daños que afectan la calidad y suscitan rechazos en el mercado de repollo, coliflor y brócoli principalmente. El porcentaje de incidencia en campo de los noctuídos Copitarsia sp. y Peridroma sp. fue respectivamente de 28 y 25% en coliflor, 13 y 33% en repollo verde, 33 y 10% en repollo morado.
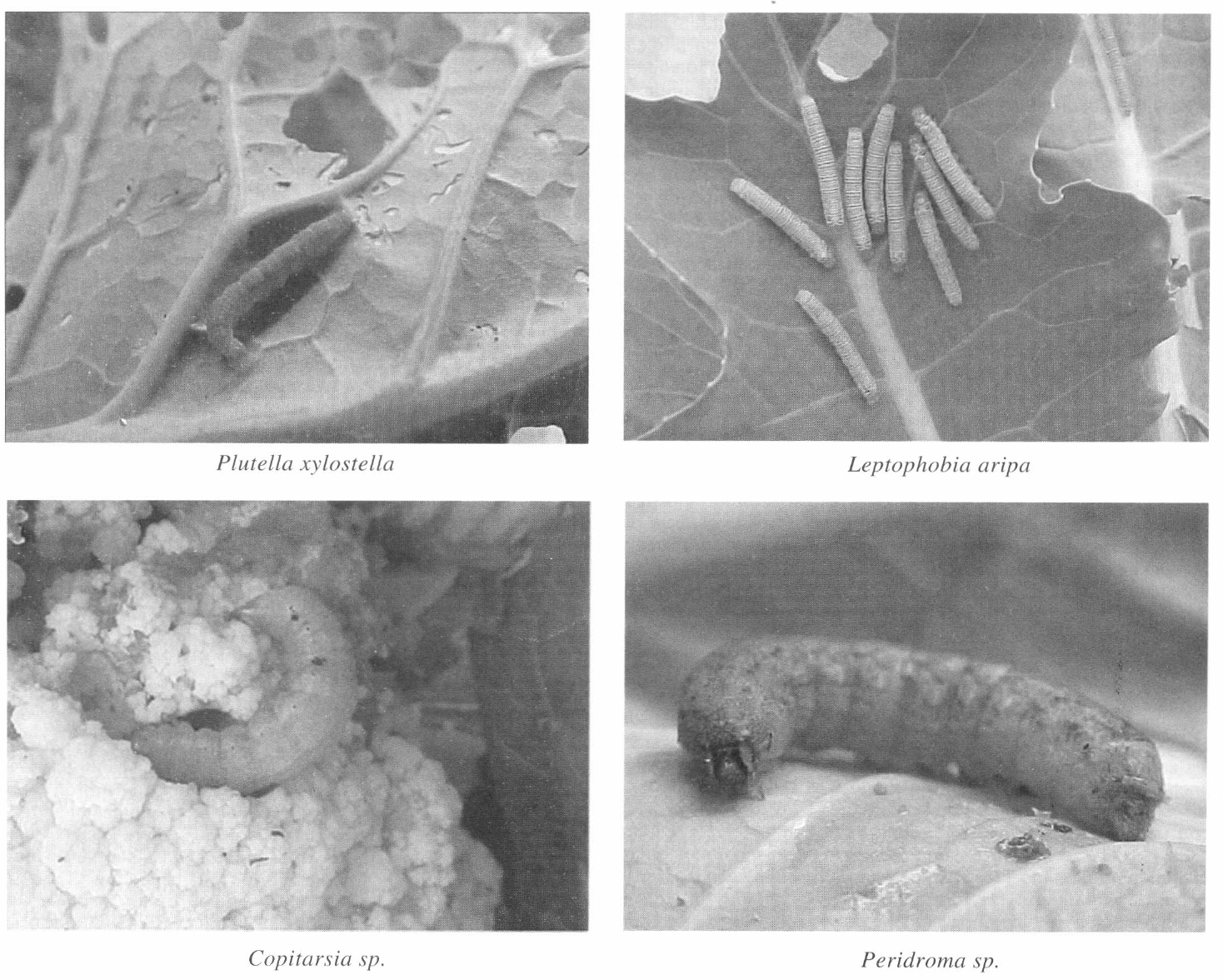
Larvas de las cuatro especies de lepidópteros consideradas en el trabajo experimental.
Sintomatología de larvas y signos del hongo
Todos los aislamientos que causaron mortalidad manifestaron signos típicos de B. bassiana en larvas de P. xylostella. Cuando los insectos se mueren por la acción del hongo quedan adheridos al follaje; posteriormente presentan micelio y esporas. Esta característica es resaltada por Vahos et al. (1998), trabajando con Collaria sp. pos. columbiensis, quienes observaron esporulación de B. bassiana en insectos tratados, desde unos pocos días hasta veinte días después. Esto además fue considerado por ellos como una característica a destacar, ya que de presentarse esporulación en condiciones de campo, los insectos muertos por el hongo aumentarían el inóculo inicial aplicado, convirtiéndose en multiplicadores del microorganismo.
La sintomatología general presentada por las larvas de Plutella enfermas (Fig. 2) se manifestó como una reducción de la alimentación después del tercer día de inoculado el hongo. También fue evidente un cambio en la coloración de verde intenso (que es el normal) a amarillo verdoso próximas a su muerte, luego se tornan de color anaranjado opaco muy característico en larvas endurecidas. Posteriormente se exterioriza el desarrollo micelial que se inicia desde las primeras suturas intersegmentales de la larva momificada, hasta aparecer en todas éstas, incluso en la región anal. Se nota inicialmente un cubrimiento parcial de los anillos de la larva llegando a quedar cubiertos totalmente por el hongo, con la subsecuente esporulación sobre la larva colonizada. Los síntomas y signos de B. bassiana en larvas de P. xylostella se presentaron desde el tercer día después del tratamiento. La frecuencia mayor de larvas con signos de B. bassiana se vio entre los días 5° al 8° post tratamiento.

Síntomas, emisión de micelio y esporulación de B. bassiana en larvas de P. xylostella.
Patogenicidad de aislamientos de B. bassiana sobre P. xylostella
La patogenicidad de los diferentes tratamientos que contenían B. bassiana se evaluó teniendo en cuenta tres características: el porcentaje de mortalidad, el tiempo letal medio (TL50) y la mortalidad intrínseca o proporción de larvas con signos del entomopatógeno. Los datos de mortalidad de larvas de P. xylostella corregidos a través de la fórmula de Schneider y Orelly, citado por Ciba Geigy (1981), para los diferentes aislamientos de B. bassiana se presentan en la tabla 1. Las mortalidades variaron entre el 26 y 98%. Los insectos en los tratamientos testigos registraron mortalidades muy bajas con porcentajes inferiores al 10%, lo cual sugiere la efectividad de los tratamientos. El aislamiento que causó la mortalidad mayor fue CLS 029, con 97,83%, seguido de los aislamientos CLS 003, CLS 018, CLS 014 y el producto comercial Bauveril con 82,61, 71,74, 65,22 y 63,04%, respectivamente. Horie et al. (1981) y Prior (1991), citados por López (1994), señalan que cuando un aislamiento proviene del mismo insecto sobre el cual se desea conocer su efecto, es más patogénico que los aislamientos de hongos obtenidos de insectos diferentes, ya que puede existir cierta especificidad entre el hongo y su huésped. Esta puede ser la razón por la cual el aislamiento CLS 029 presentó mortalidad mayor sobre la polilla, ya que procedía de larvas de campo afectadas por el hongo. Con esta variable se tiene la primera aproximación al conocimiento de un grupo de aislados de B. bassiana, en relación con su habilidad para controlar Plutella xylostella en Colombia. Es de anotar que el producto comercial Bassiana no mostró actividad biológica contra larvas de la polilla dorso de diamante. Sólo cinco de los aislamientos superaron el 60% de mortalidad sobre larvas de la polilla: CLS 029, CLS 003, CLS 018, CLS 014 y Bauveril.
Mortalidad causada por 20 aislamientos de B. bassiana sobre larvas de P. xylostella. Datos a los 10 días después del tratamiento
Otro criterio utilizado para seleccionar las cepas fue el tiempo letal medio (Tabla 2), calculado con el tiempo de la mortalidad total mediante análisis de regresión Probit. Los tiempos letales medios fluctuaron entre 6 y 13 días para los tratamientos en prueba. Los aislamientos con menor tiempo letal medio fueron el CLS 003 y CLS 029 con 6,1 y 6,7 días, respectivamente. Los aislamientos que no consiguieron matar el 50% de la población durante los 10 días post inoculación presentaron tiempos letales medios elevados, los cuales se estimaron con el programa estadístico usado.
Tiempo letal medio (TL50) de 20 aislamientos de B. bassiana inoculados sobre larvas de P. xylostella, a una concentración de 1x107 conidias/ml
La utilización de la concentración 1x107 c/ml permitió separar los aislados por su virulencia (capacidad del microorganismo para invadir, multiplicarse y causar mortalidad del huésped). Esto corrobora lo observado por Fuentes y Carballo (1995), quienes encontraron que B. bassiana evaluado sobre P. xylostella a la concentración 1x107 c/ml, facilitó la separación entre aislamientos, en comparación con concentraciones mayores, ya que de esta manera se puede cuantificar mejor la patogenicidad y virulencia. Esto les permitió además corroborar que la virulencia era una característica de cada aislamiento.
En el tercer criterio utilizado para seleccionar los aislamientos mejores, que tuvo que ver con la mortalidad intrínseca al microorganismo aplicado, pudo observarse que la mayoría de los tratamientos probados manifestaron signos del hongo a la concentración 1x107 c/ml. Sin desconocer que la mortalidad causada por un microorganismo lleva un componente bioquímico consistente de la micotoxina, también responsable de esa muerte, es muy importante para un agente de control biológico de insectos de tipo microbial, causar una alta proporción de muertes con signos del entomopatógeno; es decir, con la posibilidad de multiplicarse sobre su hospedero, aumentando el potencial de inóculo. Esto además facilitaría las posibilidades de permanencia y diseminación en programas de control en donde se le incluya. De otro lado, mirándolo desde el punto de vista de la aceptación del usuario, es más convincente un producto biológico cuando logra producir síntomas y signos sobre la población tratada.
Los aislamientos que permitieron una mayor expresión de síntomas y signos del hongo B. bassiana fueron CLS 029, CLS 003, CLS 018, CLS 014 y Bauveril. Estos mismos tratamientos fueron los que presentaron la mayor proporción entre la mortalidad intrínseca y la mortalidad total, estando muy cerca de la unidad. Esto quiere decir que del total de larvas muertas por tratamiento el 91, 90, 89, 88 y 87% se debieron a B. bassiana, en los tratamientos CLS 029, CLS 018, CLS 003, CLS 014 y Bauveril, respectivamente. Esta relación tan estrecha entre estas dos variables es un buen indicativo de la capacidad patogénica de los tratamientos mencionados y muestra una vez más al aislamiento CLS 029 con los valores mayores; dicho aislamiento no sólo está relacionado con la especie blanco, por haber sido aislado de ella, sino que también se presume que puede estar adaptado al medio y tener más habilidades para sobrevivir en él.
Vale la pena destacar la acción del producto comercial Bauveril, el cual causó mortalidades del 63% sobre larvas de la polilla dorso de diamante; requiere de 7,32 días para causar el 50% de mortalidad, lo que lo ubica en el tercer lugar entre los tratamientos comparados y el 87% de las larvas muertas presentan colonización y esporulación de B. bassiana. Este conjunto de características que lo acercan mucho en su habilidad patogénica a los aislamientos nativos más destacados, lo distingue a su vez de los demás productos comerciales evaluados, los cuales tuvieron valoraciones pobres en sus parámetros. Dado que Bauveril está ya formulado, lo posiciona como una opción biológica disponible. Faltaría entonces evaluar su comportamiento en condiciones de campo, por lo cual este tratamiento fue seleccionado para las etapas posteriores de esta investigación que condujeron finalmente a las evaluaciones de campo de los tratamientos seleccionados por virulencia.
Al escoger los dos aislamientos nativos mejores por su habilidad patogénica sobre L3 de P. xylostella utilizando para ello la conjugación de los tres criterios de selección propuestos, puede verse más claramente cómo el aislamiento CLS 029 queda por encima de los demás, ya que se destacó en las tres características comparadas, seguido por CLS 003 (Tabla 3). Nótese como este último aislamiento, a pesar de tener un TL50 menor y ser ésta una característica interesante para varios investigadores, fue superado por CLS 029, cuyo TL50 es un poco mayor. Este es uno de esos casos en los que la escogencia se dificulta por la similitud en su actividad biológica, ya que siendo ambos muy buenos para causar mortalidad sobre P. xylostella se deben ordenar de mayor a menor. Es por tanto interesante observar como las otras dos variables; mortalidad corregida y proporción de muertes por B. bassiana permitieron ordenarlos en igualdad de condiciones. Los aislamientos CLS 018 y CLS 014 también tienen una gran habilidad patogénica sobre esta plaga. Aunque en esta investigación no clasificaron para ser evaluados en condiciones de campo, valdría la pena tenerlos en cuenta para futuras investigaciones donde se evalúen otras variables como resistencia a la radiación ultravioleta, persistencia en el campo, capacidad de diseminación y eficiencia en el control de otras especies plagas asociadas a las hortalizas, entre otras.
Actividad biológica sobre P. xylostella de cuatro aislamientos nativos sobresalientes
Deseable: Porcentaje alto
Ri: Rango asignado de 1 a 4
Deseable: Tiempo bajo
Proporción Mortalidad por B. bassiana sobre mortalidad total.
El tercer aislamiento escogido para trabajar en campo fue Bauveril, por causar la mayor mortalidad entre los tres productos comerciales. Se consideró que, de presentar buen comportamiento al evaluarlo en campo, se podría contar con una fuente de B. bassiana de fácil consecución en el mercado. Además, este producto causó un porcentaje de mortalidad superior al 60% y tuvo un tiempo letal medio bajo.
Producción de conidias por larva
De los tratamientos de B. bassiana evaluados, 19 mostraron patogenicidad contra larvas de P. xylostella, produciendo micelio y conidias cuando se inocularon en larvas de tercer ínstar. Los promedios de conidias producidas por larva con cada uno de los aislamientos probados (Tabla 4) manifestaron variación desde 0,08 hasta 3,9x106 conidias, como ocurrió con los aislamientos CLS 007 y CLS 029, respectivamente; esto último indica que entre ellos existió una diferencia de 100 veces en cuanto a producción de conidias por larva se refiere. Los aislamientos CLS 029, CLS 019, CLS 020 y CLS 017 produjeron más cantidad de conidias por larva, 3,92x106, 3,61x106, 1,69x106 y 1,47x106, respectivamente; sin embargo, los tres últimos tienen porcentajes bajos de mortalidad.
Número promedio de conidias producidas por larva de P. xylostella, inoculadas con 19 tratamientos a base de B. bassiana a una concentración 1x107 conidias/ml.
La producción más baja de conidias por larva se detectó con el aislamiento CLS 007 con 0.08x106, con el cual a su vez se presentó un peso bajo de larvas; adicionalmente, se observó un volumen bajo de micelio, interpretado por el diámetro pequeño de la colonia. Sin embargo, larvas afectadas por los aislamientos CLS 005 y CLS 011 con el mismo peso que CLS 007, tuvieron 10 veces más producción de conidias/larva que este último. Las larvas afectadas por los aislamientos con mayor producción de conidias tuvieron pesos intermedios de acuerdo con la respuesta de los 19 tratamientos que aparecen en la tabla 4. Se aprecia que las larvas afectadas por los aislamientos CLS 003, CLS 010 y Cenicafé Bb 9205, las cuales tuvieron los pesos mayores, produjeron 10 veces menos conidias/larva que los aislamientos CLS 029, CLS 019, CLS 020 y CLS 017. Al parecer, estas dos variables no están muy relacionadas y la producción de conidias por larva parece más bien ser una característica propia del aislamiento.
Fuentes y Carballo (1995) evaluaron 10 aislamientos de B. bassiana sobre P. xylostella y aunque los aislamientos utilizados por ellos fueron diferentes a los utilizados en el presente trabajo, las relaciones que hacen dichos autores entre producción de conidias por larva y porcentaje de mortalidad son interesantes y se asemejan mucho a las obtenidas con los tratamientos probados. El aislamiento CLS 019 en este trabajo produjo 36,1x105 conidias por larva y sin embargo su porcentaje de mortalidad no llega al 45%; esta observación también se aprecia con el aislamiento A2 indicado por Fuentes y Carballo, con una producción alta de conidias por larva y porcentaje bajo de mortalidad. De otro lado, se observó que el aislamiento CLS 003 evaluado en la presente investigación y que produjo 0,1x106 conidias/larva causó mortalidad superior al 80%. Los mismos autores relacionados arriba también señalan que los aislamientos Achi 5, 167, A4, Achi 1 y A7 tuvieron producción baja de conidias por larva (4.1, 5.7, 2.54, 2.37 y 4.7x105, respectivamente) y sin embargo, causaron mortalidades altas. Estos autores consideran que la efectividad de un aislamiento no necesariamente está relacionada con la producción de conidias, sino más bien con su virulencia y viabilidad; aunque ellos consideran también que la efectividad tiene mucho que ver con la capacidad de diseminación y que ésta está favorecida por una producción alta de conidias por hospedero. Con base en los resultados obtenidos por Fuentes y Carballo (1995), se deduce que los genotipos más virulentos tienen por lo regular una relación más estrecha con la capacidad de producir mortalidad alta, que con producir grandes cantidades de conidias sobre el hospedero colonizado. En cambio, la producción alta de conidias y el porcentaje de mortalidad bajo, que puede deberse a pérdidas de virulencia, predomina en los genotipos poco virulentos o de baja patogenicidad sobre el hospedero.
Concentración letal media (CL50)
La concentración letal media de los aislamientos nativos CLS 029, CLS 003 y del producto comercial Bauveril se presentan en la tabla 5. El mismo análisis Probit para la CL50 permitió obtener los valores de la CL90 necesarios para realizar las pruebas de campo. Robert y Marchal (1980), ensayando B. bassiana sobre P. xylostella encontraron una CL50 de 1.5x107 c/ml, usando el método de aspersión de las conidias en discos de hojas. Dicha CL50 es aproximadamente 100 veces superior a la requerida por CLS 029 sobre la misma plaga, lo que indica una actividad mayor de este último aislamiento. Sobre este mismo insecto, Ignoffo et al. (1979) evaluaron el micro insecticida Boverin utilizando el método anterior y encontraron una CL50 de 2,7x108 c/ml, mucho menos activa que la CLS 029 del presente experimento. Fuentes y Carballo (1995) encontraron con el aislamiento 447 que la concentración para matar el 50% de la población de larvas de Plutella fue de 2,2x105 y para matar el 95% de 5,1x107 c/ml. Estos autores también observaron que al aumentar la concentración se produce un incremento en la mortalidad. De igual manera Gutiérrez (1991), con el aislamiento 117 de B. bassiana encontró una CL50 de 4,16x105 c/ml y que al aumentar la concentración aumentaba la mortalidad de Plutella. Los resultados logrados por estos dos últimos autores se asemejan a los obtenidos en el presente trabajo experimental con el aislamiento CLS 029.
Valores de las CL50 y CL90 determinados mediante análisis Probit
Los resultados obtenidos con el aislamiento CLS 029 y los registrados con el 447 de Fuentes y Carballo (1995) y el 117 de Gutiérrez (1991), muestran una concentración letal media baja, lo que permite utilizar menor cantidad de conidias, obtener mortalidad alta y garantizar mayor seguridad para organismos no blanco.
Efecto de los aislamientos CLS 029, CLS 003 y el producto comercial Bauveril sobre P. xylostella y otros lepidópteros asociados a la producción de coliflor en condiciones de campo
Lepidopteros afectados por B. bassiana
Sólo las larvas de tres especies se encontraron con signos del hongo. No se detectó ningún otro estado de desarrollo con síntomas y/o signos de B. bassiana. Sin embargo, en el insectario durante las pruebas de patogenicidad, se observaron pupas de P. xylostella afectadas por el entomopatógeno.
La mortalidad intrínseca observada en los tratamientos CLS 029, CLS 003 y Bauveril sobre xylostella bajo condiciones de campo, fue de 72,7, 69,3 y 32,4%, respectivamente. En condiciones de insectario, con los mismos tratamientos dicha mortalidad fue de 97,83, 82,61 y 63,04% como se pudo ver cuando se analizó la patogenicidad. Al comparar la mortalidad intrínseca bajo estas dos condiciones se observó que en el campo fue más baja. Esto posiblemente se deba a que la aspersión del entomopatógeno en la última condición se hizo sobre las plantas después de infestar con las larvas; mientras que en el insectario el inóculo se asperjó directamente sobre las larvas, dándole quizá en esta última condición más oportunidad al entomopatógeno de hacer un contacto mejor con el hospedero.
Al aplicar la prueba de rangos múltiples de Friedman para la mortalidad intrínseca (Tabla 6) y hacer comparaciones entre tratamientos utilizando los valores críticos de Tukey, solamente se encontró diferencia de los tratamientos CLS 029 y CLS 003 con el testigo Cancha (a = 0.05), es decir, el testigo que se sembró aislado donde no hubo presión de inóculo de B. bassiana. Se detectó que el número de larvas afectadas por B. bassiana fue estadísticamente igual cuando se aplicaron tratamientos que tenían dicho entomopatógeno (CLS 029, CLS 003 y Bauveril). Es interesante anotar cómo el tratamiento testigo vega, que en el sorteo al azar quedó dentro del bloque, también presentó larvas afectadas por B. bassiana; razón por la cual la prueba no detecta diferencias de este testigo con los demás tratamientos que llevaron el entomopatógeno. Esto está indicando la capacidad que tiene el hongo de infectar larvas de parcelas contiguas o vecinas. Es posible que las larvas del tratamiento testigo vega se infectaran por presión de inóculo de alguno de los aislamientos vecinos. No obstante, existe la posibilidad de que el hongo aparecido en el testigo. sea nativo y estuviera presente en el vegetal o en larvas de ocurrencia natural en la parcela.
Mortalidad intrínseca de larvas de P. xylostella inoculadas con tratamientos de B. bassiana en condiciones de campo. Rionegro, Antioquia, noviembre de 2000
Xβr corregido = 15.01; Χβ α 0.05 = 9.49; Ri= calculados mediante prueba Friedman; W = 12.2; d= diferencia entre tratamientos vecinos.
El hecho de presentarse larvas afectadas en el testigo (sin inocular B, bassiana) ubicado cerca a parcelas inoculadas, provee información para plantear investigaciones, quizá inoculando este entomopatógeno en fajas y evaluando factores que intervienen en su dispersión a fajas no tratadas,
Al analizar el comportamiento de L. aripa post aplicación de B. bassiana, se encontró que el número de larvas afectadas no mostró diferencia estadística entre los tratamientos, La prueba de rangos múltiples de Friedman comprueba esto (Tabla 7). Además se puede observar que aunque hubo larvas afectadas por B. bassiana en el tratamiento CLS 029 éste no difiere estadísticamente del testigo, Sólo el tratamiento CLS 029 causó mortalidad comprobable por el entomopatógeno, mientras que CLS 003 y Bauveril fueron completamente inocuos a la población de larvas expuestas, en cuanto al entorno patógeno se refiere.
Mortalidad intrínseca de larvas de L. aripa inoculadas con aislamientos nativos de B. bassiana. Rionegro, Antioquia, noviembre 2000
X2r corregido = 16; X2 u 0.05 = 9.49; Ri= calculados mediante prueba Friedman; W = 12.2, d= diferencia entre tratamientos vecinos
Los registros de mortalidad de larvas de Copitarsia sp. por efecto de los tratamientos se presentan en la tabla 8. Aplicando la prueba de rangos múltiples de Friedman no se encontró diferencia estadística entre tratamientos con y sin B, bassiana. Sólo una larva de Copitarsia apareció con signos de Beauveria y fue encontrada en una parcela tratada con el aislamiento CLS 003. Sin embargo, no se puede asegurar que fue dicho aislamiento el causante de la mortalidad, Pudo haber sido un aislamiento nativo que afectó a esta especie, De todas maneras, aunque sólo una larva fue afectada por B. bassiana se considera importante el hallazgo. ya que con este aislamiento se puede presionar sobre el hospedero para potenciar la capacidad del hongo mediante selección. Esto es. aislar el hongo encontrado y pasarlo sucesivamente por larvas de Copitarsia, reaislando e inoculando nuevamente cada vez que se presente la infección. hasta conseguir un porcentaje de mortalidad aceptable.
Mortalidad intrínseca de larvas de Copitarsia sp. inoculadas con aislamientos nativos de B, oassiana. Rionegro, Anlioquia. noviembre 2000
X2r corregido = 0,62; X2 a 0.05 = 9,49
En el ensayo de campo no se presentaron larvas de Peridroma sp. afectadas por B. bassiana y estadísticamente no hubo diferencia entre tratamientos con y sin el entomopatógeno. Todos los tratamientos de B. bassiana fueron completamente inocuos a la población de larvas de Peridroma sp. expuesta.
Cabezas de coliflor dañadas por especie plaga o por población mixta (dos o más especies)
En la tabla 9 se presentan los valores del número de cabezas de coliflor dañadas por tratamiento (transformados a arcoseno √x). Los resultados indicaron que el porcentaje menor de cabezas con daño se consiguió en el tratamiento CLS 029 con 38.46%. el cual difiere estadísticamente de todos los tratamientos (a = 0.01). El segundo porcentaje de cabezas dañadas se obtuvo en el tratamiento CLS 003 con un 54%. el cual se asemeja a Bauveril donde se presentó un 63.4% de daño. Los testigos Vega y Cancha se igualaron en un porcentaje de cabezas dañadas con 73,1 y 72.6. respectivamente. Esto indica que al no aplicar un tratamiento que proteja del daño de lepidópteros. se presentará un porcentaje alto de pérdidas. Estas cabezas con daño son castigadas en el mercado y tienen que venderse como de segunda o pasan a ser de rechazo.
Número de cabezas de coliflor dañadas por parcela al momento de la cosecha. Datos transformados a arcoseno √x
Promedios con la misma letra no difieren entre sí al 0,01 según prueba Duncan.
Conclusiones
Bajo las condiciones metodológicas utilizadas en este trabajo experimental, se puede concluir lo siguiente:
B. bassiana es un hongo con grandes opciones como controlador biológico de Plutella xylostella, dado el abundante número de aislamientos a los cuales es susceptible. Los tres tratamientos seleccionados en condiciones de insectario tuvieron una acción patogénica similar en condiciones de campo, lo que sugiere que las fuentes de B. bassiana tienen una gran habilidad para infectar a P. xylostella.
Los tratamientos con mayor actividad biológica sobre P. xylostella fueron en su orden los aislamientos CLS 029, CLS 003 y el producto comercial Bauveril, con concentraciones letales medias que fluctuaron entre 1x105 y 1x106 c/ml. Estos tres tratamientos evaluados en condiciones de campo, produjeron micelio y esporularon sobre larvas de P. xylostella, hecho que permite una mayor permanencia y facilita las posibilidades de diseminación del microorganismo bajo estas condiciones. El buen comportamiento del aislamiento CLS 029 lo perfila como el más opcionado para incluir como componente biológico en un programa de manejo integrado de la polilla dorso de diamante. Este aislamiento, a una concentración de 1x107 c/ml, permite una reducción poblacional de larvas de tercer ínstar de P. xylostella entre los 7 y 10 días post tratamiento. Su habilidad patogénica adicional sobre L. aripa, lo habilita para ser utilizado en el manejo de ambas especies de lepidópteros desde las primeras etapas de desarrollo en cultivos de crucíferas.
Ninguno de los tratamientos de B. bassiana evaluados en condiciones de campo, mostró habilidad patogénica sobre los noctuídos Copitarsia y Peridroma, lo cual amerita buscar otras alternativas para manejarlos en cultivos de crucíferas.
Agradecimientos
Los autores agradecen a los profesores Alejandro Madrigal C., Ingeniero Agrónomo, Especialista en Control Biológico y Sandra Inés Uribe Soto, Ingeniera Agrónomo, Ph.D., por las sugerencias al manuscrito. Al señor Ovidio Montoya Herrera, Auxiliar del Laboratorio de Entomología de CORPOICA. A la Universidad de Córdoba, la Asociación Hortifrutícola de Colombia, la Corporación Colombiana de Investigación Agropecuaria, CORPOICA, por cofinanciar la investigación.