
Abstract
The fly Ensina hyallipennis (Diptera: Tephritidae) and the moth Homoeosoma oconequensis (Lepidoptera: Pyralidae) are two phytofagous insects registered in Colombia by first time. Their larvae are seed feeders of the composite weeds "cerraja" Sonchus oleraceus and "escobilla" Senecio inaequidens. These weeds, highly aggressive in cold area in Colombia, are very difficult to eradicate by conventional ways. This research was conducted in order to evaluate the insect's potential of biocontrol through the study of some aspects of their life history, habitats, infestation levels, field damage, and natural enemies. The results show that the fly E. hyallipennis had high infestation levels and field damage of 94.2% affected plants and 72.5% affected head flowers/plant; its life cycle was completed in laboratory conditions in 20 average days, into the affected head flowers, which arrives to the ripening with few or none seed. Furthermore, natural enemies were not found under studied conditions. By the other hand, the moth H. oconequensis has an acceptable control potential on S. inaequidens because it has very low infestation levels and field damage of 9.8% affected plants and 10.6% affected head flowers; its life cycle was completed in laboratory conditions in 62 average days. The consume of the immature seeds and the prevention head flower ripening were the damage. As natural enemy, a fly (Diptera: Tachinidae) parasitoid was found, that can reduce its field control potential.
Introducción
En los ecosistemas agrícolas, las malezas han adquirido una importancia económi- ca significativa para el hombre, y son con- sideradas especies plaga que, en algunas ocasiones, generan problemas fitosani- tarios de igual o mayor magnitud que los ocasionados conjuntamente por insectos y enfermedades (Pimentel 1997). En nues- tro país, el promedio de pérdidas anuales, ocasionadas por las malezas en cultivos permanentes y transitorios, fueron para el año de 1996, de cerca de 27.000 millones de pesos, representandos entre un 18-20% de los costos totales de producción (ICA 1999).
El interés actual por la preservación del medio ambiente y la disminución en los costos de producción han orientado las investigaciones en el área agrícola, a la búsqueda y empleo de sistemas de pro- ducción enmarcados dentro del esquema del desarrollo sostenible. Específicamente para el área de malezas, la ineficiencia de las medidas de control manual y/o mecá- nico y los costos altos empleados en el desarrollo de nuevos herbicidas, han ubi- cado al control biológico como una alter- nativa autosostenible, de mínimo impacto ambiental y, a largo plazo, más eficiente (King y Melo 1994).
A raíz de la importancia adquirida por el com- ponente biológico dentro del manejo inte- grado de malezas, y debido a la escasez de proyectos en Colombia, en los cuales se haya profundizado en la generación de conoci- miento científico y desarrollo de tecnologías que permitan la implementación de progra- mas de biocontrol de tipo aumentativo, el área de Malezas del Programa Nacional MIP de Corpoica inició un proyecto de búsqueda y evaluación de agentes biológicos nativos como alternativa para el manejo de especies malezas de clima frío, con énfasis en el área de la sabana de Bogotá.
Como especies candidatas para un pro- grama de biocontrol se seleccionaron las malezas Sonchus oleraceus y Senecio inaequidens, especies de reproducción sexual y de difícil erradicación por méto- dos convencionales debido a su alta cара- cidad reproductiva y competitiva. S. oleraceus, conocida comúnmente como "cerraja", es una especie anual, de origen europeo, considerada como mediana a al- tamente nociva en clima frío (Bermudez 1997). La cerraja es una de las malezas más frecuentemente asociada con siste- mas de cultivos transitorios de hortalizas, papa, y cereales menores, encontrándose igualmente en praderas, jardines, orillas de carreteras y áreas no cultivadas (Fuen- tes y Plaza 1992).
Por su parte, S. inaequidens, conocida has- ta el momento como "escobilla", es una especie bianual, de origen sudafricano, introducida a Colombia en 1989. Se en- cuentra en la actualidad colonizando e in- vadiendo las praderas dedicadas a la pro- ducción lechera en la sabana de Bogotá, los bordes de caminos, los terrenos pastoriles abandonados o descuidados y en diferentes áreas consideradas no agrí- colas. Esta especie pertenece al primer género que señaló resistencia a las atrazinas (García-Torres y Fernández- Quintanilla 1991), lo que se ha compro- bado en el país, debido a la tolerancia alta que ha mostrado al grupo de herbicidas empleados para el control de malezas de potreros (Arrieta 1996).
Aponte et al. (1997) reconocieron como enemigo natural de la cerraja a la mosquita Ensina hyallipennis, cuyas larvas consumen las semillas en formación dentro de los bo- tones florales de esta especie, y como ene- migo natural de la escobilla, a la polilla Homoeosoma oconequensis, que en esta- do larval consume sus semillas en formación.
En vista del daño que causan estos insec- tos a las estructuras reproductivas de las malezas en mención y dada la inexistencia de estudios sobre ellos, se realizó el pre- sente trabajo, con el objetivo de generar información acerca del potencial de los insectos E. hyallipennis y H. oconequensis como agentes de biocontrol de la cerraja y la escobilla, respectivamente, para lo cual se propuso conocer algunos aspectos bá- sicos de su biología y hábitos, caracterizar el daño producido a las malezas, evaluar los niveles de infestación y daño que pre- sentan en campo y reconocer algunos de sus enemigos naturales.
Materiales y Métodos
La presente investigación se desarrolló en dos fases: la primera se llevó a cabo du- rante el primer semestre de 1998, en di- ferentes fincas de la sabana de Bogotá, con condiciones climáticas de 14 ±2° C y 82 ± 3% de humedad relativa. La segunda fase se realizó en los laboratorios y casa de mallas del C.I. Tibaitatá de Corpoica, ubicado en el municipio de Mosquera (Cun- dinamarca) a 2640 msnm y temperatura promedio de 14°C.
Niveles de infestación y daño en campo
Evaluación de la infestación y el daño por Ensina hyallipennis en cuatro fincas de la sabana de Bogotá con invasión de la cerraja Sonchus oleraceus
IC* Intervalo de confianza promedio.
PR*= Potencial Reproductivo= n° semillas viables/botón.
Evaluación de la infestación y el daño por Homoeosoma oconequensis en cuatro fincas de la sabana de Bogotá con invasión de la escobilla Senecio inaequidens
IC* Intervalo de confianza promedio.
PR* Potencial Reproductivo= n° semillas viables/botón

A estos niveles se les aplicó la prueba de normalidad de Kruskal-Wallis, se les calcu- laron sus intervalos de confianza con un nivel de significancia de 95%, y se clasifica- ron mediante la siguiente escala de infesta- ción propuesta por los autores: nivel muy bajo menor de 10%, nivel bajo de 10-30%, nivel medio de 30-70% y nivel alto mayor de 70%. De igual forma, cada botón ataca- do se disecó al estereoscopio para registrar el número de larvas presentes, con lo cual, se determinó la distribución poblacional de cada especie, mediante la aplicación del índice de Dispersión de Lloyd (L), siendo L= m/M, donde M= media poblacional, m= M+ (S2/M)-1 y S= varianza poblacional. Este índice se interpreta de la siguiente manera: L>1=Distr. agregada, L=1 Distr. aleatoria, L<1 Dist. regular.
Ciclos de vida y hábitos
Esta fase del estudio se realizó durante 1998. Las crías de los dos insectos se ini- ciaron a partir de las larvas encontradas en los muestreos de la fase anterior y se mantuvieron en el Laboratorio de Malezas del Programa MIP en el C.I. Tibaitatá de Corpoica, con temperatura de 22 ± 2 °C y 55±5% de humedad relativa. Los boto- nes atacados se confinaron por grupos para cada insecto en cámaras de cría con- formadas por frascos plásticos de 10 cm de alto x 5 cm diámetro, con papel filtro humedecido en el fondo y cubiertos con anjeo fino para permitir la aireación. Las larvas de la mosquita E. hyallipennis com- pietaron su desarrollo dentro de los boto- nes provenientes de campo. Las larvas de la polilla H. oconequensis debieron ser observadas diariamente para suministrar- les botones sanos, cuando fuera necesa- rio, hasta el final de su periodo larval. Una vez los insectos alcanzaron el estado de pupa, los restos de los botones consumi- dos se retiraron de las cámaras de cría y las pupas se dejaron allí hasta la emergencia de los adultos. Durante todo este tiempo se realizaron observaciones detalladas so- bre las cámaras de cría, con el fin de detec- tar la aparición de individuos parasitados o con síntomas de afección por agentes entomopatógenos.
Una vez emergieron los primeros adultos en laboratorio se determinó su sexo, sus características morfológicas, las dimensio- nes de machos y hembras, y se enviaron algunos ejemplares, para la determinación taxonómica al International Institute of Entomology incorporado al CABI Bioscience U.K, en donde quedaron ejemplares de re- ferencia, al igual que en la Colección Taxonómica Nacional. Luis María Murillo, del programa MIP en el C I Tibaitatá.
Los adultos sexados se ubicaron por pare- jas en cámaras de oviposición, conforma- das para las dos especies, por frascos de vidrio de 13 cm de alto y 5.5 cm de diáme- tro de boca, cubiertos con anjeo fino y en el fondo con papel filtro humedecido. Como alimento para los adultos de la mosquita E. hyallipennis se suministraron gotas de solución de miel al 10% en las paredes del frasco, y para la polilla H. oconequensis, algodón embebido en la misma solución, pendiendo de la boca del frasco. Como sustrato de oviposición se colocaron botones de la maleza respecti- va, dentro de las cámaras, adicionando agua para evitar su desecamiento. Los bo- tones se renovaron diariamente, y con la ayuda de un estereoscopio se disecaron para precisar la ubicación de las posturas dentro de éstos. Adicionalmente, en las cámaras de oviposición de la mosquita E. hyallipennis se realizaron observaciones de hábitos y duración de precópula, cópu- la, preoviposición y tiempo transcurrido entre la cópula y la oviposición. Las oviposiciones así obtenidas se incubaron en cajas de petri con un fondo de papel filtro previamente humedecido para evitar su desecación. De los huevos se determi- naron las dimensiones, la morfología y el periodo de incubación. Inmediatamente después de la eclosión, las larvas de cada insecto se colocaron individualmente con la ayuda de un pincel fino, dentro de bo- tones sanos de plantas mantenidas en la- boratorio, de la maleza respectiva. Los botones se marcaron y aislaron dentro de bolsas de polietileno transparente de cali- bre bajo, realizando un seguimiento dia- rio a los cambios externos y los síntomas de daño mostrados por éstos, sin perturbarlos hasta la salida de las larvas para pupar. Una vez obtenidas las pupas, los botones atacados se observaron al estereoscopio para realizar la descripción del daño interno, y las pupas se ubicaron en las cámaras de cría descritas para cada especie, en las cuales se determinó la mor- fología, dimensiones y duración de este estado. Con los adultos emergidos se re- novaron las crías de laboratorio, y se dio inicio a un nuevo ciclo.
El ensayo se realizó bajo condiciones de casa de mallas con temperatura promedio de 14± 2°C y humedad relativa de 58± 3%. Como unidades experimentales se emplearon jaulas de madera y anjeo de 60 cm ancho x 60 cm largo x 1,20 m alto, con tres plantas por jaula, dentro de las cuales se liberaron 5 parejas copuladas de adul- tos. Para cada insecto la unidad experi- mental contó con seis repeticiones. Cada una de las plantas empleadas en este en- sayo, tuvo en promedio 60 botones flora- les, con aproximadamente diez botones por estado, los cuales se diferenciaron por medio de cintas de color. Los adultos de E. hyallipennis permanecieron en las jaulas por siete días, mientras los de H. oconequensis por cinco, teniendo en cuen- ta la duración de cada estado floral de la escobilla y la cerraja, respectivamente. Fi- nalizado este tiempo, las plantas se lleva- ron al laboratorio, para el conteo bajo estereoscopio del número de posturas pre- sentes por estado floral; estos valores se compararon mediante una prueba de chi- cuadrado con un nivel de significancia de 95%, partiendo de la hipótesis nula según la cual, no existía preferencia por ovipositar en ningún estado de desarrollo floral.
Resultados y Discusión
Ensina hyallipennis Hennig (Diptera: Tephritidae)
El presente es el primer registro para Co- lombia de la mosquita E. hyallipennis. Esta especie fue descrita originalmente del Perú, y registrada por Prado y Nitsche (1989) atacando botones florales de la cerraja en Bolivia y Chile. Steyskal (1970) señaló que esta especie pertenece a un género cono- cido previamente como Protensina, el cual cobija a dos especies más, E. brevior (Hennig) y E. longiceps (Hendel), colecta- das originalmente en Perú.
Ciclo de vida y hábitos
El ciclo de vida de la mosquita E. hyallipennis bajo condiciones de laboratorio, desde la postura del huevo hasta la emer- gencia del adulto, se completó en un pro- medio de 44 días. Los adultos machos vi- vieron 19.5 días promedio y las hembras 24 días. Los datos de la duración del ciclo y las dimensiones de cada estado de desa- rrollo se presentan en la Tabla 3.
Tamaño y duración de cada uno de los estados de desarrollo de Ensina hyallipennis en condiciones de laboratorio con la duración promedio y rangos de cada estado
Dimensiones correspondientes al último ínstar.
Adulto
El adulto es una mosquita de alas anchas, hialinas y con una zona más oscu- ra en la célula subcostal (vértice de unión de las venas R1 y costal), siendo el macho más pequeño que la hembra (Tabla 3). El tórax es de color negro, presentando en los flancos áreas irregulares de color ama- rillo. El abdomen es de forma oval, más largo y abultado que el tórax y cubierto por setas negras muy finas; ventralmente es de color amarillo, mientras que su parte dorsal presenta bandas transversales de color negro. La hembra se diferencia del macho por su ovipositor largo, notorio a simple vista (Fig. 1a).
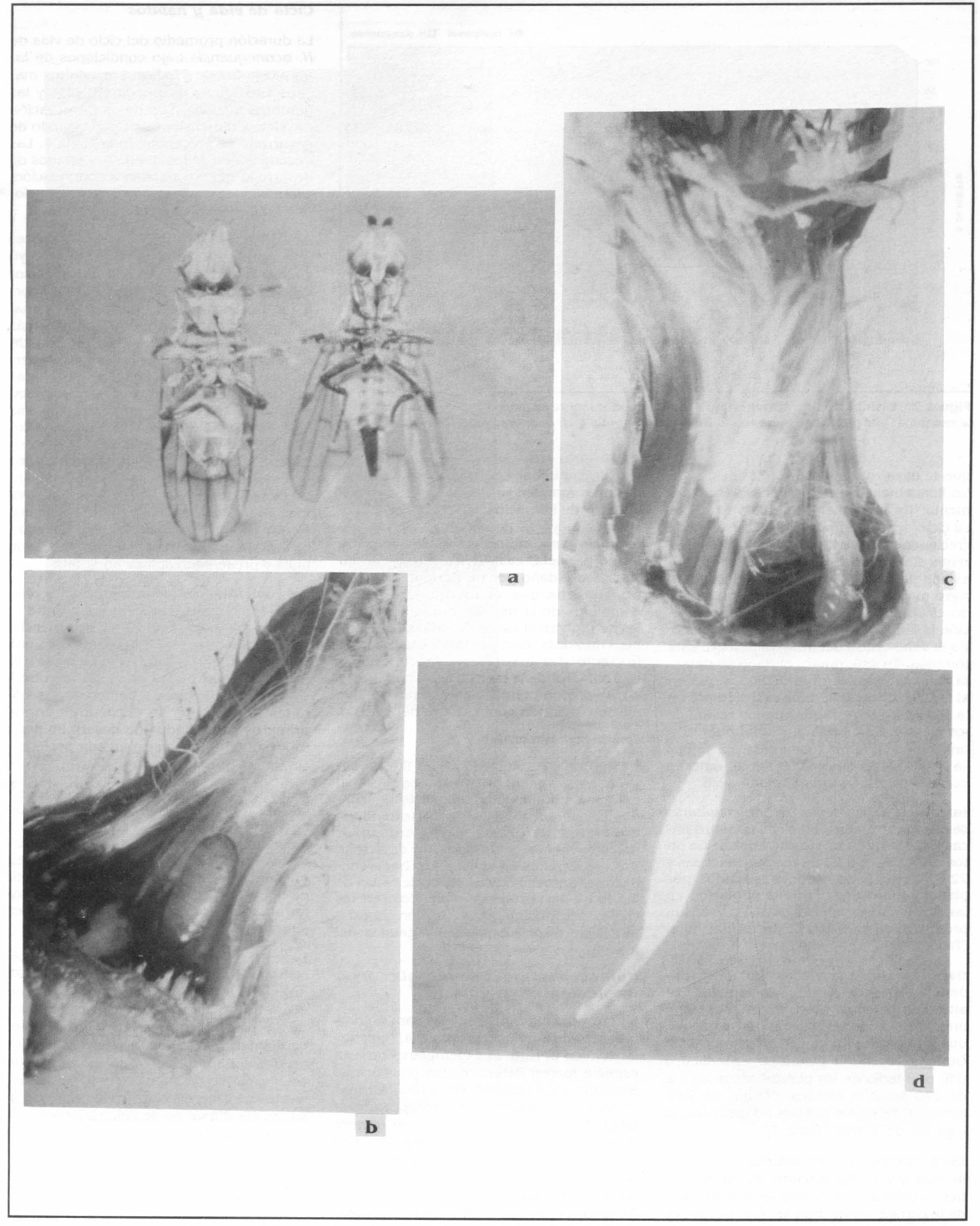
Aspectos de E. hyallipennis en la cerraja: a. Adulto macho y hembra (20x). b. Pupa y restos del botón atacado (20x). c. Larva consumiendo semillas en formación (20x). d. Huevo recién puesto (100x). Fotos: A. Nájar.
Precópula-cópula
El período de pre-có-pula, en E. hyallipennis, se inicia con un acercamiento del macho a la hembra por medio de vuelos cortos realizados a su al- rededor. Al cabo de 1 a 2 minutos, el ma- cho se posa sobre el dorso abdominal de la hembra, tratando de inmovilizarla con sus patas posteriores y ayudado de su aparato bucal con el cual la estimula por unos se- gundos, hasta que ésta se queda inmóvil. La cópula se inicia cuando la hembra incli- na su ovipositor formando un arco hacia el abdomen del macho, y se desplaza horizontalmente para elegir el sitio de per- manencia durante este período. Ocasional- mente, el macho se mueve lateralmente, quizá con el objeto de desalojar su conte- nido seminal en el oviducto de la hembra. La cópula se presenta entre el primero y el segundo día de emergencia de los adultos, tiene una duración de 30 minutos a dos horas, observándose en una pareja hasta tres cópulas por día. Al finalizar la cópula, la hembra se separa del macho y permane- ce inmóvil por unos minutos mientras que éste vuela en busca de alimento.
Oviposición
El inicio de la oviposición sucede el mismo día de la cópula o hasta el tercer día. La hembra, coloca entre 2-4 huevos por día, de 1-3 huevos por botón y de 8-13 huevos a lo largo del período de oviposición. Esta se realizó únicamente dentro de los botones florales que se en- contraban en los primeros dos estados de desarrollo (p= 0.0005), con una preferen- cia marcada por ovipositar en los botones en estado uno (Fig. 2).
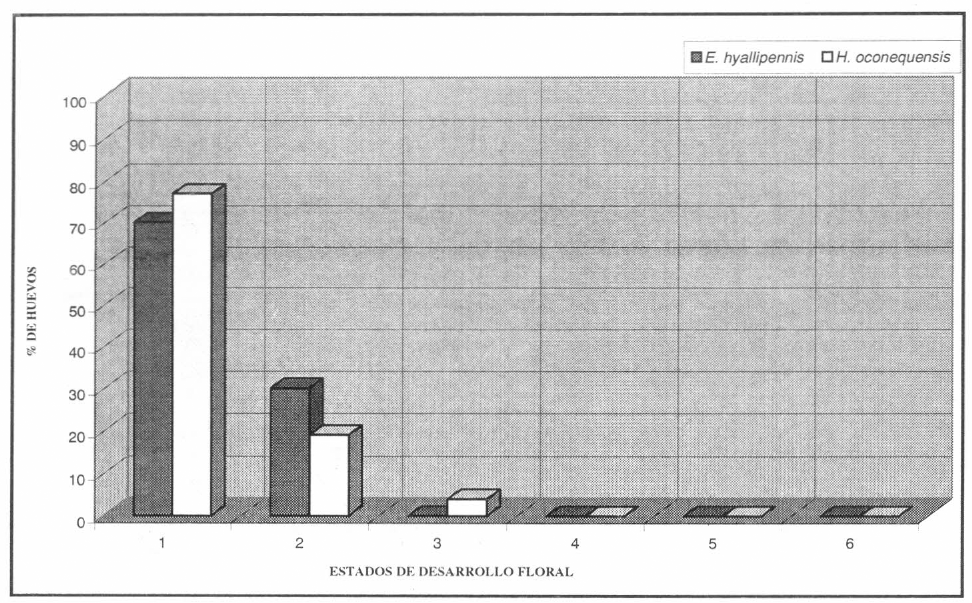
Distribución de la preferencia de oviposición sobre estados florales de la cerraja y la escobilla, por parte de las hembras de E. hyallipennis y H. oconequensis, respectivamente.
Huevo
Los huevos de este insecto poseen el corion liso y delgado, son alargados, curvos, con un extremo agudo y el otro con un pedicelo largo (Fig. 1d). Son colo- cados individualmente entre los sépalos del involucro floral o cerca a los tubos de las corolas de las flores individuales. Re- cién puestos son transparentes, pero a medida que se acerca la eclosión toman una coloración blanco cremosa.
Larva
La larva es de tipo vermiforme. Los ganchos bucales son visibles durante toda la fase larval. Presenta 11 segmentos. Re- cién nacida es de color blanco cremoso y mide, en promedio, 0.5 mm de longitud y 0.4 mm de diámetro, mientras que la larva madura presenta una coloración amarilla que se intensifica gradualmente del extre- mo posterior al anterior, y mide en prome- dio 5.2 mm de longitud y 1 mm de diámetro. No se estableció el número de instares larvales dada la imposibilidad para la recuperación de las exuvias dejadas den- tro de los botones florales.
Pupa
El puparium es de forma cilíndrica y de tipo coartata. Inicialmente es de color grisáceo claro pero se va oscureciendo a medida que se acerca la emergencia del adulto.
Infestación y daño en campo
Estos niveles altos de infestación y daño indican que E hyallipennis afecta de ma- nera significativa el potencial reproductivo de la cerraja y muestran las perspectivas y potencialidades de su utilización como agente de control de esta maleza. Sin em- bargo estos resultados plantean la necesi- dad de continuar con una segunda etapa de investigación, en la cual se evalúe el impacto de liberaciones del insecto a den- sidades mayores de las que se encontra- ron bajo las condiciones del estudio. De forma paralela, evaluar los niveles de in- festación y daño por un periodo de tiem- po que abarque al menos dos ciclos completos de la maleza, durante los cua- les se registre la variación de las condicio- nes climáticas. Esto permitirá establecer el impacto del insecto en la capacidad reproductiva de la maleza, y a su vez, defi- nir si existen condiciones climáticas que afecten la acción controladora del insecto.
Enemigos naturales
Del material recolectado en las fincas visi- tadas, no se encontraron insectos ni patógenos que afecten al insecto en esta- do larval, por lo cual la búsqueda de enemigos naturales debe enfocarse posteriormente, a las fases de desarrollo no consideradas en el estudio y hacia con- diciones agroecológicas diferentes a las de los lotes de hortalizas evaluados, en las cuales adicionalmente, se explore la exis- tencia de otros huéspedes vegetales del insecto.
Homoeosoma oconequensis Dyar (Lepidoptera: Pyralidae)
Se presenta el primer registro para Colombia de la polilla H. oconequensis, registra- da sólo en el Perú. Los ejemplares de esta especie fueron determinados por Michael Shaffer, especialista del International Institute of Entomology incorporado al CABI Bioscience U.K. Dentro del género Homoeosoma existen otras especies con hábitos similares de alimentación, como las polillas H. deltaparenensis y H. ammonastes registradas por Pastrana (1991) en Argentina y México, consumiendo en esta- do larval, las semillas de las malezas Senecio bonariensis y Cirsium repandum, respectivamente.
Ciclo de vida y hábitos
La duración promedio del ciclo de vida de H. oconequensis bajo condiciones de la- boratorio fue de 73 días. Los adultos ma- chos vivieron en promedio 10 días y las hembras 12 días. Los datos de duración del ciclo y dimensiones de cada estado de desarrollo se presentan en la Tabla 4. Las descripciones de los diferentes estados de desarrollo que se señalan a continuación, están basadas únicamente en observacio- nes realizadas en el laboratorio.
Tamaño y duración de cada uno de los estados de desarrollo de Homoeosoma oconequensis en condiciones de laboratorio con la duración promedio y rangos de cada estado
Dimensiones correspondientes al último ínstar.
Adulto
El adulto de H. oconequensis es una polilla de hábitos nocturnos, cuyo cuerpo está cubierto por escamas de color marrón grisáceo claro (Fig. 3a). El dimor- fismo sexual está relacionado con el tama- ño de los adultos, siendo el macho más pequeño que la hembra (Tabla 4). La ca- beza es pequeña, del mismo color del cuer- po, se encuentra cubierta por escamas grandes y largas, y presenta palpos labia- les prolongados hacia adelante. Las alas anteriores son elongadas y del mismo co- lor del cuerpo, mientras que las posterio- res son más anchas, casi triangulares, y con la región medial-cubital-anal de color blan- co perla, casi traslúcido. El abdomen es más largo que el tórax y de forma ahusada, siendo en la hembra mucho más robusto y ovalado que en el macho. Las patas son finas y presentan espinas en la tibia.
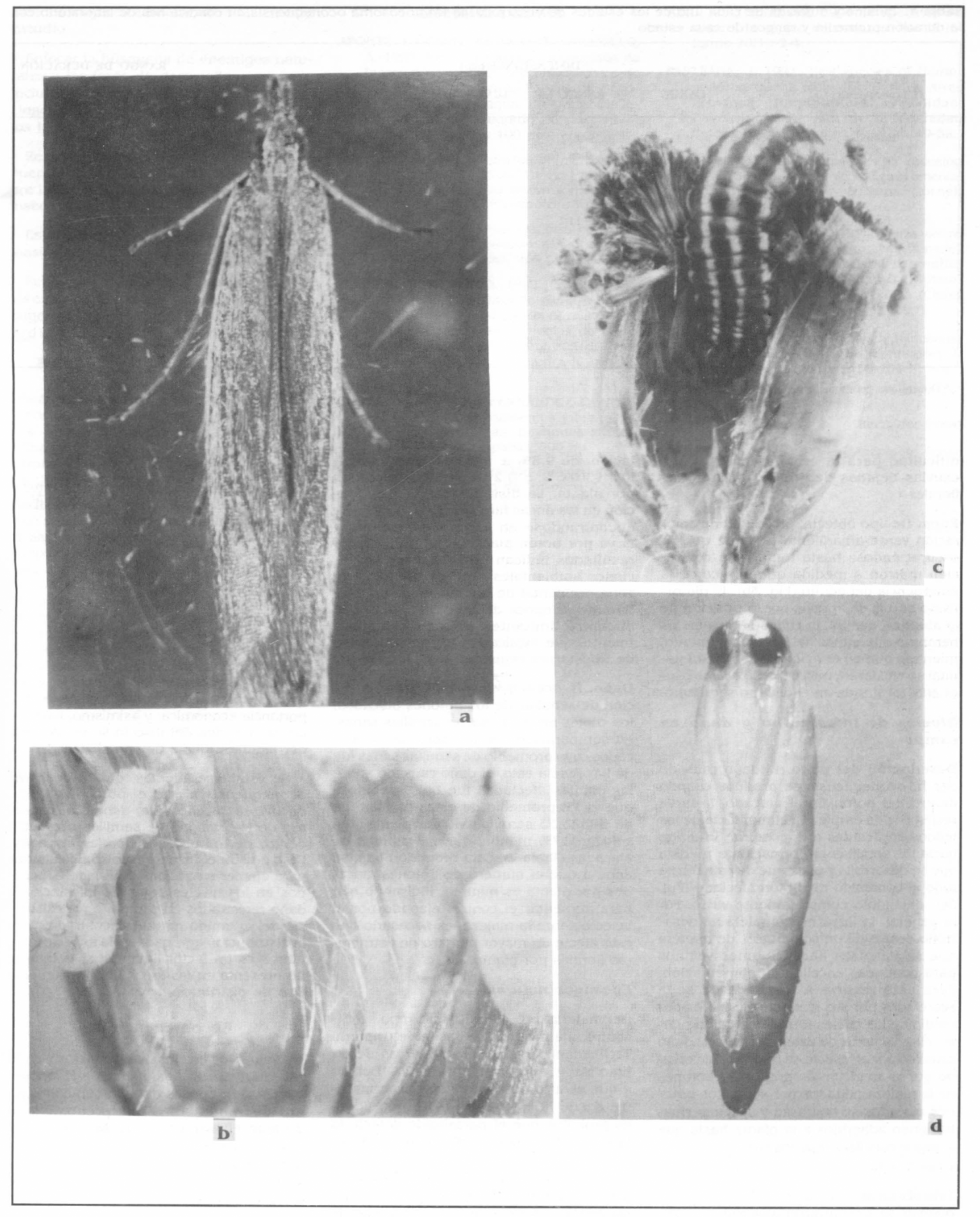
Aspectos de H. oconequensis en la escobilla: a. Adulto (10x). b. Pupa (10x). c. Larva consumiendo semillas en formación (10x). d. Huevo dentro de botón floral (100x). Fotos: A. Nájar.
Oviposición
Las hembras de H. oconequensis comienzan a ovipositar entre el tercer y el cuarto día de su emergencia, colocando entre 2 a 7 huevos por día, uno por botón floral, y entre 11-39 huevos a lo largo de todo el periodo. Las hembras ovipositan únicamente dentro de los bo- tones florales que se encuentran en los primeros tres estados de desarrollo floral (p=0.0005), con una marcada preferen- cia por el primer estado (Fig.2).
Huevo
Los huevos de H. oconequensis son de forma ovalada y poseen el corion ligeramente estriado. Se colocan en forma individual entre los tubos de las corolas de las flores individuales (Fig. 3d). Recién puestos son de color blanco, pero a medi- da que se acerca la eclosión toman un co- lor amarillo oscuro, haciéndose visible en su interior la cabeza de la futura larva.
Larva
La larva es de tipo eruciforme. Re- cién nacida es de color blanco y mide en promedio 0.7 mm de largo por 0.2 mm de diámetro. La larva madura alcanza un pro- medio de 14.5 mm de largo por 2.4 mm de diámetro, presenta la cabeza y la placa protoráxica fuertemente esclerotizada, de color amarillo naranja; la superficie ventral es de color verde claro, mientras que la superficie dorsal es de color crema y pre- senta 3 bandas longitudinales de color carmín. Al finalizar su desarrollo la larva abandona el botón floral y cae al suelo donde construye un capullo de seda de color blanco verdoso dentro del cual per- manece como prepupa por 2-4 días. En este trabajo no fue posible precisar el nú- mero de ínstares larvales dada la extrema dificultad para la recuperación de las exuvias dejadas dentro de los botones florales.
Pupa
De tipo obtecta, presenta una colo- ración verde amarillento brillante que va oscureciéndose hasta tomar una colora- ción marrón a medida que se acerca la emergencia del adulto (Fig. 3b). El dimor- fismo sexual se aprecia por la posición de la abertura genital, la cual se ubica en las hembras a la altura del octavo segmento, mientras que en el macho, la abertura ge- nital se encuentra más cercana a la abertu- ra anal localizada en el décimo segmento.
Niveles de infestación y daño en campo
Enemigos naturales
Del material proveniente de campo se en- contró una mosquita parasitoide (Diptera: Tachinidae), emergiendo de larvas de úl- timo ínstar, en porcentajes de 2%. Debido a que el ciclo de vida del insecto transcu- rre desde huevo dentro del botón floral, es probable que el parasitoide detecte la larva de H. oconequensis, cuando ésta se desplaza hacia otros botones para conti- nuar en ellos su alimentación. Este ene- migo natural encontrado, reduce en campo las poblaciones del insecto, influyendo de esta forma en los niveles bajos de infesta- ción registrados.
Conclusiones
Los resultados obtenidos en este estudio muestran las perspectivas y potencialida- des de E. hyallipennis como agente de con- trol de la cerraja, por tratarse de un insecto adaptado a las condiciones agroecológicas de la sabana de Bogotá, por el daño pro- ducido sobre el potencial reproductivo de las plantas afectadas, y por presentar un ciclo de vida sincronizado con el desarrollo floral de su huésped. Sin embargo, es ne- cesario aclarar en estudios posteriores si el daño producido por el insecto afecta la ca- pacidad reproductiva de la maleza, y por tanto su prevalencia como especie de im- portancia económica; y asimismo, estable- cer si la acción del insecto se ve afectada por alguna variable de tipo climático.
Se establece además, que la polilla H. oconequensis afecta el potencial repro- ductivo de la escobilla, debido al consu- mo de la totalidad de semillas presentes dentro de los botones florales atacados. Pese a esto, el insecto mostró dificultades para ejercer su acción controladora, basa- dos en los niveles bajos de infestación y daño registrados en campo, en la existen- cia del enemigo natural encontrado, y en la distribución agregada de la especie; esta última es un factor limitante si la maleza no presenta en las áreas de invasión este tipo de distribución.
Recomendaciones
Evaluar los niveles de infestación y daño causados por E. hyallipennis y H. oconequensis, en épocas del año con condicio- nes diferentes a las que predominaron durante el estudio, e igualmente exten- der los muestreos a otras regiones agroecológicas en las cuales las malezas se encuentren presentes en niveles de daño económico.
Evaluar el impacto sobre las malezas de liberar las dos especies de insectos a den- sidades mayores a las encontradas en este estudio.
Enfocar la búsqueda de enemigos natu- rales a las fases de desarrollo que no se incluyeron en este estudio, y hacia condi- ciones agroecológicas diferentes a las de los lotes de hortalizas estudiados.
Realizar estudios para determinar la influencia de las condiciones climáticas so- bre la acción controladora ejercida por los insectos.
Estandarizar metodologías para la cría masiva de los insectos en laboratorio.
Revisar la identificación taxonómica de las especies malezas y establecer el sitio de origen de las dos especies de insectos es- tudiados.
Footnotes
Agradecimientos
Los autores expresan su agradecimiento a Omar Bahamón y Esther Delgado, auxilia- res técnicos de Corpoica en el C.I. "Tibaitatá" por su valiosa colaboración durante el desarrollo de esta investigación, y a Corpoica y Pronatta por la financiación de la misma.