
Abstract
Coleoptera fauna in a colombian rain forest was compared in gaps and undisturbed forest. Five replicas in each site were studied through pitfall tramps to catch its insects. Samples were determined until family level, and Margaleff index was used to determinate family riches. Homogeneity between samples were compared and between sites through statistical tests in its rich indexes. Statistic significant differences between samples and between sites weren't found, but differences in Coleoptera fauna composition in sites were registered.
Introducción
Cerca de la tercera parte de los insectos del mundo se encuentra en el neotrópico, especialmente en Perú, Colombia y Brasil, en donde el conocimiento de la megadiversidad de insectos sólo se remite a algunos grupos, pero una gran cantidad de especies endémicas está por descubrir (Brown 1991).
El estudio de la fauna de insectos proporciona información sobre el estado de los ecosistemas, su productividad y los niveles de contaminación acuática y atmosférica, puesto que interacciones como intercambios genéticos, biomasa y energía transferidas en los ecosistemas se encuentran directamente relacionadas con las poblaciones de insectos (Brown 1991); así mismo, es importante la identificación de especies de insectos indicadoras de los diferentes tipos bosques que, mediante monitoreos periódicos, registren su continuidad en el tiempo (Nilsson et al. 1994), debido, al papel preponderante que estos organismos desempeñan como factores formadores y reguladores de los ecosistemas (Camero y Chamorro 1995).
El estudio de los insectos dentro del ámbito de la indicación del estado ecosistémico elimina la utilización de alternativas dispendiosas y costosas; permite extrapolar datos cuantitativos para otros ecosistemas similares; proporciona información acerca de las dinámicas poblacionales como competición, herbivoría y predación en insectos, y determina el estado crítico de los ecosistemas en amenaza por la acción antrópica (Brown 1991). En los ecosistemas tropicales, el estudio de la fauna de insectos es primordial en virtud al alto grado de deterioro que presentan los ecosistemas naturales, lo cual genera la extinción de gran cantidad de recursos naturales regionales, incluyendo especies aún no descritas y, posiblemente, de gran importancia económica (Gordon 1985).
Uno de los campos de más reciente investigación se relaciona con el estudio y análisis de los componentes que, de acuerdo con Wallwork (1970), constituyen la macro y mesofauna del suelo: estos organismos en unión con la microbiota, permiten distinción entre un suelo maduro y el material parental del que se ha derivado (Wood 1988), además ayudan a conservar, por lo menos en niveles estables, la fertilidad natural del medio edáfico (Cortés 1990).
Los coleópteros representan alrededor del 10% de la riqueza de la entomofauna estudiada en los suelos colombianos, que corresponden al 45% del total de familias del mundo. Cerca de 600 mil especies de coleópteros han sido descritas hasta el momento, siendo el mayor número de organismos dentro de la entomofauna mundial.
La coleopterofauna del ecosistema edáfico cumple funciones de vital importancia debido a la diversidad de nichos que éstos organismos ocupan, y a la distribución de sus poblaciones a lo largo y ancho del perfil del suelo, donde desempeñan funciones diversas como la regulación de poblaciones, contribución al flujo energético a través de las cadenas tróficas y participación en la constitución física y química de los suelos por la transformación y transporte de las unidades estructurales del material edáfico.
En Colombia se carece de información ecológica pertinente a las especies y poblaciones de coleópteros, y de colecciones de referencia lo suficientemente completas, a pesar de tener el orden Coleoptera, gran capacidad para indicar cambios en el suelo, debido a factores climáticos y/o antrópicos.
Según Campbell (1987), sería posible obtener información detallada a partir de resultados de caracterizaciones ecológicas periódicas espacio-temporales para la fauna de coleópteros en ecosistemas naturales, lo cual facilitaría el estudio del comportamiento de las comunidades naturales con miras a establecer posteriores comparaciones en ecosistemas con distinto grado de alteración.
El conocimiento de los coleópteros en Colombia apenas se inicia. Su clasificación taxonómica, su ubicación sistemática y su comportamiento poblacional en zonas de baja intervención, constituyen las herramientas básicas para la determinación de indicadores que definan la productividad y susceptibilidad de los suelos colombianos de manera rápida y consistente (Camero y Chamorro 1996).
Tanto la composición como la distribución de las especies de las familias de Coleoptera debe variar en distintos ecosistemas; las especies encontradas en transectos geográficos caracterizan un ecosistema natural en particular, debido al alto grado de especiación alopátrica, y de endemismos del neotrópico.
En el presente estudio se busca establecer las diferencias de abundancia a nivel de familia de coleópteros de dos ambientes de bosque con distinto grado de alteración.
Materiales y Métodos
Area de estudio
El presente trabajo se llevó a cabo en el bosque municipal de Mariquita, ubicado en el departamento del Tolima, a 5°12 N y 74°55 W y a 590 m de altitud (Fig. 1), y que hace parte del 34.9% del Bosque Húmedo Tropical colombiano (IGAC 1995). Tiene un área de 123 ha y presenta una temperatura media anual de 27°C. El relieve es de ondulado a quebrado, con pendientes que oscilan entre 30 y 80%. El bosque es atravesado por la quebrada "El Peñón", la cual moldea el terreno. Los suelos del bosque están constituidos, en general, por arcillas rojas, abigarradas, con altos componentes silíceos de aspecto conglomerado (Pachón y Bohórquez 1991).
En su vegetación se distinguen tres estratos verticales bien diferenciados: el estrato superior, dominado por individuos de Cassia grandis, Pera sp., Jacaranda sp. y Vochisia ferruginea; el estrato medio por Rheedia madroño, Callophylum sp., Virola sp., Oenocarpus mapora, Bactris pilosa y Myrcia sp. y el sotobosque por Astrocarium malybo, Bactris pilosa, Anthurium sp., Dieffenbachia cf. Orstedii, Inga sp., Schizaea sp., Ocotea sp., Myrcia sp., Serjania sp. y Virola sp.
Localización del área de estudio. Escala 1:1′000.000
Métodos
Se escogieron cinco sitios de muestreo para cada uno de los ambientes (claros y bosques climácicos); en cada tipo de ambiente se instalaron doce trampas pitfall para captura de insectos. La identificación taxonómica de los coleópteros se realizó hasta el nivel de familia según las claves sistemáticas de Borror et al. (1995) y Dillon y Dillon (1972). Se calcularon índices de Riqueza de Margaleff para cada ambiente y se caracterizó cada ambiente en sus grupos de coleópteros.
Para determinar la homogeneidad de las muestras en cada ambiente se utilizó la Q de Cochran y para determinar la similaridad entre los dos ambientes estudiados se utilizó la prueba de Mann-Whitney con base en el registro de las familias de coleópteros encontradas en cada ambiente.
Resultados y Discusión
Para los dos ambientes de muestreo en los diez sitios de colección, se determinaron nueve familias del orden Coleoptera: Phalacridae, Staphylinidae, Scolytidae, Nitidulidae, Scydmaenidae, Ptiliidae, Pselaphidae, Carabidae y Curculionidae. Seis de éstas fueron registradas en los dos ambientes; las familias Carabidae y Curculionidae no se registraron en ambientes de bosque climácico (Fig. 2), y la familia Scydmaenidae no se encontró en los claros (Fig. 3).
La composición de la fauna de coleópteros en ambientes de bosque climácicos (Fig. 2) muestra la presencia de siete familias: Phalacridae, Staphylinidae, Scolytidae, Nitidulidae, Scydmaenidae, Ptiliidae y Pselaphidae, entre las cuales, Staphylinidae, seguida de Ptiliidae y Phalacridae, son las que presentan el mayor número de especímenes colectados. Estos resultados concuerdan con los obtenidos por Castillo y Amat (1986), Medina (1990), Sánchez et al. (1992) y Naranjo y Peñaranda (1995), en los cuales se encontró mayor riqueza de estas familias en bosques climácicos, en comparación con la fauna colectada en ecosistemas con distinto grado de alteración.
Para ambientes de claros (Fig. 3) se estableció la presencia de ocho familias de coleópteros: Phalacridae, Staphylinidae, Scolytidae, Nitidulidae, Ptiliidae, Pselaphidae, Carabidae y Curculionidae, dentro de las cuales, las familias Staphylinidae, Scolytidae, Nitidulidae y Ptiliidae, son las que presentan mayor cantidad de especímenes colectados. Para el caso de ecosistemas intervenidos, estos resultados son similares a los obtenidos por Sánchez et al. (1992) en cuanto a la riqueza de familias como Phalacridae y Staphylinidae, y a los resultados obtenidos por Naranjo y Peñaranda (1995) para el caso de la familia Scolytidae.
En general, los resultados muestran tanto diferenciación numérica como de riqueza de las familias colectadas; lo cual indica que, de la misma forma que existen etapas sucesionales de ecosistemas en cuanto a composición florística, puede existir también una sucesión faunística en ecosistemas en recuperación, la cual es notablemente diferente con respecto a ecosistemas con un grado máximo de madurez.
De esta forma, podría pensarse que los componentes entomofaunísticos tienen variación estacional en cuanto a composición y población dependiendo de la etapa seral de desarrollo y madurez que tenga un ecosistema, y que el comportamiento de la fauna en una etapa seral dada, puede asociarse a estrategias poblacionales r y k como ocurre en procesos de composición florística.
El índice de Riqueza de Margaleff para los coleópteros colectados en cada ambiente mostró como resultado para los bosques climácicos valores entre 0 y 1.550 y para los claros valores entre 1.443 y 2.038. Sin embargo, la comparación entre las medianas de estos índices para cada ambiente no muestra diferencias significativas según la prueba de MannWhitney a un nivel de significancia de 0.05 (Tabla 1).
Indice de Riqueza para los coleópteros colectados en cada sitio de muestreo del Bosque Municipal de Mariquita.
Z=1.592 < P0.05 = 1.96
Es posible que aunque estadísticamente no se encuentren diferencias en la composición faunística entre los dos ambientes a nivel de familias de coleópteros, ésta si se pueda llegar a detectar al determinar los especímenes colectados hasta nivel de especie. Los valores de Indice de Riqueza más altos encontrados para cada ambiente; esto es, 1.550 para bosques climácicos y 2.038 para claros, sugieren que existe diferencia a nivel de composición faunística entre los dos y que no obstante la similaridad de algunos factores abióticos entre los ambientes, deben existir factores microclimáticos que inciden en la composición y la riqueza faunística.
La ausencia de familias como Carabidae y Curculionidae en ambientes de bosques climácicos, y de Scydmaenidae en ambientes de claros, según los métodos utilizados, y la variación en cuanto a número de individuos para cada familia de un ambiente a otro, son la base para relacionar la composición faunística con la madurez ecosistémica.
La comparación de homogeneidad de las muestras en los dos ambientes (Tablas 2 y 3) mediante la prueba Q de Cochran indica que no hay diferencia entre ellas; tanto las muestras colectadas en ambientes de claros como aquellas de bosques climácicos son estadísticamente similares, y la composición faunística en cuanto a familias de coleópteros entre las muestras colectadas en cada ambiente, son también, cualitativamente similares, dando como resultado familias propias para cada ambiente (Figs. 2 y 3).
Similitud entre muestras colectadas en bosques climácicos del Bosque Municipal de Mariquita (Colombia) según la Q de Cochran
Q=10.714 > P0.05 = 7.815
Similitud entre muestras colectadas en ambientes de claros del Bosque Municipal de Mariquita (Colombia) según la Q de Cochran
Q=10.667 > P0.05 = 5.991
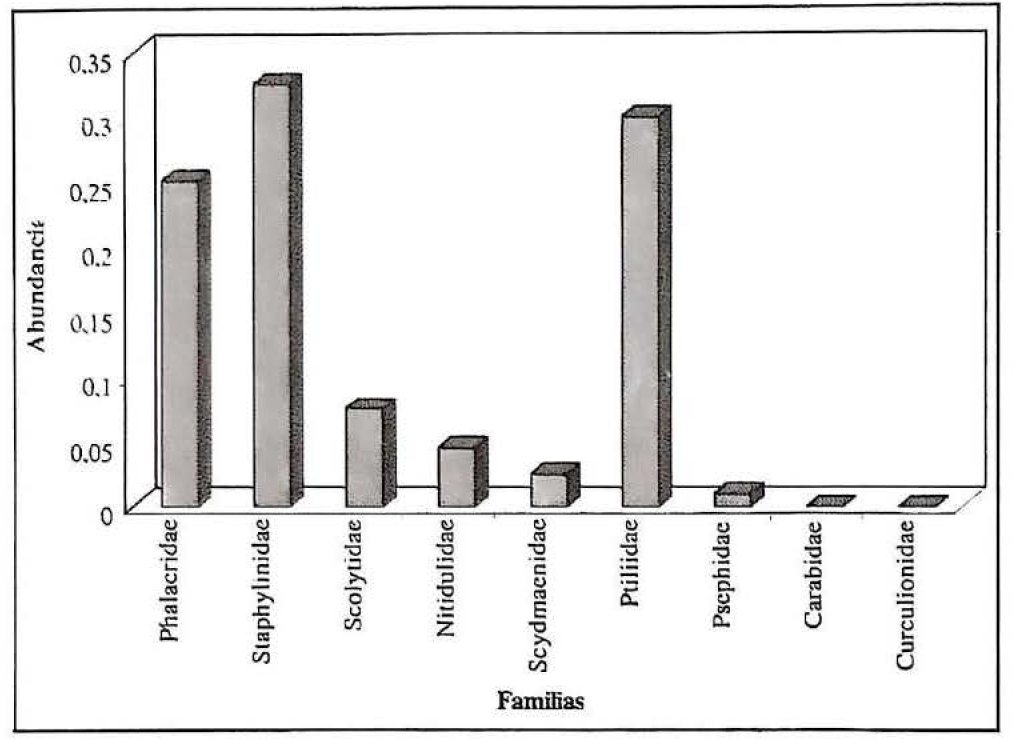
Familias de coleópteros encontradas en ambientes de bosque climácico del Bosque Municipal de Mariquita (Colombia)
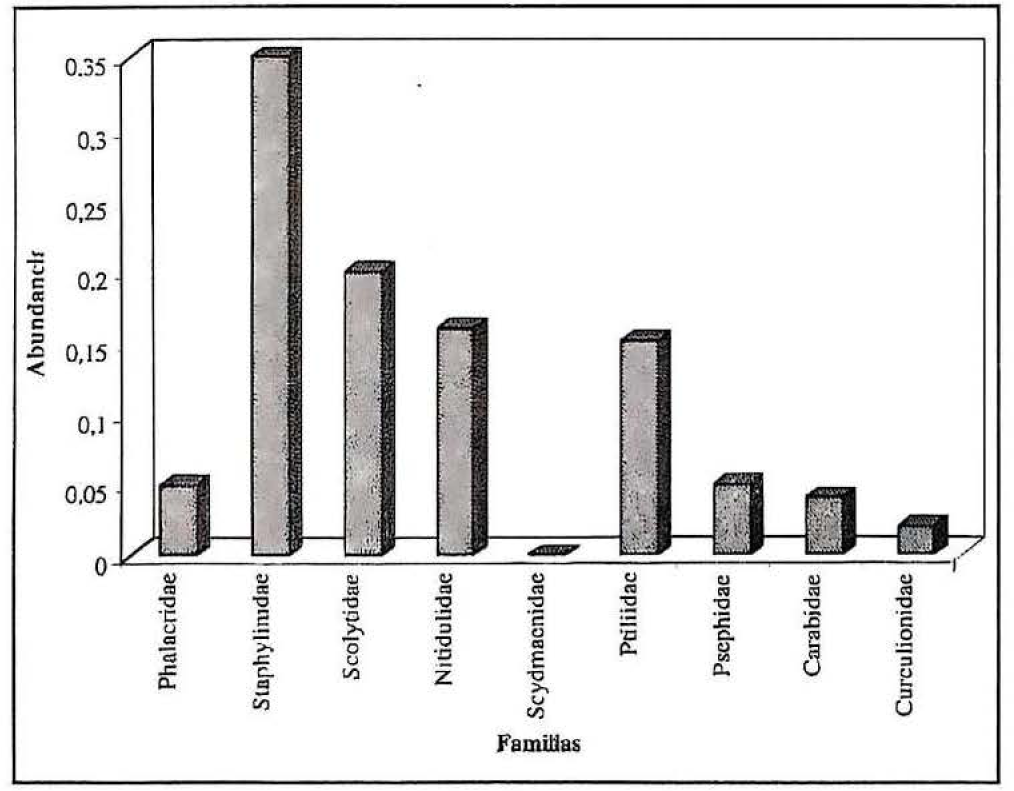
Familias de coleópteros encontradas en ambientes de claros del Bosque Municipal de Mariquita (Colombia)
Conclusiones
La composición en cuanto a presencia o ausencia de las familias de coleópteros y el número de individuos colectados por familia varía de un ambiente a otro, lo cual induce a pensar en un paralelo como ocurre en la composición florística de los ecosistemas en sus etapas serales.
Existe homogeneidad entre las muestras de cada ambiente. Las muestras procedentes de ambientes de claros, al igual que las muestras procedentes de bosque climácicos, no presentan grandes variaciones en cuanto al tipo de fauna colectada.
Los ambientes de claros y de bosques climácicos no presentan diferencias estadísticamente significativas en cuanto a similaridad de fauna de coleópteros medida en índices de riqueza, pero si presentan diferencias en cuanto a la presencia o ausencia de familias colectadas.