
Abstract
Summary
A vector-borne disease model has been developed through system dynamics to represent the entomological, epidemiological and climatic interactions of malaria transmission conductive to disease outbreaks in Nuquí prone-region, Chocó province, along the Pacific coast. The entomological exogenous variables considered relevant for malaria incidence have been collected from laboratory experiments and field data (indoor and outdoor captures). The parasite transmission between linking mosquitoes and vertebrate host populations during a blood meal has been represented using three infectious disease models: the Entomological Inoculation Rate, the Basic Reproduction Rate and the Vectorial Capacity for diverse vector density scenarios. Considering a breeding place availability model and several predator-prey-food models allow us to represent the vectorial densities fluctuations observed during the fields campaigns. The comprehensive model has been applied to represent malaria incidence during the period Nov/1997-Feb/2001 (40 months, 1200 days simulation period), when both El Nino and La Nina events strongly affected the hydro-climatology of Colombia. The model has been run for observed climatic patterns such as mean daily temperatures, total daily precipitation records, and mean daily relative humidities gathered by a nearby climatological station. Diverse temperature scenarios have been considered to deepen the understanding of the entomological-climatic linkages conductive to malaria outbreaks. Sensitivity analysis and instabilities cases have been also studied during the experimentation-validation processes. Obtained results allow us to conclude that the model constitutes a promising tool to deepen the understanding of the ecological, entomological, and epidemiological linkages conductive to malaria outbreaks.
Introducción
La atención de recientes trabajos de investigación en hidroclimatología ha estado orientada al análisis de los posibles impactos en la salud humana de los cambios atmosféricos globales (Bouma y Van Der Kaay 1994; Bouma y Dye 1997; Martens et al. 1995a, 1995b, 1997a, 1997b; Martens 1997b, 1998; Patz 1998; Patz et al. 1998; Poveda y Rojas 1996, 1997; Poveda et al. 1999, 2001). Como posibles efectos indirectos mediados ecológicamente se consideran las enfermedades transmitidas por vectores biológicos, las enfermedades generadas por degradación de los cuerpos de agua y las deficiencias asociadas al agotamiento y deterioro de los cultivos de alimentos. La alteración de las incidencias, emergencias y distribuciones de enfermedades transmitidas por vectores biológicos, principales causas de morbilidad y mortalidad en regiones tropicales, parece responder a cambios en la temperatura y la precipitación que influencian el comportamiento (frecuencia de alimentación sanguínea) y la distribución geográfica de vectores (dispersión en áreas que restringían su desarrollo), así como favorecen la incubación de los parásitos dentro de los mismos (en tasas de reproducción y maduración). Las enfermedades infecciosas transmitidas por mosquitos vectores (malaria, dengue, fiebre amarilla y algunos casos de encefalitis), son consideradas las más afectadas debido al cambio climático global.
La malaria, enfermedad infecciosa con mayor prevalencia en regiones tropicales, le quita la vida a 3.000 individuos diariamente, en su gran mayoría niños (Epstein 2000). Algunos modelos proyectan que para el final del siglo XXI, el calentamiento atmosférico global extenderá la zona de transmisión potencial de la enfermedad de un 45% de la población mundial a un área con un 60% de la misma (Martens 1997). Aunque esta expansión se ajusta ciertamente a las predicciones, la causa de tal desarrollo no puede estar estrictamente asociada al cambio climático global. Otros factores pueden estar también relacionados, tales como el deterioro del medio ambiente -favorece la proliferación de la población de mosquitos-, la suspensión o disminución de las medidas de control de mosquitos y otros programas de salud pública, y el incremento en las resistencias a la droga y el uso de pesticidas. Sin embargo, la hipótesis del cambio climático global y su contribución a la transmisión de las enfermedades infecciosas, ha adquirido aún más fuerza.
Ahora, si se desarrollan los mecanismos necesarios para entender las relaciones clima-enfermedades infecciosas, vía profundización en las interacciones entomológicas-climáticas de la transmisión, es posible diseñar campañas futuras de control y erradicación soportadas en señales de alerta temprana. Es así como el principal objetivo de este trabajo consistió en desarrollar un modelo matemático en dinámica de sistemas para representar la transmisión de la malaria y tratar de entender las relaciones existentes entre los brotes epidémicos y los patrones climáticos (haciendo particular énfasis en la ocurrencia de la fase cálida del evento, el Niño-Oscilación del Sur).
Los modelos abstractos ya han suscitado interés a nivel mundial, resaltándose los trabajos publicados por Aracil y Toro (1988, 1993), Aron y May (1982), Blower et al. (1996, 1998), Blower y Gerberding (1998), Dietz (1988), Dyner (1993), Garrett-Jones (1964), Haile (1989), McKenzie et al. (1998), MacDonald (1957) y Martens (1997). Teniendo como base las fortalezas y debilidades de los modelos mencionados, el trabajo que se ha desarrollado ha concentrado sus esfuerzos en torno a los siguientes puntos fundamentales: (1) profundización en el ciclo de transmisión de la enfermedad, con el fin de definir los estados más relevantes y las variables endógenas que son altamente influenciadas por variables climáticas; (2) profundización en la ecología del vector, sus patrones de comportamiento, hábitos alimenticios y variables entomológicas relevantes, haciendo especial énfasis en sus relaciones con el clima; (3) análisis de viabilidad de hábitat de reproducción para el desarrollo de la población de mosquitos en sus estados preimaginales; (4) representación de interacciones de las larvas del vector con sus predadores naturales; (5) recopilación de información entomológica de campañas de colecta de adultos y bioensayos, e información epidemiológica levantada en el horizonte de tiempo seleccionado; (6) análisis de escenarios alternos que consideren variaciones en densidad vectorial, modelos de interacción, incrementos y disminuciones en las series climáticas observadas, entre otras; (7) análisis de situaciones de inestabilidad generadas debido a cambios repentinos de algunas variables exógenas del modelo; (8) análisis de sensibilidad para la posterior determinación de los parámetros que provocan variaciones apreciables en los resultados; y (9) detección de las limitaciones del modelo con miras a concentrar futuros esfuerzos.
Metodología
Módulo principal
Partiendo de la concepción sistémica de la malaria, se conciben tres componentes fundamentales: población de mosquitos (vectores biológicos), población de humanos (hospederos vertebrados) y patrones climáticos. Estos últimos controlan únicamente la interacción entre las poblaciones mencionadas (alimentación sanguínea), la ecología del vector y la disponibilidad de nichos larvarios para el desarrollo de la población de mosquitos en sus estados preimaginales.
En la figura 1 se presenta la concepción sistémica de la fase asexual endógena (esquizogonia) en el hospedero vertebrado. Se ignora, en una primera aproximación, la esquizogonia exo-eritrocítica secundaria, hecho que limita la utilización del modelo para la representación de la transmisión de la malaria por Plasmodium falciparum. Un hospedero
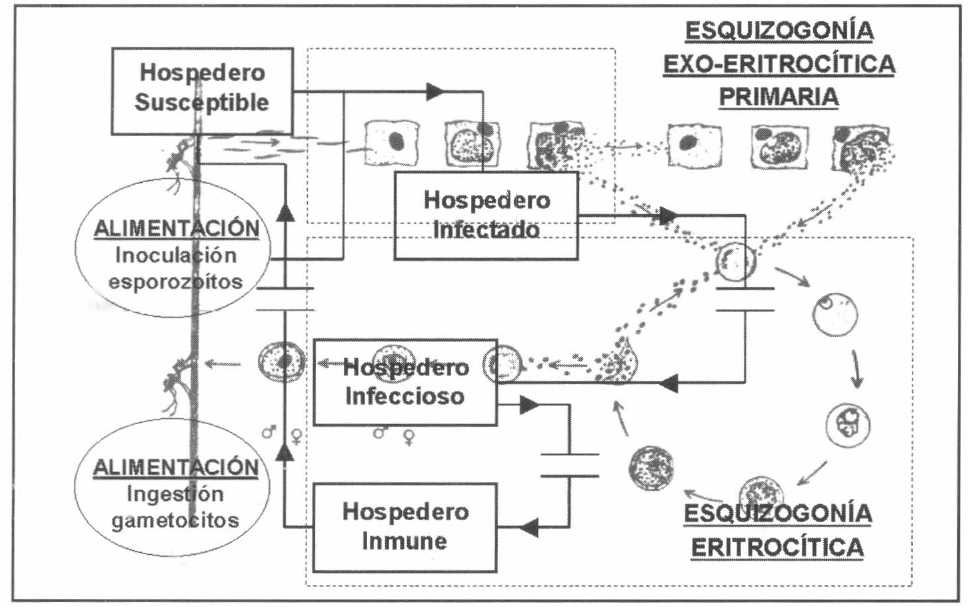
En la figura 2 se presenta la concepción sistémica de la fase sexual exógena (esporogonia) en el mosquito hospedero. Un vector adulto hembra pasa del estado
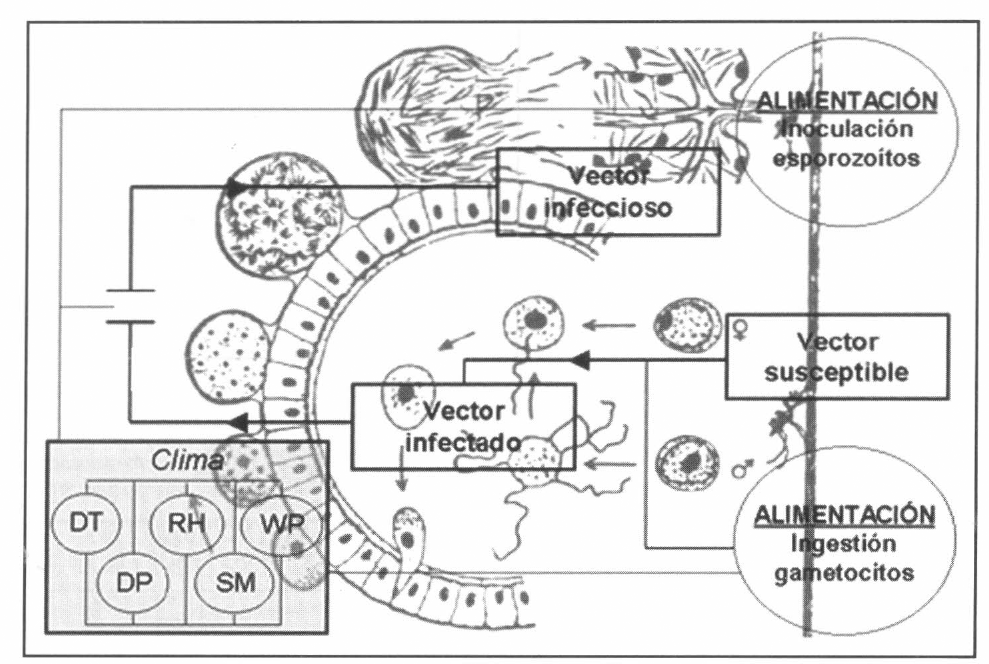
En la figura 3 se presenta el esquema general (macro-diagrama de interacciones) del modelo desarrollado. Se utiliza un modelo matemático continuo dinámico soportado en el sistema de ecuaciones diferenciales ordinarias no lineales, presentadas a continuación y que se resuelven por el método numérico Runge-Kutta IV. Cabe notar, que las ecuaciones para la representación de la ecología del vector están aún en proceso de elaboración y gran parte de los parámetros están siendo estimados gracias a la información de campo y bioensayos.
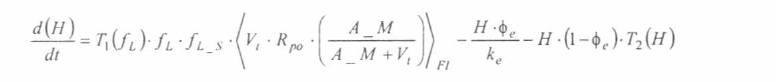
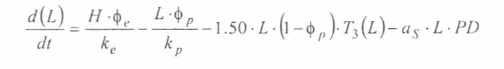
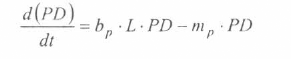
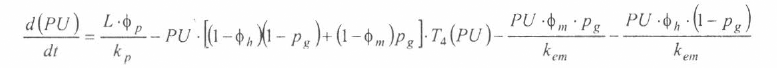
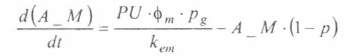

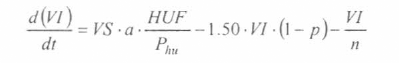

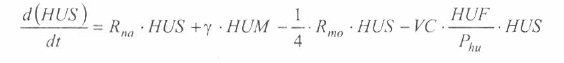
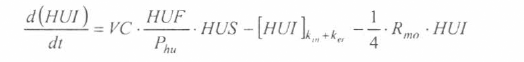

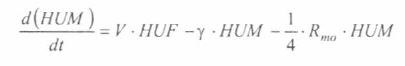
La variable de estado 'humanos susceptibles' disminuirá por mortalidad natural de individuos (tercer término) y por aquellos individuos que son infectados durante la alimentación sanguínea de vectores adulto hembra en estado infeccioso (inoculación de esporozoitos).
Concepción sistémica de la esquizogonia. La flecha continua entre los dos primeros estados representa la transferencia de la totalidad de información. Las flechas discontinuas, los retardos en las transferencias de material.
Descripción de variables de la componente población de mosquitos.
Módulo de disponibilidad de criaderos
Se construyeron, vía lógica difusa, unas funciones tabuladas exógenas al módulo principal que representaban las fluctuaciones de nivel en criaderos de diferente capacidad (viabilidad de los hábitat de reproducción en cuanto a la disponibilidad del recurso hídrico) y afectaban la tasa de ovipostura en la componente población de mosquitos. Se asumieron unos criaderos tipo embalse sometidos a precipitación observada, evaporación calculada por diferentes metodologías (Fig. 4) e infiltración. Se requirió de la estimación del número de días consecutivos que un criadero de una lámina dada, partiendo de una determinada capacidad inicial, y sometido a las tres variables mencionadas anteriormente, permaneció en un nivel de interés. Como de forma general los Anopheles prefieren las aguas frescas libres de materia orgánica (caso analizado Anopheles albimanus), se determinó el número de días consecutivos (que requiere una cohorte de huevos para adelantar todos sus estados preimaginales y alcanzar la emergencia) que un determinado criadero permaneció lleno (90-100% de capacidad) cuando se sometió a flujo neto de agua (diferencia entre precipitación observada, evaporación real estimado e infiltración constante). El óptimo de un determinado criadero se alcanza si permanece entre el 90 y 100% de su capacidad por lo menos 12-15 días consecutivos (Fig. 5). Finalmente, como los huevos de la gran mayoría de Anopheles no pueden sobrevivir luego de la desecación, se asumió que el criadero no garantizaba el desarrollo de la especie en sus estados preimaginales si excedía 2 días consecutivos SECO.
Módulo de interacciones predador-presa-recursos
Se representaron las interacciones entre los estados preimaginales de la población de mosquitos (principalmente larvas) y sus predadores naturales. Se consideró en una primera aproximación el modelo tipo Lotka-Volterra (Lotka 1925; Volterra 1926), integrándolo al sistema de ecuaciones diferenciales principal (Fig. 6). Los parámetros de los demás modelos analizados [(predador-presa tipo Henize (1971), presa-recursos tipo Kaibab III (Aracil y Toro 1993), modelo ecológico con forzamiento externo (Fig. 7) (Aracil y Toro 1993) y sistema ecológico autónomo (Fig. 8) (Aracil y Toro 1993)] están siendo calibrados vía bioensayos en los laboratorios del proyecto.
Simulación
Se representó la incidencia de malaria por P. falciparum en el municipio de Nuquí, departamento del Chocó, costa pacífica colombiana (Fig. 9), para el período 01/nov/1997-28/feb/1999. Las mediciones entomológicas en campo (tasas de picadura intra y peridomiciliaria, tasas de paridad y número de larvas capturadas en criaderos (Fig. 10)) se adelantaron en conjunto con la Universidad de Antioquia y la Corporación para Investigaciones Biológicas; la información hidrometeorológica (valores medios diarios de temperatura [°C], valores medios diarios de humedad relativa [%] y registros totales diarios de precipitación (Fig. 11) se recopiló por la estación climatológica 5601503 Amargal (Arusí, 05°36′N, 77° 30'W, 30 msnm), operada por el IDEAM para el período 01/nov/1997-28/feb/2001 (40 meses de registros, 1.200 días de horizonte); las mediciones epidemiológicas (incidencia mensual de malaria por P. vivax y P. falciparum para el período nov/1997-feb/1999) se obtuvieron del Servicio Seccional de Salud Departamental (Fig. 12).
Concepción sistémica de la esporogonia.
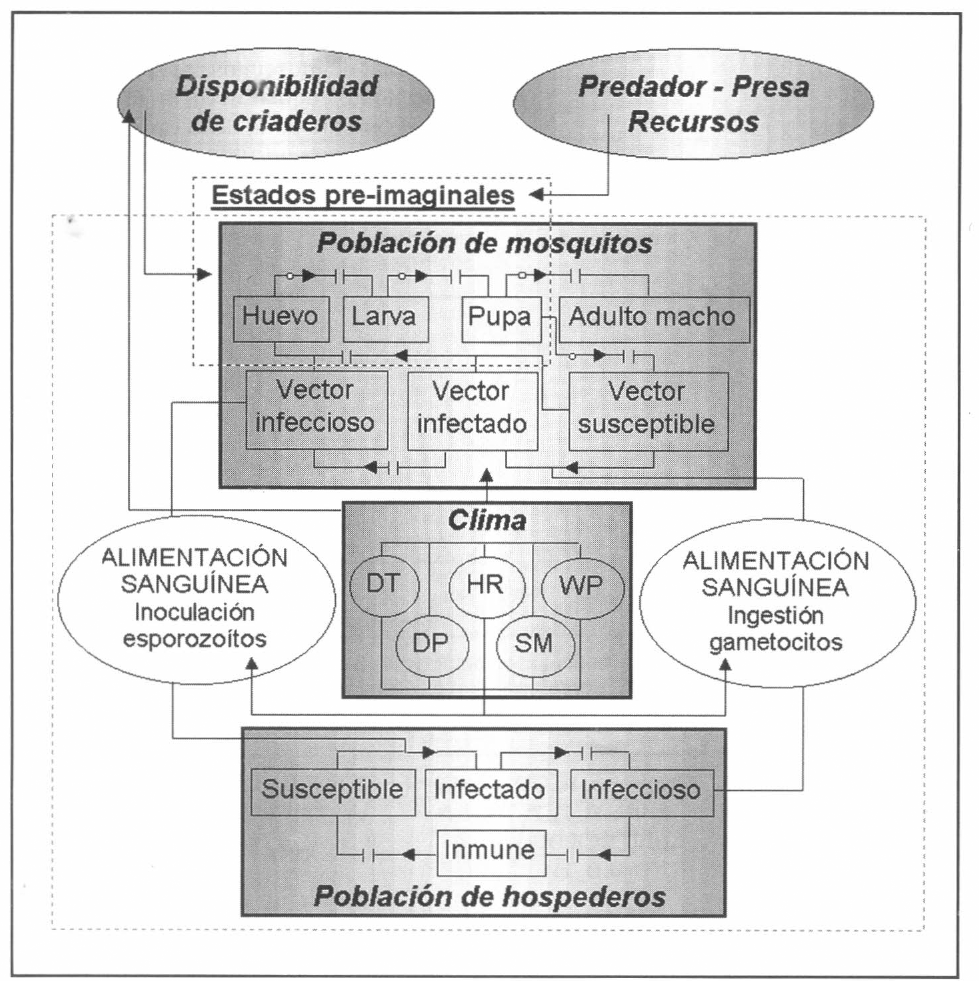
Macro-diagrama de interacción. Los óvalos laterales denotan los mecanismos de transmisión (interacción entre poblaciones vía alimentación sanguínea). Los óvalos superiores, los módulos paralelos al modelo principal que afectan los estados preimaginales de la población de mosquitos. En las flechas discontinuas, los pequeños círculos denotan las regulaciones en las transferencias de material: eficiencias de eclosión, pupación y emergencia. Las variables DT, HR, WP, DP y SM denotan, respectivamente, los valores medios diarios de temperatura, valores medios diarios de humedad relativa, patrón de vientos (en velocidad y dirección), valores totales diarios de precipitación y humedad del suelo. En una primera fase se consideran únicamente las variables climáticas DT, HR y DP.
Descripción de variables de la componente población de hospederos
Descripción de variables de los modelos de interacción
Resultados
El proceso de análisis de escenarios se ejecutó para los primeros 480 días del horizonte de simulación (01/nov/1997-28/feb/1999), debido a que la información epidemiológica sólo estaba disponible para este período. La población total en riesgo se asumió equivalente a 2.477 individuos que habitaban la zona rural del municipio de Nuquí a finales del año 1996 (información demográfica del Ministerio de Salud). El número de individuos en estado infeccioso ascendió a 111 pobladores, equivalente al total de casos de malaria por P. falciparum registrados en el mes de octubre de 1997. Los resultados de simulación para el escenario base asumido se presentan en la figura 13.
Se consideraron además dos escenarios alternos de simulación: incrementos en las temperaturas diarias registradas de +1 y +2°C (Fig. 14a) y disminuciones de -0.5 y -1°C (Fig. 14b) para todo el horizonte de simulación.
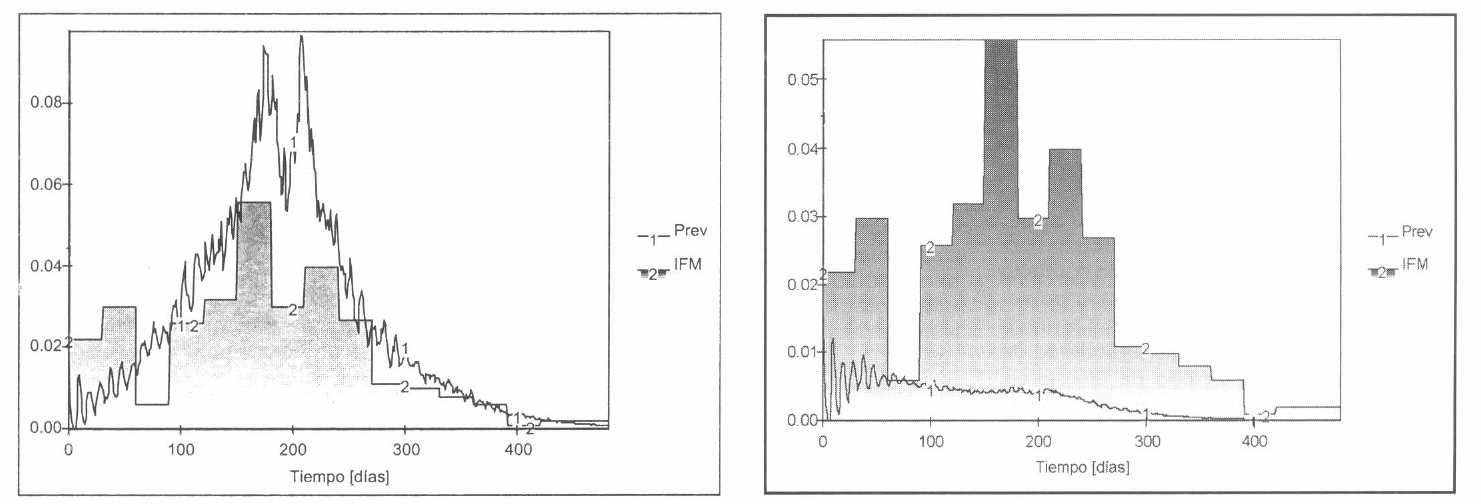
En el proceso de evaluación-validaciónanálisis, se consideraron las posibles situaciones de inestabilidad generadas debido a cambios repentinos de algunas variables exógenas (Fig. 15) y se detectaron los parámetros que provocaban variaciones apreciables de los resultados (Fig. 16).
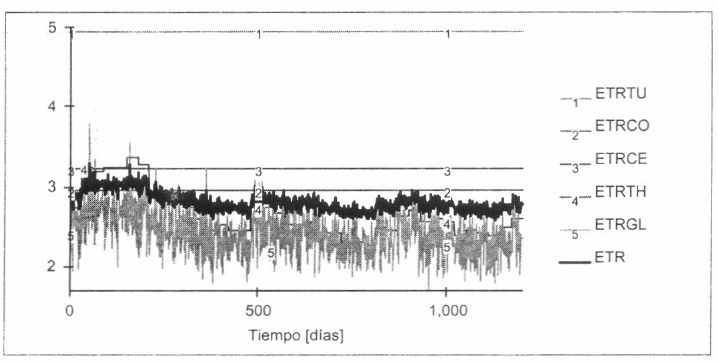
Láminas de evaporación real ETR [mm/día] estimadas. (1) ETRTU: ecuación de Turc (1945); (2) ETRCO: Coutagne (1974); (3) ETRCE: Cenicafé (1997); (4) ETRTH: Thornthwaite (1948); (5) ETRGL: García-López; y ETR: promedio de (2), (3), (4) y (5). Horizonte: 1.200 días (nov/1997-feb/2001); Time Step: 1 día; Método de Integración: Euler (fixed step).
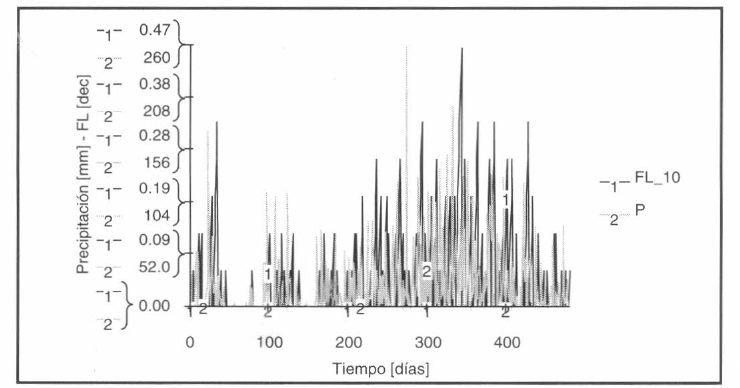
Modelo de disponibilidad de criaderos. Nicho larvario de 10 mm de lámina asumido inicialmente al 30% de capacidad. Si la función FL_10 iguala 0.0, el criadero NO es viable; si la función FL_10 iguala 1.0, el criadero es viable (permanece entre 90 y 100% de capacidad más de 12 días consecutivos).
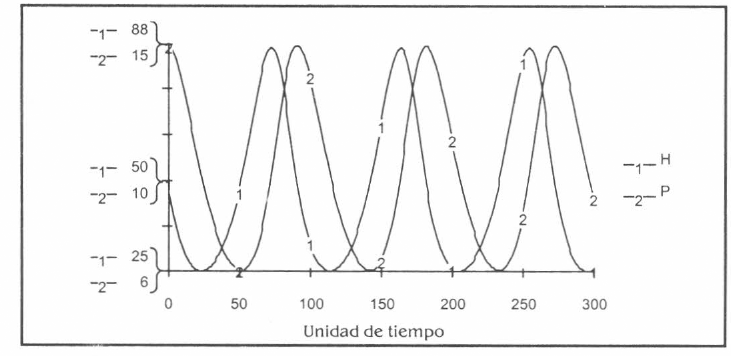
Evolución en el tiempo de las poblaciones presa H y predador P según modelo tipo Lotka-Volterra. Horizonte de tiempo: 300 unidades, Dt: 1 unidad. Algoritmo de solución: Runge-Kutta IV (fixed step).
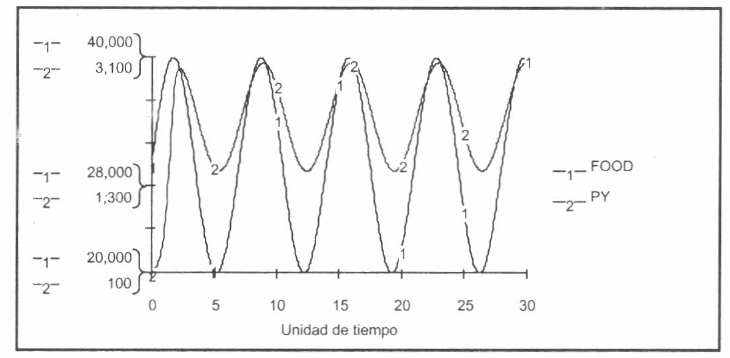
Evoluciones de la población presa PY y los recursos de comida del hábitat FOOD según modelo ecológico con forzamiento externo. Horizonte: 30 unidades de tiempo, Dt=0.05. Algoritmo: Runge-Kutta IV (fixed step).
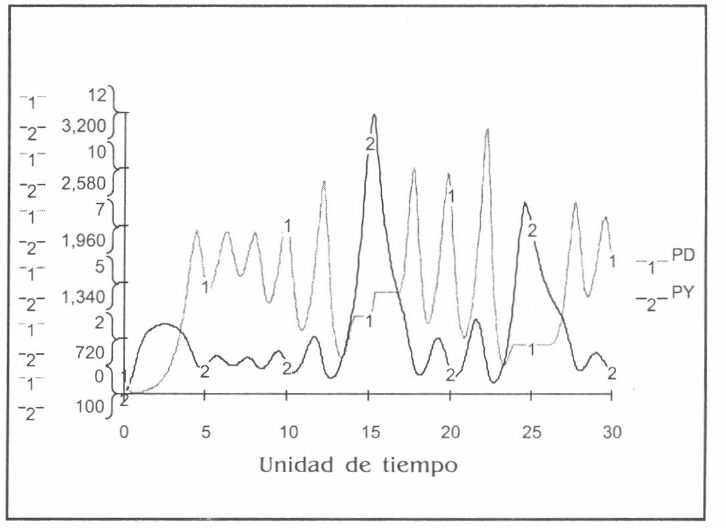
Evoluciones de las poblaciones predador PD y presa PY según sistema ecológico autónomo. Horizonte: 30 unidades de tiempo, Dt=0.05. Algoritmo: Runge-Kutta IV (fixed step).
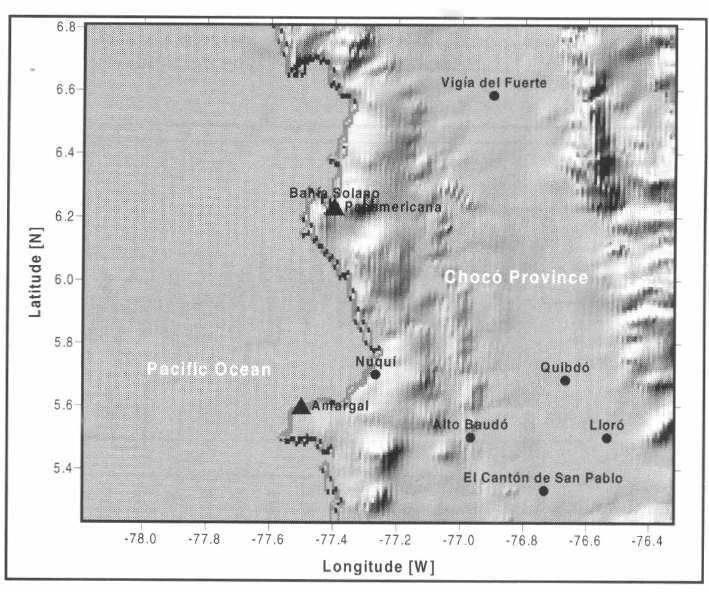
Zona de estudio. Mapa de elevación digital con una resolución espacial de 30" de arco. Principales localidades y estaciones climatológicas disponibles en la zona.
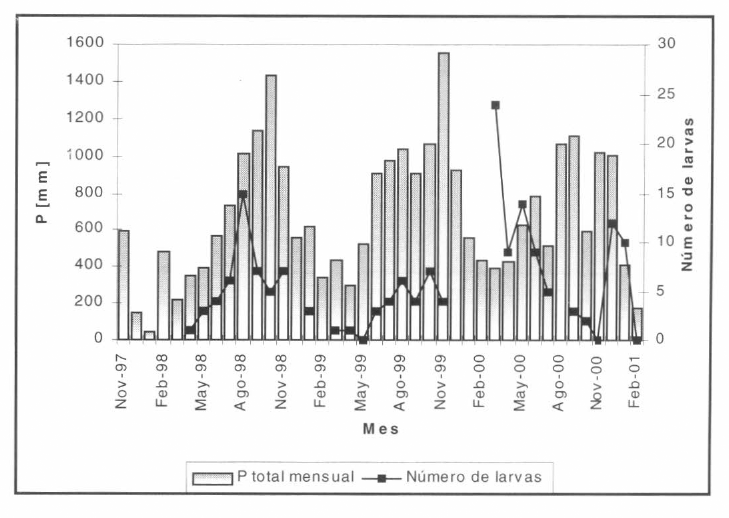
Número total de larvas capturadas en los criaderos. Campaña de caracterización de nichos larvarios abril/1998-marzo/2001.
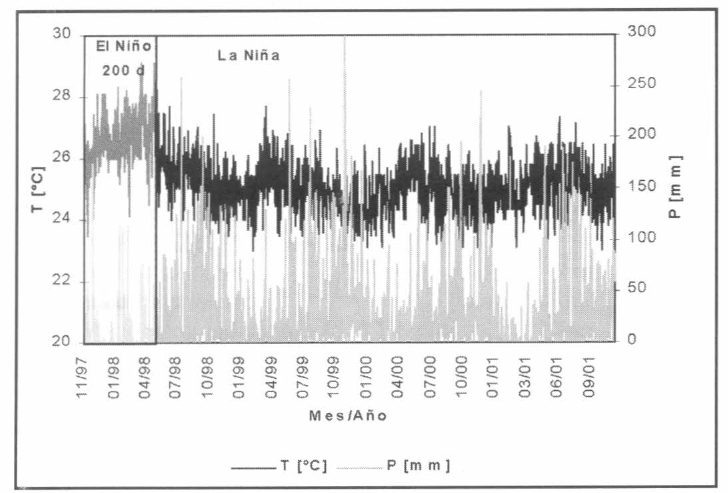
Información hidrometeorológica. Series de valores medios diarios de temperatura [°C] y valores totales diarios de precipitación [mm]. Período de registros seleccionado: 01/nov/1997-28/feb/2001 (40 meses, 1.200 días de horizonte de simulación). La
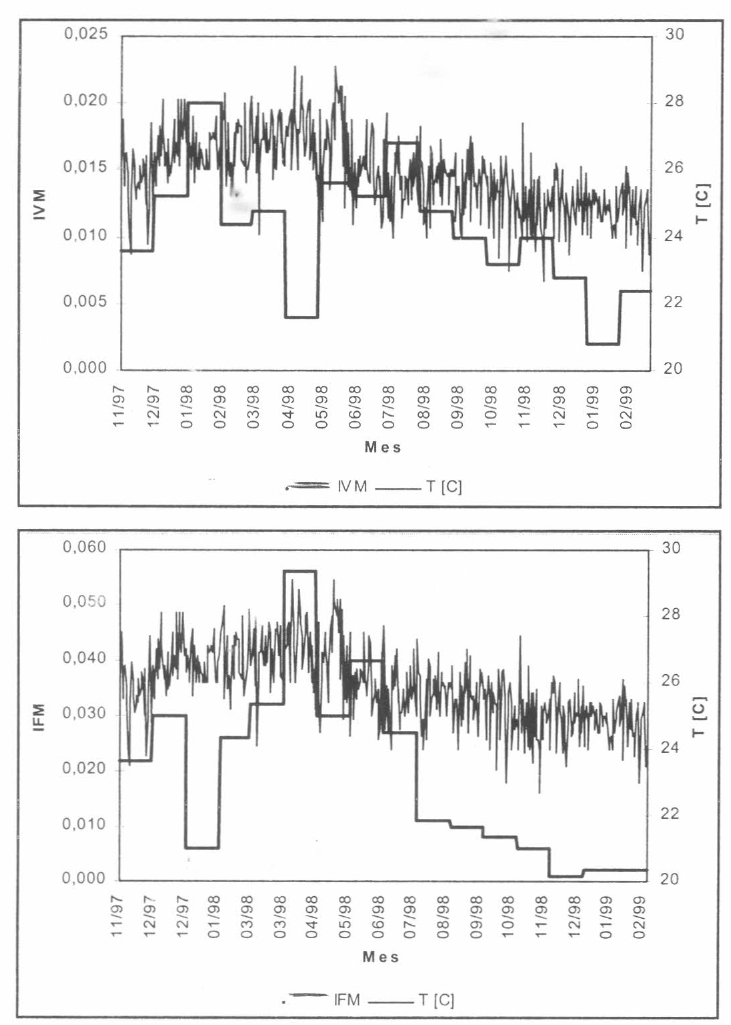
Incidencia mensual de malaria por P. vivax IVM y P. falciparum IFM. Número total de casos registrados / totalidad de la población en riesgo. La variable T denota los valores medios diarios de temperatura registrados por la estación climatológica (obsérvense las tendencias creciente (nov/1997-abr/1998) y decreciente (abr/ 1998-feb/1999) de la serie).
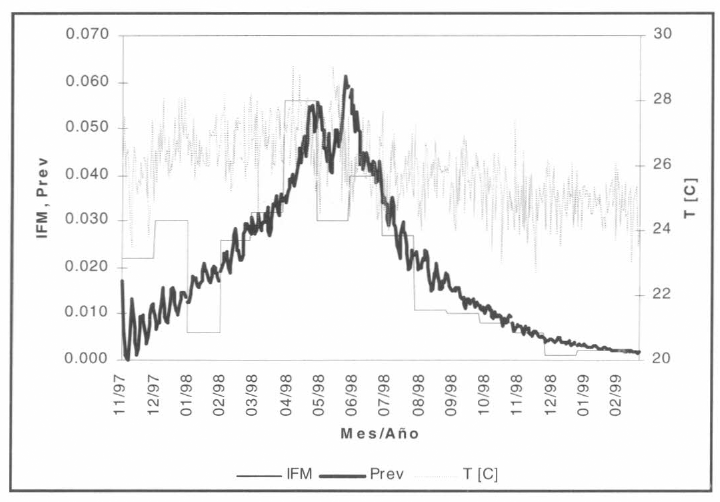
Escenario base de simulación de malaria por P. falciparum.
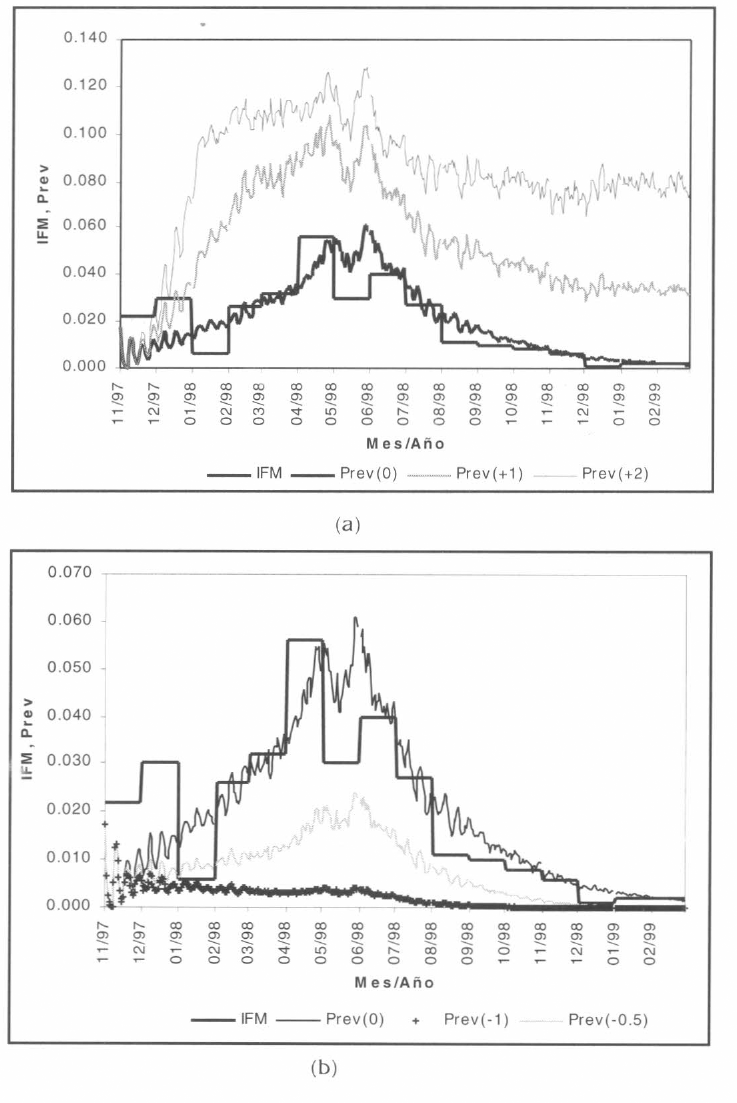
Resultados de simulación, escenarios alternos.
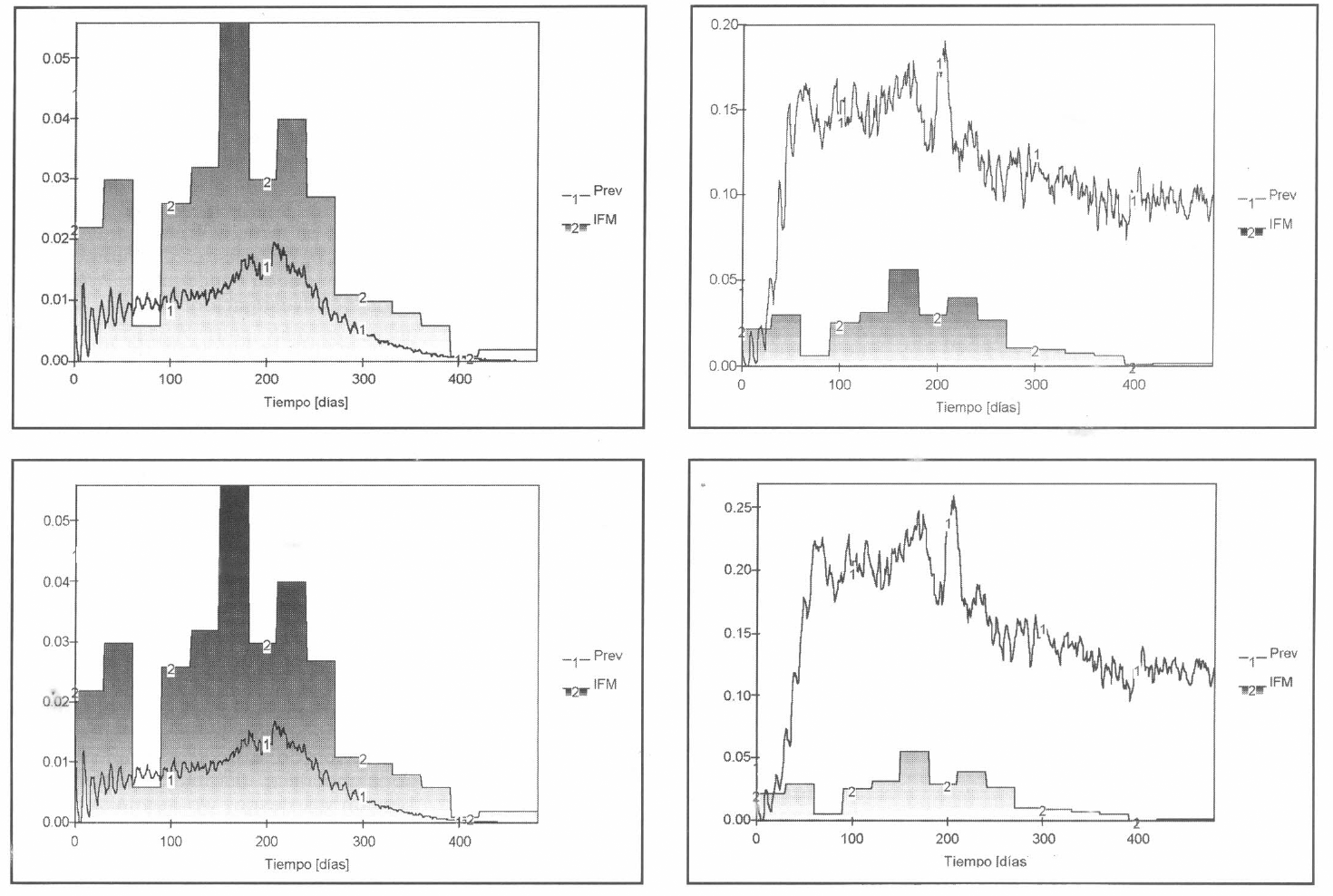
Posibles situaciones de inestabilidad.
Análisis de sensibilidad.
Discusión
El modelo matemático desarrollado reproduce la incidencia mensual de malaria por P. falciparum registrada en el municipio de Nuquí, Chocó, para el período disponible 01/noviembre/1997-28/febrero/1999. Mediante el sistema de ecuaciones diferenciales concebido se logra representar el brote epidémico de malaria que ocurrió en esta zona endémica del país como respuesta a la ocurrencia de la fase cálida del evento ENSO en los 200 primeros días de simulación. El modelo tiene la capacidad de reflejar las tendencias crecientes y decrecientes observadas en la incidencia de la enfermedad en este período de tiempo, como consecuencia de los incrementos y disminuciones, respectivamente, de los valores promedio diarios de temperatura observados. Se logran representar, además, las fluctuaciones en la densidad vectorial (respuesta a los valores totales diarios de precipitación) de acuerdo con las tasas de picadura a seres humanos registradas en las campañas de colecta de adultos.
El ejercicio de simulación llevado a cabo en el horizonte de tiempo seleccionado indica que la temperatura es la variable climatológica que controla en mayor grado las variables entomológicas más relevantes de transmisión e incidencia: ciclo de ovoposición, ciclo esporogónico y longevidad, que finalmente permiten la determinación de las tasas diarias de contacto potencialmente infeccioso entre las poblaciones de hospederos y vectores. La precipitación, que controla la disponibilidad de criaderos para el desarrollo de la especie en sus estados preimaginales, finalmente permitirá el entendimiento de las fluctuaciones estacionales de la densidad vectorial. En el momento en el cual se desarrolle un modelo de interacción que articule la densidad y la incidencia, se podrá analizar el peso de esta variable climatológica en la transmisión y posterior prevalencia.
Para un incremento de +2°C en las temperaturas promedio diarias observadas, el modelo matemático muestra que el punto máximo de prevalencia de la enfermedad se incrementa del 6 al 12% de la población en riesgo (duplicación de casos por P. falciparum). Además, se observa que la malaria deja de evidenciar el brote epidémico registrado para convertirse en una endemia que somete al 8% de la población (agosto/1998 en adelante). Esto responde directamente a que el incremento en la temperatura genera una evidente disminución en los ciclos esporogónico y de ovoposición de la población vector.
Para el caso de las disminuciones en los registros climáticos observados, la modelación sugiere que si las temperaturas promedio diarias del municipio de Nuquí no hubieran experimentado el incremento de +0.5-1°C como respuesta a la ocurrencia de la fase cálida del evento ENSO, es posible que el punto máximo de prevalencia de la enfermedad por P. falciparum en el período considerado tan sólo hubiera ascendido al 2% de la población en riesgo. Si las temperaturas mostraran el comportamiento hipotético considerado, la enfermedad podría llegar a ser controlada por la misma respuesta del ecosistema (en términos de que se controla el desarrollo de la especie vector y el parásito incubado en la misma).
Aportes
El modelo matemático y el proceso de simulación constituyen, aún a este nivel preliminar, una herramienta fundamental para el entendimiento de las dinámicas de transmisión y las asociaciones clima-malaria. Se destacan las siguientes fortalezas: (1) articulación de gran parte del conocimiento desarrollado en torno a la enfermedad por las disciplinas de entomología, epidemiología, medicina, limnología, hidrología y climatología; (2) representación de la ecología del vector como un primer esfuerzo para simular las fluctuaciones estacionales de la densidad de mosquitos adultos hembra; (3) identificación en el ciclo de transmisión de las variables entomológicas que son altamente influenciadas por los patrones climáticos; (4) inclusión del módulo paralelo de disponibilidad de criaderos con miras a representar el desarrollo de la población de mosquitos en sus estados preimaginales; (5) inclusión del módulo paralelo de interacciones predador-presa en los estados preimaginales de la población de mosquitos; (6) concepción como una herramienta para el
Conclusiones
Conclusiones generales
Teniendo presente el sistema de ecuaciones diferenciales desarrollado y el horizonte de tiempo seleccionado, el resultado de la simulación sugiere que las variables entomológicas más relevantes para la determinación del grado de interacción entre las poblaciones de mosquitos y hospederos vertebrados, y la posterior estimación de la prevalencia de la enfermedad son: el
Si se acepta la hipótesis de que la sola disminución en el período de incubación del parásito dentro del vector como respuesta al incremento en la temperatura de una determinada zona endémica genera los brotes epidémicos de la enfermedad, se está ante una amenaza evidente. En este sentido, se debe generar una estrategia defensiva que considere una o más de las siguientes tres componentes: (a) diseñar sistemas que alerten sobre la emergencia o resurgencia de este tipo de enfermedades infecciosas o de los vectores que la portan. Esto podría generar medidas de control de la proliferación de vectores sin perjuicio para el medio ambiente, diseñar campañas de educación ambiental para la protección de la población humana de la zona endémica, distribuir vacunas a la población en riesgo, y/o suministrar tratamientos a los casos infecciosos; (b) concentrar esfuerzos en la predicción de condiciones climatológicas y ambientales que se pueden tornar favorables para los brotes epidémicos de la enfermedad, de tal manera que los riesgos sean minimizados; y (c) enfrentar el calentamiento global directamente. Esto es, limitar actividades humanas que contribuyen al calentamiento o que agravan sus efectos.
Implicaciones
Aunque la malaria tiene origen en numerosos factores socio-económicos, demográficos y hereditarios, los factores ambientales y la variabilidad climática también contribuyen a explicar las fluctuaciones en las incidencias de la enfermedad, de tal forma que es posible aprovechar esta porción de la varianza para apoyar medidas de control y mitigación. El evento ENSO se puede considerar como un 'Sistema de Alerta Temprana' (SAT) (Poveda et al. 1999) para prevenir y mitigar la enfermedad en el país. Así, una vez se tengan predicciones con alta confiabilidad y/o se esté presentando un evento del ENSO, se puede comenzar a utilizar insecticidas, a impregnar mosquiteros con piretroides, a tomar medidas de control biológico, y otras medidas no permanentes como planificar inventarios de drogas, implementar campañas de diagnóstico y vacunación, acelerar el entrenamiento de personal, diseminar información a través de los medios de comunicación, entre otras (Poveda et al. 1999).
En general, con este trabajo de investigación, se ha construido una herramienta de análisis tendiente a dar pautas para el establecimiento de sistemas de prevención y control de malaria en Colombia, no sólo durante las anomalías climáticas asociadas con la ocurrencia del El Niño, sino con respecto al ciclo anual normal de la enfermedad y su asociación con la variabilidad climática 'normal'.