
Abstract
The use of entomopathogenic nematodes or entomonematodes (EN), is a possible alternative for the control of the coffee berry borer Hypothenemus hampei, in berries that have fallen to the soil. The texture of the soil is a limiting factor of EN persistence, displacement and host finding. In this experiment, the displacement of two EN species: Heterorhabditis bacteriophora (Rhabditida: Heterorhabditidae) and Steinernema feltiae (Rhabditida: Steinernematidae) towards coffee berries (mature and dry) was evaluated. The variables evaluated were index of displacement (IA), and penetration into berry (IPF), variables evaluated initially in sand. Once the berry type was selected, the effect of two soil textures, Loam clay Sandy (F.Ar.A) and loam (F) was evaluated. A rectangle of acrylic was used as an experimental unit, and 100 infective Juveniles (IJ) were placed to 5cm from the berry (mature or dry) in agreement with the treatments. The time of evaluation was 168 hours after the beginning of the experiment. In the selection of the berry, the greatest value for IA was exhibited by the dry berry and had a value of 21,13 ± 3,3% and IPF of 10,91 ± 2,2% (average + standard error), results which show that the values differ statistically (LSD 0,05) from those obtained for the mature berry (IA: 5,63 ± 1,48%; IPF: 3,56 ± 1,06%) independent of EN. With regards to the evaluated textures, they did not present any effect in the displacement of EN towards the dry berry, the greatest values for IA (15,73 ± 4,87%) and IPF (7,47 ± 3,51%) being obtained for H. bacteriophora in loam texture and for S. feltiae in Loam clay Sandy texture with 7,33 ± 3,51 (IA) and 4,33 ± 1,12% (IPF) respectively. The displacement tendencies of EN, and their effect on the mortality of individuals of H. hampei in different stages of development are discussed in this paper.
Introducción
La broca del café, Hypothenemus hampei (Ferrari), se considera como la plaga más importante del cultivo, por atacar directamente al fruto, causando pérdida de peso, depreciación del grano y pérdida en la calidad de la bebida. Su primer registro en Colombia fue en agosto de 1988, en Tumaco (Nariño). Posteriormente, se evidenció un incremento rápido de su población con una adaptación alta (Bustillo et al. 1998). Hasta la actualidad afecta, en Colombia, la totalidad del área cultivada en café (860.000 Ha). Uno de los mayores riesgos de infestación por la broca del café, en nuevas cosechas, lo ocasionan los frutos infestados con broca y los frutos sanos que han caído al suelo y han sido atacados por la plaga (Bustillo et al. 1999). Éstos albergan en su interior estadios inmaduros y adultos de la plaga y constituyen el potencial más importante en el incremento de las poblaciones. Después de un período seco se presenta un aumento poblacional de la broca en estos frutos y, con la llegada de las lluvias, los adultos emergen y se dispersan por el cafetal, infestando frutos desarrollados a partir de las primeras f1oraciones de la siguiente cosecha (Baker et al. 1994).
En la actualidad, para el control de la plaga en el suelo no se cuenta con una herramienta efectiva que esté enmarcada dentro del conocido manejo integrado de la broca (MIB), utilizándose, sólo para su control, la aplicación de insecticidas y hongos entomopatógenos en el plato del árbol y la recolección de frutos del suelo (Bustillo et al. 1998).
Para el control de la broca en frutos que han caído al suelo, una estrategia posible es aplicar un agente biocontrolador a la superficie del suelo. En teoría, los entorno nemátodos (EN) probablemente tienen la habilidad de penetrar en el fruto y causar una mortalidad significativa de los estadios de la plaga (Baker 1999). Es así como los EN al considerarse enemigos naturales de la broca (Commonwealth Institute of Biological Control 1990; Georgis y Hom 1992), se constituyen en un agente potencial para su control, al causar patogenicidad y muerte a sus estadíos de acuerdo con diversos estudios realizados en laboratorio (Allard y Moore 1989; Georgis y Hom 1992; Castillo 1995; Baker 1999). Sin embargo, para su incorporación en el control de la plaga en el suelo como una herramienta efectiva dentro del MIB, es necesario establecer si tienen la capacidad de desplazarse en el suelo, penetrar en el fruto y causar patogenicidad y muerte a los estadios de la plaga en el interior de éste. Para tal efecto, se realizó el primer estudio básico con el objetivo de determinar el desplazamiento y parasitismo de dos especies de EN: Heterorhabditis bacteríophora (Rhaditida: Heterorhabditidae) y Steínernema feltiae (Rhabditida: Steinernematidae), hacia frutos maduros y secos de café, infestados artificial mente con broca.
Materiales y Métodos
Esta investigación se realizó en el Centro Nacional de Investigaciones de Café, Cenicafé, localizado en el municipio de Chinchiná, Caldas, y ubicado a 1475 msnm.
Las especies de EN evaluadas en el experimento (H. bacteríophora y S. feltiae) fueron suministradas por los Drs. Bernard Briscoe y William Hominick del CABI * . Dichas especies se muitiplicaron y reactivaron en larvas de Gallería mellonella (Fab.) (Lepidoptera: Pyralidae) de acuerdo con el método descrito por Poinar (1979) y se mantuvieron en laboratorio a una temperatura de 12 ± 2°C.
Centre For Agriculture and Biosciences International. CABI-Biosciences, Lane, Egham.-Surrey. Inglaterra.
Los frutos de café utilizados fueron de la variedad Colombia con edad fisiológica entre 190 y 260 días de desarrollo; se infestaron utilizando una relación de tres (3) brocas por fruto durante 30 días. Al final el tiempo de infestación, se obtuvieron frutos de 220 y 290 días, que correspondieron a dos tipos de frutos: maduros y secos. Finalmente, se utilizaron frutos con un máximo de dos perforaciones por broca y se desinfectaron con NaOCI al 0,05%.
En la investigación se hicieron tres experimentos:
Desplazamiento de entomonemátodos hacia frutos infestados por broca cubiertos con arena
Se evaluó el desplazamiento de las dos especies de EN hacia frutos infestados con broca cubiertos con arena y se seleccionó el tipo de fruto que más influyó en dicho desplazamiento. Como sistema de bioensayo se utilizó una cámara de acrílico (12 x 2 x 2 cm), con secciones removibles, colocadas a cada centímetro a lo largo de la cámara (Fig. 1). Inicialmente se retiraron todas las secciones, se adicionaron 68 g de arena y se colocó un fruto (maduroo seco) en un extremo. A 5 cm del fruto (en todo el centro de la cámara), se dispensaron 100 JI. La arena utilizada previamente se lavó y se secó en estufa, a una temperatura de 110°C, se esterilizó en autoclave por 15 minutos a 15 libras de presión y posteriormente se pasó por tamices (ASTM E-1l-87 Standar U.S.A) número 60 y 325 obteniéndose un tamaño de partículas entre 425 a 585 mm, respectivamente. Finalmente esta arena se llevó a una humedad volumétrica (HV) del 15 ± 1,49% (P/P), ajustada mediante un determinador halógeno de humedad (Mettler Toledo HR 73 con precisión de 0,01%).
Desplazamiento de entomonemátodos hacia frutos infestados por broca dispuestos sobre arena
Se evaluó el desplazamiento de especies de EN hacia frutos infestados con broca dispuestos sobre arena; tanto el sistema de bioensayo como la metodología fueron las mismas relacionadas anteriormente, pero los frutos se colocaron sobre una capa fina de 30,5 g de arena. Al comprobarse desplazamiento, penetración y parasitismo en las primeras dos etapas del experimento, se seleccionó el mejor tipo de fruto para realizar el último experimento.
Desplazamiento de entomonemátodos hacia frutos infestados cubiertos y dispuestos sobre dos texturas de suelo
Se evaluó el desplazamiento de las especies de EN en dos texturas de suelo y se determinó el efecto de éstas. Las dos texturas de suelo fueron: Franco Arcilla Arenosa (FArA)y Franca (F), las cuales procedieron de dos unidades características de suelos de la zona cafetera (Chinchiná y Montenegro, respectivamente). Cada textura de suelo se esterilizó en autoclave por seis horas a 15 libras de presión y posteriormente se acondicionó a la HV registrada para condiciones de campo para las mismas texturas (FArA con 35 ± 0,43% y F con 38 ± 1,15% (P/P)). Para la evaluación de frutos cubiertos y sobre textura de suelo, en cada cámara, se adicionaron 39,5 g y 16 g de suelo, respectivamente.
Una vez montado el sistema de bioensayo para todos los experimentos, se sellaron las cámaras y se llevaron a incubación en oscuridad constante bajo condiciones controladas de temperatura (24 ± 2°C) y humedad rel tiva (80%), hasta un tiempo final para la evaluación, de 168 horas; como testigo para cada especie se utilizó el mismo sistema de bioensayo, empleando ADE sin JI. Al término de este tiempo, se colocaron todas las secciones divisorias de la cámara para contar el número de EN, en cada una de ellas, sólo para la primera etapa del experimento. Finalmente en todas las etapas se abrieron los frutos y se contó el número de JI que penetraron en éstos y el número de estadios de la plaga muertos.
Diseño experimental y variables evaluadas
La unidad experimental (U.E.) la constituyó la cámara de acrílico. Para el primer y segundo experimentos, se utilizó un diseño completamente aleatorio dispuesto en arreglo factorial 2x2+2, cuyos factores fueron: dos tipos de fruto, dos especies de ENy dos testigos, uno por cada tipo de fruto. En total se conformaron 6 tratamientos, cada uno con quince repeticiones (Tabla 1). En el tercer experimento en frutos cubiertos y sobre las texturas de suelo, se empleó el mismo arreglo factorial 2x2+4, cuyos factores fueron: dos texturas de suelo, dos especies de EN y cuatro testigos: (ADE sin JI, por cada textura y EN), para un total de ocho tratamientos, cada uno con quince repeticiones (Tabla 2).
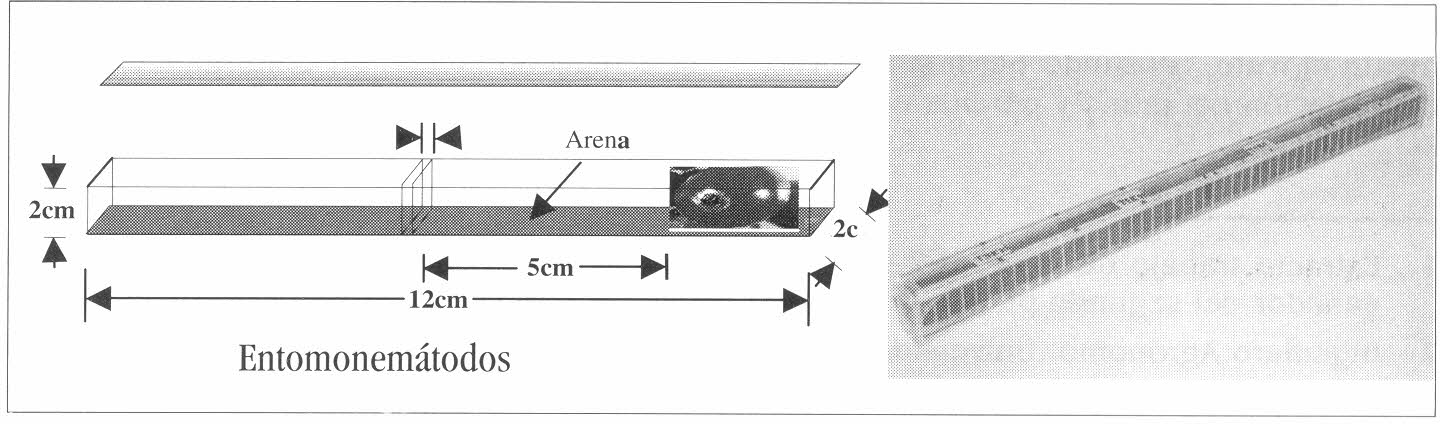
Modelo de las cámaras de acrílico utilizadas en el bioensayo.
Descripción de los factores y tratamientos involucrados en el experimento de desplazamiento de entomonemátodos hacia frutos brocados cubiertos por arena
Tratamientos evaluados en el desplazamiento de entomonemátodos hacia frutos brocados cubiertos y sobre dos texturas de suelo
Las variables que se evaluaron en los tres experimentos, en las cuales se determinó el desplazamiento de las especies de EN (IE, IA), la penetración (IPF) y la mortalidad de individuos en diferentes estadios de la broca (PMIB) en el fruto, se describen a continuación:





Luego de obtener los porcentajes de mortalidad para cada tratamiento y testigos, se procedió a estimar el

En todos los experimentos se calcularon para las variables respuesta IA, IPF, PMIB, IE, IR promedios, variación, análisis de varianza (ANAVA) al nivel del 5%. Se aplicó una prueba de contraste al nivel del 5%, en el caso que la interacción simple entre los factores de los tratamientos evaluados por variable, fuera significativa. Se aplicó una prueba de comparación de diferencia mínima significativa (LSD) y Tukey al nivel del 5. Finalmente para el análisis de los resultados, los datos que presentaron una alta variabilidad, se transformaron con la fórmula:
Resultados y Discusión
Desplazamiento de entomonemátodos hacia frutos infestados en una cámara cubiertos con arena
En general, las dos especies de EN (H. bacteriophora y S. feltiae) se desplazaron 5 cm a lo largo de la cámara dispuesta en sentido horizontal, desde el punto de inoculación inicial de los JI hasta localizar el fruto, penetrándolo por el orificio de perforación hecho por la broca. Los resultados de dicho desplazamiento y la mortalidad de los estadios de la plaga a causa de las especies de EN evaluadas, se observan en las Tablas 3-7 donde se presentan el promedio y coeficiente de variación de las variables evaluadas IE, IA, IPF, PMIB, e IR, respectivamente.
Promedios y variación para la variable IE en los tratamientos evaluados en el desplazamiento de JI hacia frutos cubiertos con arena
Promedios y variación para la variable IA en los tratamientos evaluados en el desplazamiento de JI hacia frutos cubiertos con arena
Los datos fueron transformados por la fórmula
Promedios y variación para la variable IPF en los tratamientos evaluados en el desplazamiento de JI hacia frutos cubiertos con arena
Los datos fueron transformados por la fórmula
Promedios y variación para la variable PMIB en los tratamientos evaluados en el desplazamiento de JI hacia frutos cubiertos con arena
Los datos fueron transformados por la fórmula
Promedios y variación para la variable IR en los tratamientos evaluados en el desplazamiento de JI hacia frutos cubiertos con arena
El desplazamiento de los JI en arena es gradual, puesto que a las 168 horas, momento de la evaluación para todos los tratamientos, los IA e IPF guardan una relación directa. Los primeros JI que lograron alcanzar el fruto penetraron, quedando los demás JI en la proximidad del fruto o en las diferentes secciones de la cámara en dirección a éste (Tablas 3 y 4).
En este orden de ideas, esta misma tendencia de desplazamiento se refleja en el IE para las dos especies de EN evaluadas, con promedios de 12,06% у 36,18% para S. feltiae y H. bacteriophora, respectivamente. Todos los JI no se desplazaron al mismo tiempo y una parte de éstos permaneció en la primera sección de la cámara (punto de inoculación de los JI).
Por otra parte, las dos especies de EN presentaron respuesta para las variables IA e IPF en los dos tipos de frutos de café (maduros y secos), donde S. feltiae fue la especie que presentó los mayores valores para dichas variables (Tablas 4 y 5); estos valores explican la menor respuesta para la variable IE para dicha especie. Sin embargo, para H. bacteriophora, a pesar de que presentó un IE mayor y una menor respuesta para las variables IA e IPF mostró el PMIB más alto de este experimento (Tabla 6).
Estos resultados demuestran que una parte de los JI de H. bacteriophora se desplazaron, penetraron el fruto e infectaron individuos de broca en diferentes estadios en un tiempo menor al transcurrido para la evaluación final del experimento (168 horas); en las distintas secciones de la cámara (sin tener en cuenta la sección donde se inocularon los JI), sólo se recuperó el 3% de sus JI, a diferencia de la especie S. feltiae donde se recuperó el 14% de sus JI en las mismas secciones.
Schroeder y Beavers (1987), en un bioensayo al evaluar el desplazamiento de algunos EN utilizando una columna en sentido horizontal, encontraron que Heterorhabdítidos como H. bacteriophora y H. heliothidis presentaron un mayor desplazamiento con relación a algunos Steinernemátidos como S. glaserie. Así mismo, trabajos realizados por Mannion y Jansson (1992) señalan que Jl de la especie H. bacteriophora tienen una mayor velocidad de desplazamiento en sentido horizontal y causan mayor infectividad, en relación con los Steinernemátidos S. feltiae y S. glaserie.
Cabe resaltar que a pesar de que las mortalidades de individuos de broca fueron menores en fruto maduro así mismo es alta, con promedios en el orden del 42,49 у 32,33% para la especies S. feltiae у Н. bacteriophora, respectivamente (Tabla 6). Estos resultados son satisfactorios si se tiene en cuenta que los JI en menos de 168 horas tuvieron que desplazarse y localizar los estadios de la plaga para causar las mortalidades registradas. Teniendo en cuenta el IPF obtenido para ambas especies, 3,56 ± 1,06%, se puede suponer que a mayor tiempo de evaluación, el PMIB aumente.
En cuanto a la respuesta para la variable IR, ésta fue muy baja para las dos especies de EN, debido a que no se encontraron Jl en las secciones en sentido opuesto hacia el fruto, antes del punto de inoculación inicial, lo que reafirma que los volátiles emitidos por el fruto y los estadios de la plaga son factores de atracción para los JI (Tabla 7).
En segunda instancia, en este experimento se seleccionó el GMF que influyó más en la respuesta de las variables evaluadas. El análisis de varianza no sugiere efecto de la interacción el tipo de fruto y las especies de EN evaluadas para las variables IA e IPF, pero en el PMIB sí presentó efecto dicha interacción, lo que determinó que el fruto es factor de atracción de los EN e incidió directamente en las variables respuesta. Por otra parte el análisis de varianza mostró efecto del GMF en el IA e IPF y la diferencia mínima significativa (LSD P≤0.05) mostró diferencias a favor del fruto seco para las mismas variables (Tablas 3 y 4) seleccionándose finalmente el fruto seco para los bioensayos con las texturas de suelo en el último experimento.
Así mismo, al evaluar el PMIB, la prueba de comparación utilizada en el análisis de covarianza, igualmente sugiere efecto a favor del fruto seco. Esta respuesta se presenta en el PMIB a causa de la acción patogénica de la especie H. bacteriophora, cuya mortalidad fue la más alta de todo el experimento (48,82%) (Tabla 6).
La respuesta presentada a favor del fruto seco en todas las variables evaluadas, puede obedecer a que éste presentó cerca de un 50% más de individuos de broca en su interior (15,89 ± 1,37) en relación con el número de estadios encontrados en fruto maduro (8,64 ± 0,76), lo que pudo atraer en mayor escala a los EN, por la concentración de volátiles emitidos por la plaga.
Desplazamiento de entomonemátodos hacia frutos infestados por broca dispuestos sobre arena
Se presentó una respuesta muy baja para las variables evaluadas, IA e IPF, con valores de 0,93 ± 0,18% y 0,07 respectivamente.
De esta forma se confirma que la posición del fruto en arena determina el desplazamiento y penetración de los JI, debido a que éstos, al no estar directamente en contacto con el orificio de perforación del fruto, se encuentran impedidos para ingresar e infectar los estadíos de la broca, a diferencia del experimento anterior, donde las partículas de arena se constituyeron como el puente de contacto entre los JI y el orificio del fruto para que éstos ingresen.
Desplazamiento de dos especies de entomonemátodos hacia frutos de café cubiertos por texturas de suelo
En general H. bacteriophora y S. feltiae se desplazaron en las texturas de suelo presentando respuesta para las variables evaluadas IA, IPF y PMIB (Tablas 8-10). Dichas variables, al igual que los experimentos en arena, mostraron una relación directa, donde parte de los JI que lograron desplazarse al fruto (IA), penetraron a éste (IPF), para el tiempo de evaluación final, tendencia que se observa en las dos texturas y especies de EN evaluadas.
Promedios y variación para la variable IA en los tratamientos evaluados en el desplazamiento de JI hacia frutos cubiertos en diferentes texturas de suelo
Los datos fueron transformados por la fórmula
Promedios y variación para la variable IPF en los tratamientos evaluados en el desplazamiento de JI hacia frutos cubiertos en diferentes texturas de suelo
Los datos fueron transformados por la fórmula
Promedios y variación para la variable PMIB en los tratamientos evaluados en el desplazamiento de JI hacia frutos cubiertos en diferentes texturas de suelo
Los datos fueron transformados por la fórmula
El análisis de varianza no mostró efecto de la interacción entre las texturas de suelo y las especies de EN avaluadas (factores) para las variables respuesta IA e IPF; sin embargo, los factores por separado sí presentaron significancia. Igualmente, no existieron diferencias significativas para estas variables ni por especie de EN ni por textura de suelo, de acuerdo con la prueba de diferencia mínima significativa (LSD P≤0.05) (Tablas 8 y 9).
Estos resultados confirman que las texturas de suelo no presentan efecto alguno en el desplazamiento de las especies de EN hacia el fruto, ya que éstas tienen características óptimas que facilitan la movilidad de los JI, como la baja densidad aparente con valores de 0,7 y 0,8 g/cc para las texturas FArA y F, respectivamente.
De esta forma la especie H. bacteriophora es favorecida principalmente por la baja densidad aparente en las texturas empleadas, presentando los valores más altos para las variables respuesta IA e IPF, debido a que los JI se desplazaron con facilidad, conservando su viabilidad (Tablas 8 y 9). De acuerdo con estudios realizados por Portillo et al. (2000), una densidad alta aparente afecta el desplazamiento y sobrevivencia de los JI, por el reducido tamaño de poro, ya que los JI de algunas especies de EN pueden tener un diámetro mayor con respecto al diámetro de poro de la textura, ante lo cual ven impedido su desplazamiento hacia el hospedante. Por el contrario las densidades bajas permiten el movimiento continuo de los JI entre las partículas.
Con base en lo anterior se plantea que, a pesar de que no hay un efecto en el desplazamiento y penetración hacia el fruto infestado para las texturas probadas y las especies de EN, cada una presenta características particulares que hacen que su desplazamiento hacia el fruto se vea favorecido en mayor o menor grado, donde H. bacteriophora y S. feltiae presentaron una mayor respuesta para las variables evaluadas en las texturas F y FarA, respectivamente (Tablas 8 y 9). En este mismo sentido, trabajos realizados por Barbercheck y Kaya (1991), concluyen que el desplazamiento y movilidad de H. bacteriophora es mayor en texturas franco arenosas y orgánicas que en suelos de texturas arcillosas; así mismo, S. carpocapsae presenta un desplazamiento mayor en un suelo arenoso que en un suelo orgánico.
Con relación a la variable PMIB, el análisis de varianza sugiere el efecto de la textura y la prueba de comparación utilizada en el análisis de covarianza, sugiere el efecto a favor de la textura F para ambos EN, donde los mayores PMIB se obtuvieron por S. feltiae (33,43%) y H. bacteriophora (31,20%) (Tabla 10). En trabajos realizados con otros insectos tales como Galleria mellonella (Fab.), Lucilia cuprina (Wied.) y Cyrtomenus bergi (Fro.), donde se determinó la infectividad de especies como H. bacteriophora, S. glaserie y S. carpocapsae, se pudo comprobar que el tipo de textura influía en la infectividad de los JI y decrecía si la proporción de arcilla aumentaba (Choo y Kaya 1991; Barbercheck 1992). Lo anterior respalda los resultados encontrados en este experimento, de tal manera que suelos con contenidos de arcilla dificultan el desplazamiento de las especies de EN hacia el fruto infestado y por ende afectan la infectividad sobre los individuos en diferentes estadios de la broca.
Es de resaltar que la especie H. bacteriophora, en las dos texturas evaluadas, fue la única que se desarrolló y multiplicó en estadios de broca dentro del fruto, con emergencia de 141,6 ± 41,86 y 26,8 ± 7,8 JI en las texturas F y FarA, respectivamente.
En adultos de broca se observó emergencia de JI por las regiones intersegmentales del insecto, iniciando con la separación de la cabeza y el tórax del insecto (Fig. 2.A.Β.) y culminando con la separación de todos los segmentos, hasta dejar totalmente desintegrado el insecto (Fig. 2.C.). De la misma forma se presentó un desarrollo completo pasando de JI, una vez llega al interior de la larva de broca, a J4, formándose finalmente el adulto de H. bacteriophora, que rompe la cutícula del insecto y emerge muchas veces con su carga de huevos. En la Figura 2 (D.E.F.) se presenta la secuencia de desarrollo encontrada en las disecciones para este evento.
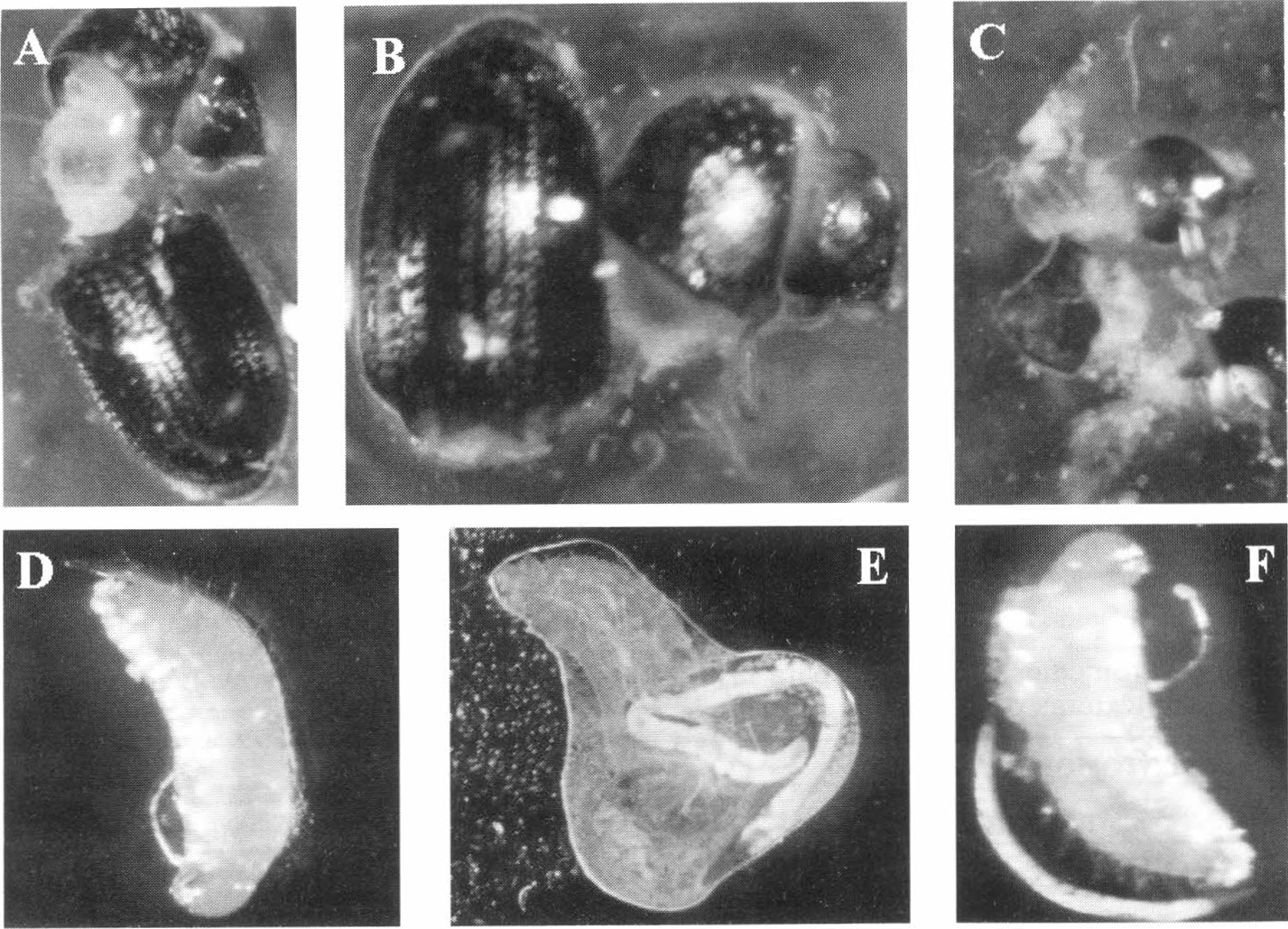
A.B.C. Secuencia del proceso de emergencia de JI de adultos de H. hampei D.E.F. Secuencia del proceso de emergencia de adultos de H. bacteriophora de larvas de H. Hampei. Fotos tomadas por Juan Carlos López N. y D, E, F tomadas por Juan Pablo Molina A. Disciplina de Entomología. Cenicafé.
Finalmente tanto en larvas como en adultos de broca dentro de fruto, se presentó un completo desarrollo de H. bacteriophora. Estudios realizados en laboratorio con estadios de la broca, también han evidenciado la reproducción de JI de Heterorhabditidos en adultos y larvas de broca (Alard y Moore 1989; Castillo 1995).
Desplazamiento de entomonemátodos hacia cerezas infestadas, colocadas sobre texturas de suelo
Se presentó igualmente para esta posición de fruto, una respuesta muy baja para las variables evaluadas, IA e IPF, con promedios de 0,83 ± 0,14% y 0,66 ± 0,03% respectivamente. Estos resultados corroboran los obtenidos en los experimentos con arena, donde el Jl no es capaz de desplazarse desde el suelo hasta subir por la superficie del fruto y penetrarlo, sino que necesariamente tiene que haber un sustrato (puente) con el orificio de perforación del fruto para que los JI accedan a éste.
Conclusiones
Ambas especies de entomonemátodos tienen alta capacidad de desplazamiento hacia frutos cubiertos por arena, a partir del punto de inoculación. Sin embargo, el Índice Estático (IE) para la especie S. feltiae fue menor que para H. bacteriophora. Lo anterior indica que S. feltiae parece tener mayor capacidad de desplazamiento en grupos de Jl que H. bacteriophora.
Con las variables IA e IPF se demuestra que los entomonemátodos que migraron del punto de inoculación se dirigieron en dirección al fruto infestado e inclusive llegaron hasta el interior del mismo.
Ambas especies de entomonemátodos tuvieron mayor preferencia y atracción hacia frutos secos. Sin embargo, el fruto maduro, a pesar de que presentó una respuesta menor, atrajo a los EN a su interior y se observó un efecto en la reducción de poblaciones de broca.
A pesar de que H. bacteriophora mostró menores índices de desplazamiento, alcance y penetración menores que S. feltiae, el PMIB fue mayor, lo que indica que algunos JI de H. bacteriophora tuvieron una mayor velocidad de desplazamiento, los cuales penetraron y mataron los estadios de broca dentro del fruto en un tiempo menor que los JI de S. feltiae.
En el experimento utilizando frutos dispuestos sobre arena, se demostró la preferencia de desplazamiento y penetración de S. feltiae por frutos secos.
Cuando el sustrato (arena o suelo) está en contacto directo con el fruto se facilita más el ingreso de los JI al orificio de perforación del fruto.
La textura del suelo no tuvo efecto sobre el desplazamiento de las especies de entomonemátodos, ya que éstos alcanzaron el fruto, lo penetraron y en su interior se presentó una importante mortalidad de individuos en diferentes estados biológicos de la plaga.
A pesar que no se presentaron diferencias estadísticas en el desplazamiento y penetración entre las dos especies de entomonemátodos en las diferentes texturas de suelo, H. bacteriophora fue la única especie capaz de infectar y reproducirse dentro de estados biológicos de la broca presentes en el interior del fruto.
S. feltiae fue la única especie que presentó una respuesta positiva a la variable IA, cuando tienen que alcanzar frutos de café que se encuentran sobre diferentes texturas de suelo.
El uso de entomonemátodos, para el control de poblaciones de broca en el suelo, se presenta como una herramienta efectiva que podría constituir un nuevo componente dentro del MIB; sin embargo, es necesario continuar con estudios referentes a la producción masiva de entomonemátodos, formulación y evaluación de las especies nativas en campo.
Recomendaciones
Se deben realizar trabajos de investigación básica para evaluar las características de desplazamiento y penetración de especies nativas de entomonemátodos aisladas de suelos de zona cafetera e investigar métodos de producción masiva in vitro de entomonemátodos con el objetivo de elaborar una formulación para la aplicación en frutos infestados con broca del café en el campo.
Footnotes
Agradecimientos
A los Drs. Bernard Briscoe y William Hominick del CABI por el suministro de las especies de EN, al personal de la Disciplina de Entomología del Centro Nacional de Investigaciones de Café "Cenicafé", en especial al Sr. Uriel Posada por su colaboración durante la realización del experimento en el laboratorio, al Dr. Fernando Cantor de la Facultad de Ciencias Básicas de la Universidad Militar Nueva Granada (Bogotá), por sus valiosos aportes en la versión final del documento.